
Abstract
Background:
We previously identified two highly discriminating and predictive radiation-induced transcriptomic signatures by comparing series of sporadic and postradiotherapy thyroid tumors (322-gene signature), and by reanalyzing a previously published data set of sporadic and post-Chernobyl thyroid tumors (106-gene signature). The aim of the present work was (i) to compare the two signatures in terms of gene expression deregulations and molecular features/pathways, and (ii) to test the capacity of the postradiotherapy signature in classifying the post-Chernobyl series of tumors and reciprocally of the post-Chernobyl signature in classifying the postradiotherapy-induced tumors.
Methods:
We now explored if postradiotherapy and post-Chernobyl papillary thyroid carcinomas (PTC) display common molecular features by comparing molecular pathways deregulated in the two tumor series, and tested the potential of gene subsets of the postradiotherapy signature to classify the post-Chernobyl series (14 sporadic and 12 post-Chernobyl PTC), and reciprocally of gene subsets of the post-Chernobyl signature to classify the postradiotherapy series (15 sporadic and 12 postradiotherapy PTC), by using conventional principal component analysis.
Results:
We found that the five genes common to the two signatures classified the learning/training tumors (used to search these signatures) of both the postradiotherapy (seven PTC) and the post-Chernobyl (six PTC) thyroid tumor series as compared with the sporadic tumors (seven sporadic PTC in each series). Importantly, these five genes were also effective for classifying independent series of postradiotherapy (five PTC) and post-Chernobyl (six PTC) tumors compared to independent series of sporadic tumors (eight PTC and six PTC respectively; testing tumors). Moreover, part of each postradiotherapy (32 genes) and post-Chernobyl signature (16 genes) cross-classified the respective series of thyroid tumors. Finally, several molecular pathways deregulated in post-Chernobyl tumors matched those found to be deregulated in postradiotherapy tumors.
Conclusions:
Overall, our data suggest that thyroid tumors that developed following either external exposure or internal 131I contamination shared common molecular features, related to DNA repair, oxidative and endoplasmic reticulum stresses, allowing their classification as radiation-induced tumors in comparison with sporadic counterparts, independently of doses and dose rates, which suggests there may be a “general” radiation-induced signature of thyroid tumors.
Introduction
Sporadic and radiation-induced thyroid tumors have similar pathological features (2) and aggressiveness (3). Higher aggressiveness reported in short-latency solid-variants post-Chernobyl PTC is not related to radiation exposure but rather to young age at diagnosis (4). The higher frequency of RET/PTC rearrangements observed in post-Chernobyl PTC compared with sporadic PTC is not specific to radiation but rather is associated with young age at radiation exposure, with the RET/PTC3 rearrangement being more frequently observed in aggressive tumor variants that occurred with a short latency in younger children, and the RET/PTC1 rearrangement in less aggressive variants occurring with a longer latency in older subjects (5,6).
Yet radiation-induced thyroid tumors display molecular specificities, as several studies have reported sets of genes or protein markers that appeared to be deregulated in these tumors, compared with sporadic tumors (7 –12). All studies focused on post-Chernobyl tumor analysis, and only one study reported the transcriptomic signature of postradiotherapy thyroid tumors (10). However, most studies were not designed to identify, or failed to identify, sets of markers that could be suitable for an accurate case-by-case diagnosis of radiation-induced thyroid tumors (7 –9,12 –14). Some authors used post-Chernobyl PTC of short latency and sporadic PTC diagnosed later in life (7,8), and so the identified markers could also be related to the higher aggressiveness of the Chernobyl tumors. Moreover, the classification methods most commonly used for microarray analysis appeared to be unsuited to the identification of a highly discriminating transcriptomic signature of etiology when applied to rather small series of sporadic and post-Chernobyl PTC (9,11,13,14). To identify such a transcriptomic signature, our laboratory has developed a specific microarray analysis strategy (EMts_2PCA method), which is focused on the selection, from small series of samples, of highly probable candidate genes whose expression potentially distinguishes between the two groups of samples (e.g., sporadic from radiation-induced tumors), followed by the validation of these markers by the blind prediction of the etiology in an independent series of thyroid tumors (11). This strategy was successfully applied to finding transcriptomic signatures of radiation-induced sarcomas, thyroid and breast cancers (10,11,15). Regarding thyroid tumors, a 322-gene discriminating transcriptomic signature was identified from a series of postradiotherapy and sporadic follicular thyroid adenomas and PTC, and was validated by the accurate etiology classification of an independent series of 29 thyroid tumors (10). Moreover, a 106-gene signature was identified in a series of post-Chernobyl PTC previously analyzed by Detours et al. (9), which also displayed a high discriminating potential for etiologic classification of post-Chernobyl tumors (11).
The relevance of extrapolation from data on tumors occurring after exposure to external radiation therapy to post-Chernobyl tumors that occurred after internal 131I contamination is not known. To highlight the similarities/differences between the two transcriptomic signatures identified after either external exposure (10) or 131I contamination (11), we compared the molecular pathways deregulated in the two series of tumors, and we tested the abilities of genes subsets from each signature to qualify postradiotherapy and post-Chernobyl thyroid tumors as radiation-induced tumors compared to sporadic tumors, independently of mode of exposure, dose, and dose rate. Indeed, five genes common to both the postradiotherapy and the post-Chernobyl signature could accurately classify the two series of tumors by a conventional principal component analysis (PCA). Moreover genes belonging to each signature (postradiotherapy or post-Chernobyl signature) could cross-classify the series of tumors (post-Chernobyl or postradiotherapy series respectively). This highly suggests that a common core of genes are deregulated in radiation-induced tumors whatever external exposure versus internal contamination and thus independently of doses and dose rates.
Materials and Methods
Post-Chernobyl papillary carcinoma series
The post-Chernobyl PTC series described in Table 1 included 14 sporadic PTC (sPTC) from French patients with no history of irradiation, and 12 post-Chernobyl PTC (cPTC) from the Chernobyl Tissue Bank (CTB). Evaluation of the dose at the thyroid for these tumors was provided by the CTB. The complete dosimetry report with calculation of the dose using deterministic or stochastic procedure is given in Supplementary Data 1 (Supplementary Data are available online at
Data on the tumors of the post-Chernobyl tumor series [(9), Chernobyl Tissue Bank]. Boldface indicates tumors included in the learning/training set for the identification of the discriminating 106-gene signature [(11); Materials and Methods].
PTC, papillary thyroid carcinoma, classical papillary variant; PTCFV, papillary thyroid carcinoma, follicular variant; PTCSV, papillary thyroid carcinoma, solid variant; S, sporadic; R, radiation-induced; n.g.a., no gene alterations [BRAF V600E, KRAS, NRAS, HRAS alterations, or PTC rearrangement (9)]. Dosimetry data released from the Chernobyl Tissue Bank: AM, arithmetic mean resulting from 1000 stochastic dose estimates for each subject; SD, standard deviation of the distribution of the 1000 stochastic dose estimates.
This tumor series was previously used to identify the 106-gene discriminating transcriptomic signature of radiation exposure by 131I contamination in the thyroid (11), by reanalyzing the data set of Detours et al. (9). To search for this signature, a learning/training set (see Materials and Methods: Identification of the discriminating molecular signatures of tumor etiology), including seven sPTC and six cPTC, was designed so that the age at tumor diagnosis of sPTC and cPTC was matched as well as possible, and was roughly balanced in terms of sex and genetic alterations between the sPTC and cPTC subgroups.
In the present study, the whole tumor series was used to assess the cross-classification potential of the postradiotherapy 322-gene subset.
Postradiotherapy papillary carcinoma series
The French postradiotherapy PTC series described in Table 2 included 15 sPTC from patients with no history of radiation exposure, and 12 PTC developed in the field of a radiotherapy treatment for a primary cancer (rPTC). Irradiation dose received at the thyroid was estimated for all but one patient.
Tumors are subsets of series previously analyzed by Ory et al. (10). All patients were Caucasian except for one who was a Black African. Boldface indicates tumors included in the learning/training set for the identification of the discriminating 322-gene signature [(10); Materials and Methods].
NR, not relevant; IF, incidental finding; Sc, screening; U, unavailable.
This series was included in a larger tumor series, including PTC and follicular thyroid adenomas either sporadic or radiation-induced, which was used to identify the 322-gene discriminating transcriptomic signature of external radiation exposure (10). Radiation-induced tumors included in this series met Cahan's criteria (16): (i) after radiotherapy, the thyroid tumor must arise in the irradiated field and be proved histologically; (ii) a latent period of several years must have elapsed between radiation exposure and development of the tumor; and (iii) the thyroid tumor must be histologically different from the primary tumor. Moreover, only patients treated before the age of 15 years (period of high thyroid radiation sensitivity) were included in the learning/training tumor set (see Materials and Methods: Identification of the discriminating molecular signatures of tumor etiology).
In the present study, this PTC series was used to assess the cross-classification potential of the post-Chernobyl 106-gene subset.
Identification of the discriminating molecular signatures of tumor etiology (prediction)
Identification of the discriminating signatures using the EMts_2PCA method (10,11), relies on a learning/training step that selects sets of candidate genes whose expressions discriminate between the sporadic and radiation-induced subgroups (learning) and evaluates the potential of the candidate genes to classify new tumors (training).
To maximize the selection of genes that will specifically discriminate the etiology, the design of the learning/training tumor set is critical: (i) only highly probable either sporadic or radiation-induced tumors are considered, (ii) the effect of any confirmed or likely confounding factor (age at diagnosis, sex, gene alteration, histological type) should be minimized for the gene selection. This can be done in two ways: (i) by matching one parameter between the two subgroups, or (ii) by the learning/training process itself, which is based on combinatorial matrices (four radiation-induced tumors vs. four sporadic tumors) built from the learning set (at least 12 tumors). All combinatorial matrices that may introduce a bias in the gene selection, for example four BRAF sPTC versus four rPTC bearing no mutations, were systematically discarded.
Then, the learning process will select, in each combinatorial matrix, candidate genes similarly expressed in each subgroup and differently expressed between the subgroups (sub-signature), the remaining tumors of each combinatorial matrix being used as training tumors. A sub-signature is retained only if it correctly classifies at least one training tumor but never misclassifies any training tumors of each combinatorial matrix; other tumors could be unclassified. Then, validated subsets are compiled in a unique discriminating final signature that includes genes found in at least 70% of the validated sub-signatures. This final signature is then validated by the blind classification, case-by-case, of an independent set of tumors (testing tumors).
In addition to the discriminating signature, the genes of each validated sub-signature not retained in the final signature could be considered as putative deregulated genes (extended list), and thus they were used to get an overview of the molecular pathways deregulated in radiation-induced tumors. Pathways deregulated in the radiation-induced tumors as compared with sporadic tumors were built based on signaling and metabolic KEGG pathways and on published papers with special attention regarding thyroid tumorigenesis, response to radiation exposure, and redox status.
Cross-analysis of postradiotherapy and post-Chernobyl signatures (classification)
Postradiotherapy and post-Chernobyl tumors were hybridized on different microarrays: human 25K oligo-microarrays from a national genomic platform (Resogen Program, RNG/MCR, Evry, France, ArrayExpress accession number E-MEXP-2739), or human 1 cDNA microarray slides (Agilent Technologies, Massy, France, GEO database accession number GSE3950) respectively. Consequently, the comparison between the 322-gene postradiotherapy and the 106-gene post-Chernobyl signature was possible only when the probes could be analyzed in the two series (significant measurements for all tumors of both series). The gene subsets of each postradiotherapy and post-Chernobyl discriminating signature were used to cross-classify the postradiotherapy and the post-Chernobyl PTC series respectively. Tumor classification was done by conventional PCA (17).
Results
Comparison of the post-Chernobyl and postradiotherapy signatures and cross-classification of postradiotherapy and Chernobyl tumors respectively
An overlap of five genes (PABPC1, SERPINE1, GTF2H2, DHCR24, and CLU) was found between the 322-gene postradiotherapy and the 106-gene post-Chernobyl signatures. This overlap was discriminant enough to separate either the postradiotherapy PTC (rPTC) (Fig. 1A and C) or the post-Chernobyl PTC (cPTC) series (Fig. 1B and D) from their respective sporadic PTC series (sPTC) into two groups as a function of etiology with one error in each series. The classification as sporadic and radiation-induced using the five common genes was observed in the postradiotherapy (Fig. 1A) and post-Chernobyl (Fig. 1B) learning/training tumors subsets (see Tables 1 and 2, and Materials and Methods: Corresponding to the tumors used to identify the respective signatures). Interestingly, these five genes also classified in two groups independent series of postradiotherapy (Fig. 1C) and post-Chernobyl (Fig. 1D) tumors (testing tumors).
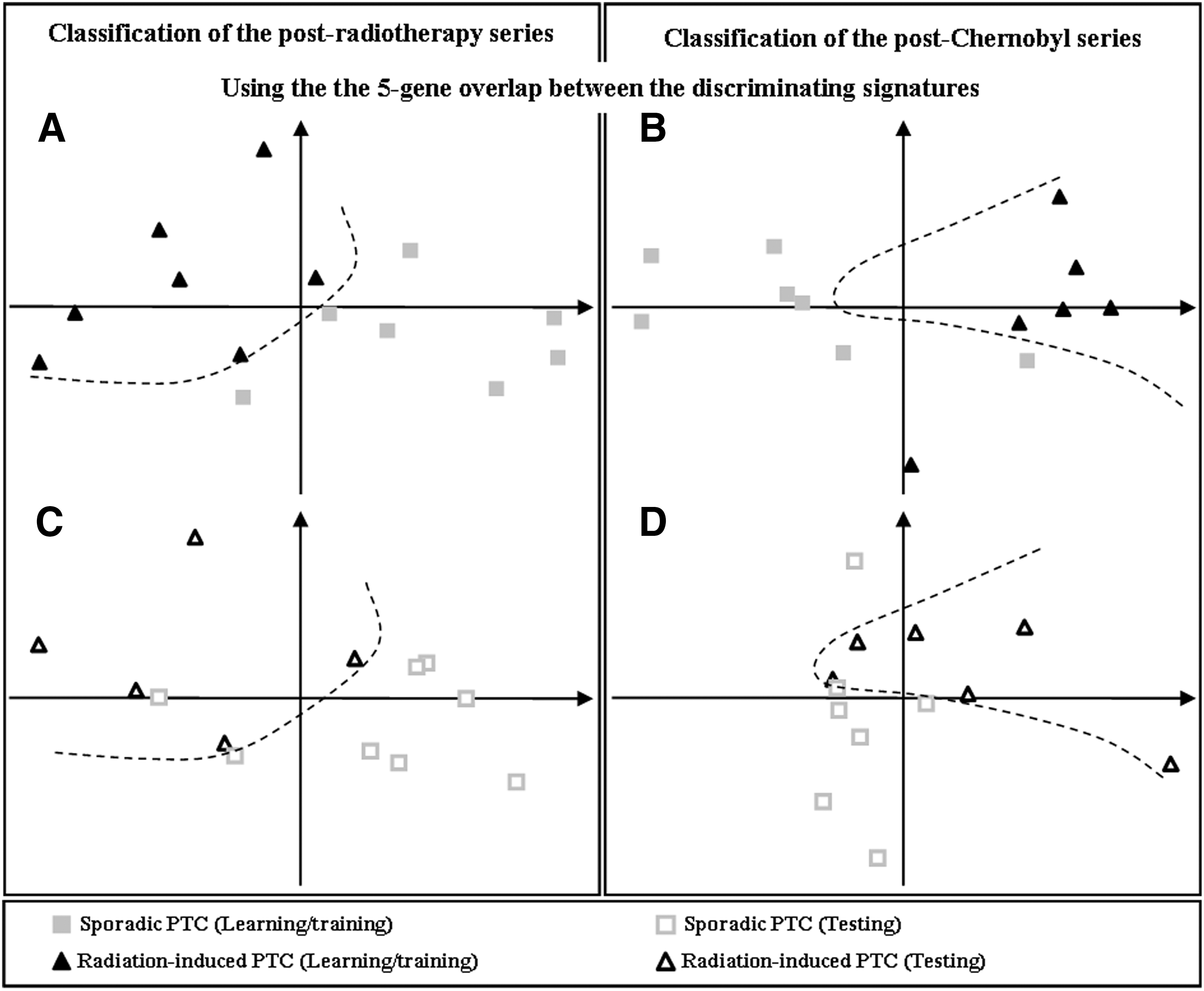
Principal component analysis (PCA) classification of the postradiotherapy and the post-Chernobyl series using the five-gene overlap between the discriminating signatures. Two-dimensional plots of the spatial distribution of the tumors as a function of the two eigenvectors (with greater eigenvalues) of PCA considering the five-gene overlap between the postradiotherapy signature and the post-Chernobyl signature, for the postradiotherapy PTC series
Next, we wished to assess whether discriminating markers identified in a context of either external radiation exposure (postradiotherapy) or internal radiation exposure (post-Chernobyl) could cross-classify either the post-Chernobyl or the postradiotherapy PTC respectively versus sporadic PTC. Unfortunately, the two series were not hybridized on the same microarrays. Consequently, the 322 genes of the postradiotherapy signature could be associated with 145 probes on the post-Chernobyl array, and the 106 genes of the post-Chernobyl signature could be associated with 96 probes on the postradiotherapy array. Moreover, since for clustering we need significant measurements for all tumors, only 18/106 probes corresponding to 16 genes could be used to classify the postradiotherapy series (Supplementary Data 2), and 37/145 probes corresponding to 32 genes for the post-Chernobyl series (Supplementary Data 3). As shown in Figure 2, both the 16-gene post-Chernobyl subset and the 32-gene postradiotherapy subset could cross-classify the post-Chernobyl or the postradiotherapy PTC respectively, as compared with their respective series of sPTC. Only one rPTC was misclassified (Fig. 2A), and the post-Chernobyl series was perfectly separated into two groups (Fig. 2B).
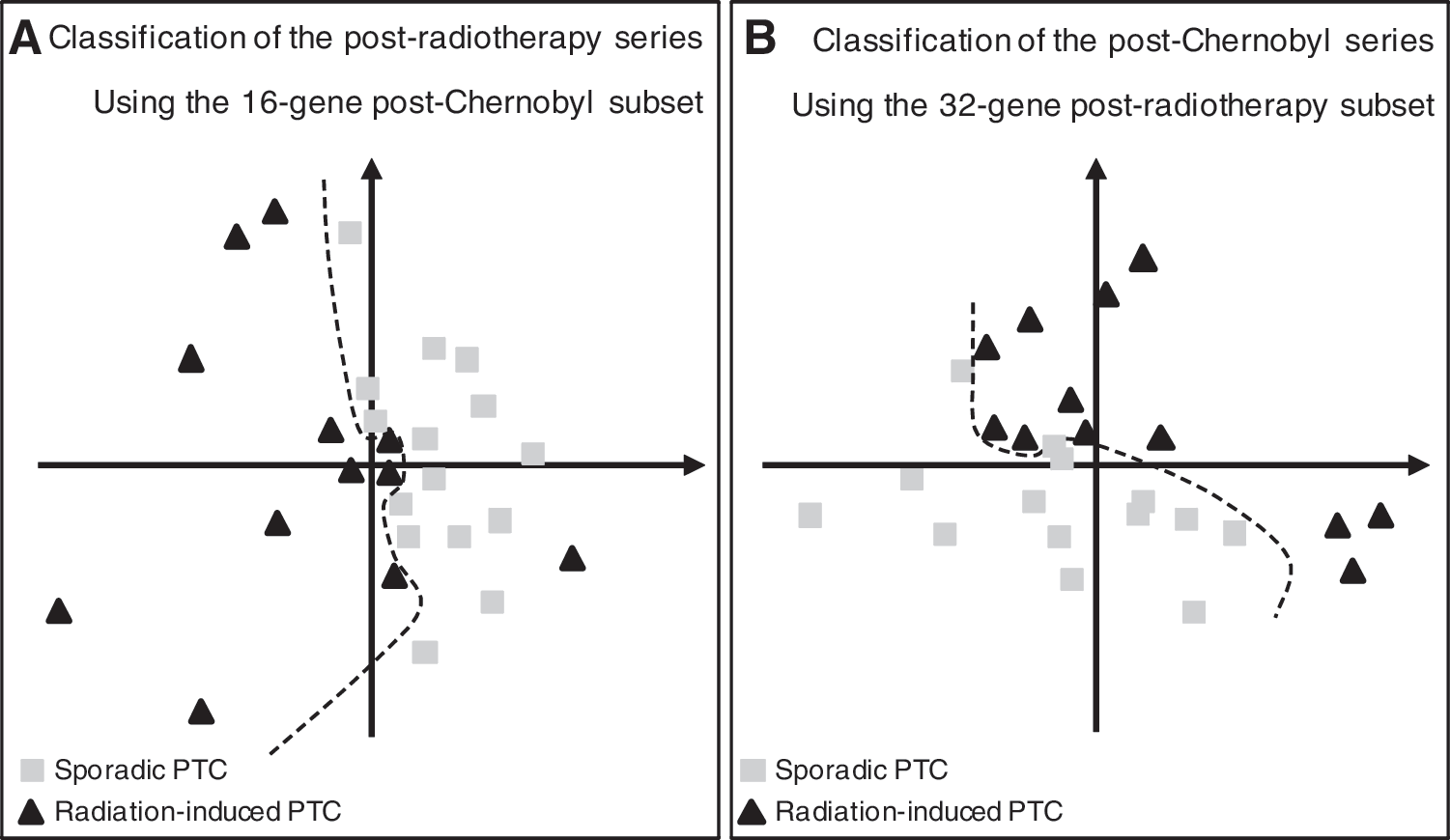
Cross-classification by PCA of the postradiotherapy and the post-Chernobyl series of tumors compared with sporadic tumors. Two-dimensional plots of the spatial distribution of the postradiotherapy PTC series
Analysis of molecular mechanisms deregulated in post-Chernobyl tumors
Molecular analysis of the 106-gene signature indicated that DNA damage response, mitochondria, p53 functions, and hypoxia response/angiogenesis regulation are mainly deregulated in cPTC compared with sPTC (Fig. 3). To compare the molecular mechanisms deregulated in post-Chernobyl tumors with those identified in postradiotherapy tumors (10), and to be as exhaustive as possible in the analysis of these molecular pathways, we considered the 106 genes of the discriminating signature [Supplementary Data 4; (11)] and the 2164 preselected probes (corresponding to an extended list of 1901 genes) found to be deregulated in cPTC (training step; Supplementary Data 5). This more complete overview showed that repair of DNA damage (Supplementary Data 6), regulation of cellular oxidative stress status and oxidative stress signaling, including mitochondrial function (Supplementary Data 7), and the response to misfolded protein, including the mTOR, endoplasmic reticulum stress, and autophagy pathways (Supplementary Data 8), are deregulated in both postradiotherapy and post-Chernobyl tumors.
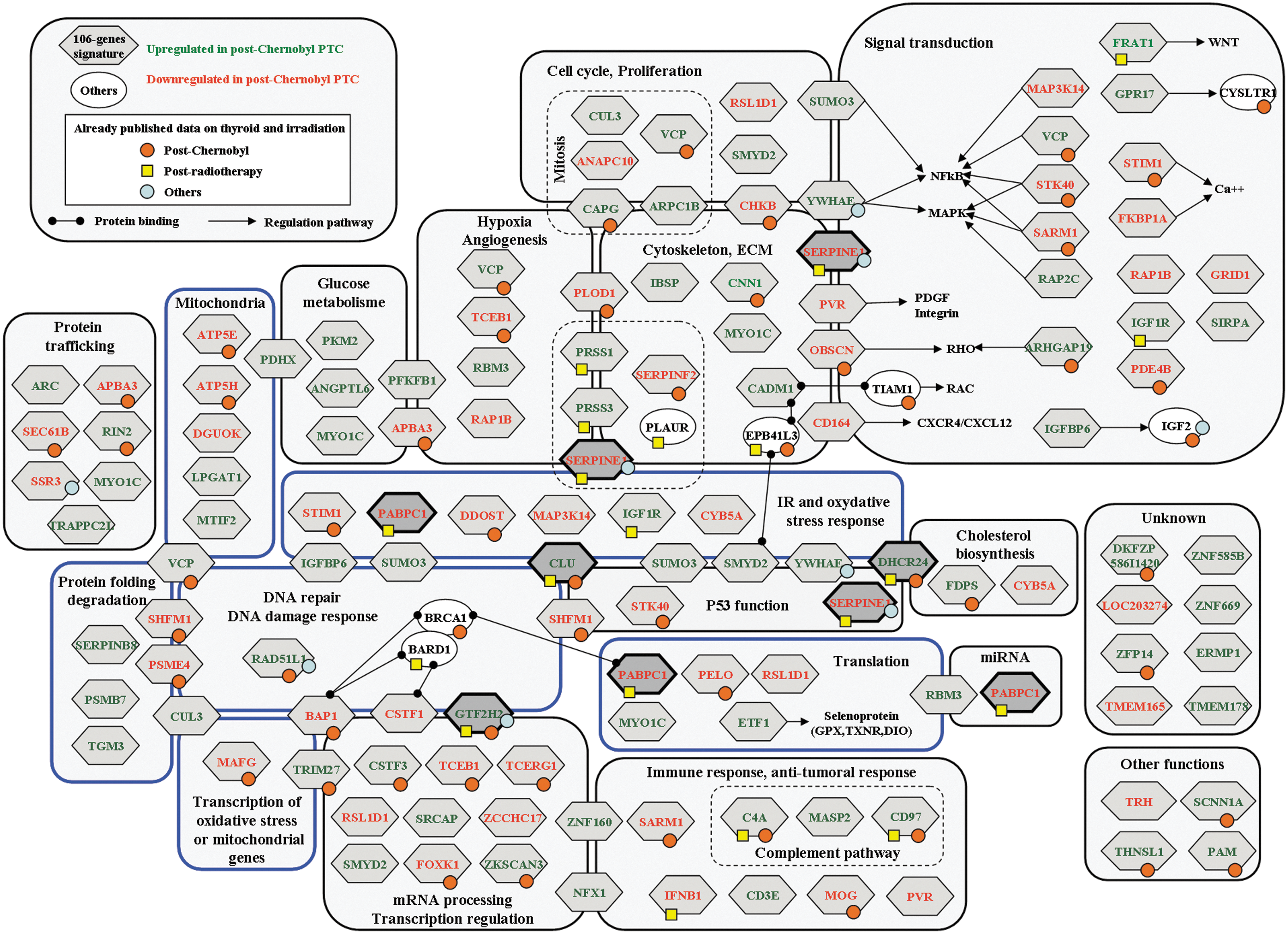
The 106-gene post-Chernobyl signature classified by functional categories. Darker diamonds highlight the five genes common to both the postradiotherapy signature and the post-Chernobyl signature. Other symbols refer to already published deregulated genes (Supplementary Data 4 for references): in postradiotherapy thyroid tumors (yellow squares), post-Chernobyl thyroid tumors (orange circles), or in short-term response to radiation (blue circles). Blue boxes, pathways also deregulated when analyzing the extended list (Materials and Methods and Supplementary Data 6–8).
Discussion
Confidence in the series of samples is critical when searching for molecular signatures of thyroid tumor etiology. Sporadic thyroid tumors of both the Chernobyl [Table 1; (9)] and radiotherapy [Table 2; (10)] series developed in French patients without a history of radiation exposure. People living in France are exposed to natural background radiation in the range of 2–3 mSievert/year. There is still no consensus on the existence of a radiation threshold that triggers carcinogenesis, but if such a risk exists, at very low doses and dose rate, it still depends on the dose. This means that, assuming the linear nonthreshold relationship between the dose and cancer risk, with a risk of 5% per Gy (at high dose rate), one can assume that the risk of having radiation-induced tumors in the sporadic series is 0.00125%. Thus, the sporadic series could be used with confidence as the control group when searching for a molecular signature of thyroid tumor etiology. All postradiotherapy tumors met Cahan's criteria [Materials and Methods; (16)]. The estimated dose delivered to the thyroid ranged from 12 to 43 Gy (learning/training tumors), and only patients treated before the age of 15 years (the period of greatest radiation sensitivity of the thyroid) were included in the learning/training tumor set. Thus, the likelihood that postradiotherapy tumors have been radiation-induced is very high, and this is confirmed by the identification of a robust discriminating signature of tumor etiology with a sensitivity of 92% (10). Post-Chernobyl tumors in the Detours et al. study (9), obtained from the CTB, were from patients living in the more contaminated oblasts (administrative areas) of Ukraine (18,19), aged between 1 and 16 years at the time of the Chernobyl accident, and with a latency of 15–17 years for tumor development. When identifying the 106-gene signature from the microarray data set of Detours et al. (9), the proportion of radiation-induced tumors in this series was estimated to be around 70% according to the age at exposure and taking into account the latency for tumor development (19). Consequently, fewer than 1.8 radiation-induced tumors could be sporadic in the series. For this reason, we applied our previously described methodology, which allowed us to find a predictive signature for blind classification, even if a wrongly labeled tumor is included in the learning/training tumor set and doesn't find a signature when more than one wrongly labeled tumor is present (11). Meanwhile, the subproject of the CTB, which was dedicated to link the large number of dose measurements with the CTB cases, reached completion (mid-2012), and thyroid exposure dose estimation was thus obtained from the CTB. In this series, for half of the post-Chernobyl tumors, the received dose at the thyroid was estimated to be less than 0.1 Gy, which is the limit for which a significant increased risk of thyroid cancer is observed by conventional epidemiology (Table 1). Consequently, the 106-gene signature was identified with a learning/training group of post-Chernobyl tumors from patients who received radiation doses estimated at between 0.014 Gy and 2.5 Gy. This signature was validated by blindly classifying an independent series of seven sporadic tumors and six post-Chernobyl tumors developed in children exposed at doses ranging from 0.023 Gy to 1.7 Gy between the ages of one and five years, the highest period of thyroid sensitivity. Regarding the dose received by children included in the validation group, it is highly probable that these tumors have been radiation-induced. The 106-genes signature well classified, as sporadic or radiation-induced, 12 out of these 13 tumors, one sporadic tumor (PTC18) was unclassified, but not misclassified. It indicates that tumors developed in children exposed at a radiation exposure dose of a magnitude of 10 mGy, present common molecular imprinting, with radiation-induced tumors developing after exposure at higher radiation doses. If confirmed by studying larger series of tumors, our data open the way for an informative molecular epidemiology to estimate the risk of thyroid cancer after radiation exposure at doses lower than 100 mGy.
Thus, using a unique microarray analysis strategy, we found a 322- and a 106-gene signature discriminating postradiotherapy (acute high doses of external exposure) and post-Chernobyl (lower doses after 131I contamination) thyroid tumors compared with sporadic tumors (10,11). Both external exposure and 131I contamination can induce thyroid tumors, but most of the published studies analyzed post-Chernobyl tumors, and the relevance of extrapolation of conclusions from the data on tumors occurring after external radiation exposure to post-Chernobyl tumors that occurred after internal 131I contamination has still not been investigated. In this study, we explored the molecular similarities between tumors developed after external exposure and 131I contamination by comparing molecular pathways deregulated in the two tumor series and by evaluating the ability of gene subsets from each signature—the postradiotherapy signature or the post-Chernobyl signature—to qualify postradiotherapy and post-Chernobyl thyroid tumors as radiation-induced tumors compared to sporadic tumors, independently of mode of exposure, dose, and dose rate.
Molecular analysis of post-Chernobyl tumors and comparison with postradiotherapy tumors
Molecular analysis of the 106-gene signature indicated a general scheme of cellular oxidative stress response (Fig. 3). Forty of these 106 genes were reported in the study by Detours et al. (9) [(9); Supplementary Data 4], and three genes (CD97, ARHGAP19, and DKFZP586I1420) were found to be deregulated in cPTC (8,12).
To get a more exhaustive overview of the pathways deregulated in post-Chernobyl tumors, we also analyzed an extended list of 1901 genes found to be deregulated in cPTC as compared with sPTC during the training step (Supplementary Data 5). Although suitable for classifying the tumors according to their etiology, these genes were not retained in the final signature because they were not discriminating enough to predict the etiology of independent tumors (Materials and Methods). As cPTC and sPTC could be classified by using a signature discriminating lymphocyte responses to ionizing radiation and to H2O2 (9), it was suggested that cPTC may be associated with a pattern of radiation response and DNA repair, and sPTC may be associated with a pattern of oxidative stress due to deregulation of thyrocyte metabolism during sporadic tumorigenesis. However, both cPTC and rPTC exhibited gene deregulations targeting the regulation of cellular oxidative stress status and signaling (Supplementary Data 7). Radiation-induced thyroid tumors may have specific oxidative status as compared with sporadic tumors, since many genes coding for mitochondrial functions, including the oxidative phosphorylation system, were deregulated (Supplementary Data 7). This may be of consequence for radiation-induced tumorigenesis, as, in parallel, radiation-induced thyroid tumors also exhibited gene deregulations targeting DNA repair pathways and proteasome subunits known to sensitize cells to DNA-damaging agents by regulating the efficiency/rapidity of the DNA damage response [(20,21); Supplementary Data 6]. Increase in reactive oxygen species due to mitochondrial respiratory chain dysfunction also leads to lipid and protein peroxidation, with consequences for apoptosis and autophagy regulation (22). Accordingly, this analysis highlighted the potential involvement of genes coding for the endoplasmic reticulum stress, mTOR, and autophagy pathways in radiation-induced tumors (Supplementary Data 8). Recently, it was shown that induction of autophagy increases the radiosensitivity of thyroid cancer cells (23). In conclusion, without any direct evidence, we can speculate that deregulation of mitochondria, oxidative status, DNA repair, proteasome, and autophagy could contribute to the specific molecular signature of radiation-induced tumorigenesis, as they are deregulated in both cPTC and rPTC compared with sporadic thyroid tumors.
Cross-classification of postradiotherapy and post-Chernobyl tumors using post-Chernobyl and postradiotherapy specific deregulated genes, respectively
The identification of specific signatures of postradiotherapy (external exposure) or post-Chernobyl (internal contamination) tumors led to the question of whether each signature cross-classifies the other tumor series. Given that the two series were hybridized on different arrays, only a limited number of genes of each signature were exploitable for cross-classification: 32 out of 322 genes and 16 out of 106 genes for the postradiotherapy and the post-Chernobyl signatures respectively. Remarkably, despite this small number of genes, each of the 32- and 16-gene subsets permitted cross-classification of either the postradiotherapy or the post-Chernobyl PTC series respectively, as compared with the sporadic tumors (Fig. 2A and B). This illustrates that even if these genes were not discriminating enough to be included in the two signatures for tumor etiology prediction, they are deregulated in both radiation-induced thyroid tumor series (as compared with sporadic tumors) and are efficient for tumor classification. This means that common genes are deregulated whatever the mode of exposure, dose, and dose rate. The 16- and 32-gene subsets, while separating sporadic and radiation-induced tumors, are not discriminating enough for prospective tumor prediction. This result was expected, since the two signatures of 322 and 106 genes were specifically optimized for tumor etiology prediction.
Biological significance of the five genes common to postradiotherapy and post-Chernobyl signatures: the central role of CLU?
Postradiotherapy and post-Chernobyl series were hybridized and analyzed on distinct microarray platforms, and thus there are many biases for full data comparison (signal detection, background treatment, data normalization, etc.). This could explain why very small overlaps are found when comparing published papers on a given pathology. Consequently, when an overlap is found, as in the five genes in the present work, it can be trusted, bearing in mind, however, that genes could have been missed. The five genes (PABPC1, SERPINE1, GTF2H2, DHCR24, and CLU) common to the two signatures, which are sufficient for classifying either cPTC or rPTC versus sporadic thyroid tumors (Fig. 1), could constitute molecular markers of the thyroid gland radiation exposure history, whatever the dose, dose rate, and mode of exposure. However, they are not discriminating enough for a prospective tumor prediction. This was expected, since they belong to both 322- and 106-gene signatures, which were specifically optimized for tumor etiology prediction. These five genes have been reported to be regulated by and/or to participate in radiation and oxidative stress responses and/or DNA repair. SERPINE1 was linked to regulation of angiogenesis and fibrosis after irradiation (24,25). PABPC1, a marker for stress granules, is induced by hydrogen peroxide (26). Both DHCR24/seladin-1 (27) and clusterin (CLU) (28) regulate cell survival after oxidative stress. CLU levels in thyroid cancer patient sera were found to be lower than in disease-free people (29), and could have a central role in radiation-induced thyroid carcinogenesis, since, according to the dose, it could have a pro- or an anti-apoptotic effect after irradiation (30). Further studies are needed to confirm the central role of CLU and to specify which isoforms are expressed in radiation-induced thyroid tumors. A scheme summarizing the hypothetical role of CLU is proposed in Figure 4. Finally, GTF2H2, which is essential for both transcription initiation and nucleotide excision repair, was reported to be upregulated in response to high-linear energy transfer particles exposure in a human thyroid epithelial cell line (31), and SERPINE1 was upregulated after 137Cs and neutron exposure of human thyroid tissue transplanted in SCID mice (32). These two genes, which participate in the short-term response to radiation exposure in different thyroid models, are also hallmarks of the long-term radiation response, since they are also deregulated in radiation-induced thyroid tumors after external acute exposure and after contamination by chronic low doses of 131I.
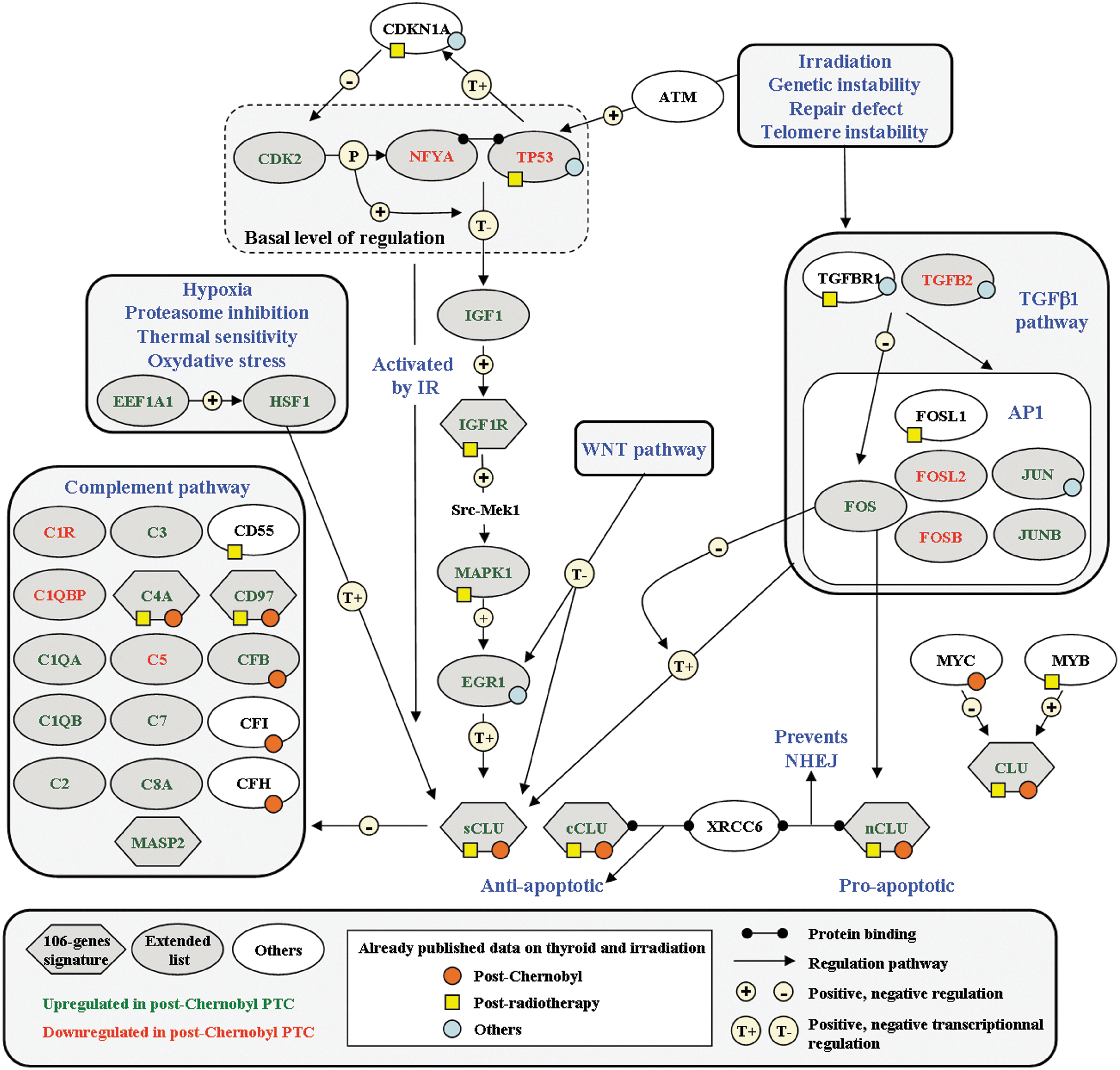
Clusterin regulation in radiation-induced thyroid tumors. CLU induction depends on ATM-dependent inhibition of p53-NFYA transcription factors (28,37,38). WNT pathway, found to be significantly deregulated in postradiotherapy tumors (10), downregulates CLU (39). Interestingly, TGFβ and EGF increase the secretion of CLU and upregulate SERPINE1 during porcine thyrocyte de-differentiation (40). Post-Chernobyl deregulated genes found in the present analysis are indicated as gray diamonds (106-gene signature) or ovals (extended list). All other symbols refer to deregulated genes already published (Supplementary Data 4 and 5 for references): yellow squares or orange circles to postradiotherapy or to post-Chernobyl thyroid tumors respectively, and blue circles to short-term response to radiation. sCLU, cCLU, and nCLU, secreted, cytoplasmic, and nuclear CLU respectively; IR, irradiation; NHEJ, nonhomologous end-joining.
Radiation-induced thyroid tumor signature: genetic susceptibility or radiation exposure?
It has been suggested that the molecular specificities of cPTC versus sPTC could mirror a pattern of genetic susceptibility to radiation-induced carcinogenesis rather than specific radiation-induced mechanisms of tumorigenesis (14,33). Strikingly, it was reported that GC heterozygosity at the miRNA146a polymorphism locus is associated with an increased risk of developing PTC. Moreover, this heterozygosity impacted the regulation of the transcriptome in both normal and thyroid tumor cells as compared with homozygote GG samples (34), and part of these miRNA146a signatures overlap with the post-Chernobyl deregulated genes. Interestingly, miRNA146a is one of the miRNAs reported to be upregulated in the radiation response of thyrocytes (35). Overall, these data suggest that miRNA146a could be a good candidate to modulate radiation-induced thyroid tumorigenesis through thyroid tissue response. Alternatively, the molecular pattern found in radiation-induced thyroid tumors could also be the hallmark of a history of radiation exposure independently of cancer development, since several of these genes are involved in the short-term radiation response [(31,32,36); Supplementary Data 4 and 5]. In the latter case, a specific gene expression signature could be found by comparing normal thyroid tissue surrounding radiation-induced thyroid tumors with normal thyroid tissue surrounding sporadic thyroid tumors, and it should share similarities with signatures of radiation-induced tumors.
Overall, our results show that (i) the five genes common to both signatures classified postradiotherapy and post-Chernobyl tumors from their respective series of sporadic tumors (Fig. 1); (ii) at least part of the postradiotherapy signature classified radiation-induced post-Chernobyl tumors, and, reciprocally, part of the post-Chernobyl signature classified postradiotherapy tumors (Fig. 2); and (iii) molecular pathways deregulated in each series of cPTC and rPTC are associated with cellular response to radiation and oxidative stress. These results suggest that postradiotherapy (external exposure) and post-Chernobyl thyroid tumors (131I contamination) display a common core of molecular specificity, which may lead to the identification of a “general” radiation-induced signature whatever the genetic background, since the two sporadic series paired to either postradiotherapy and post-Chernobyl tumors present the same genetic background.
At present, we cannot draw firm conclusions regarding the causality of the molecular specificities found in radiation-induced thyroid tumors in terms of genetic susceptibility to radiation sensitivity and/or to thyroid cancer, or regarding the radiation-induced specific mechanisms of carcinogenesis. However, we found that thyroid tumors that developed after acute external exposure to high doses of radiation shared gene expression specificities with thyroid tumors that developed after internal contamination at chronic low doses.
Footnotes
Acknowledgments
This work was supported by the European Union 6th Framework GenRiskT, Electricité de France and the European DoReMi framework. The authors gratefully acknowledge the confirmation of diagnosis provided by the following members of the International Pathology Panel of the Chernobyl Tissue Bank: A. Abrosimov, T.I. Bogdanova, M. Ito, V. LiVolsi, J. Rosai, and E.D. Williams.
Author Disclosure Statement
No competing financial interests exist.