
Abstract

The active principle(s) in the thyroid gland
In 1896, Baumann discovered that the thyroid gland contained considerable amounts of iodine (4). In fact, this was not a great surprise since the association of goiter with iodine deficiency had already been recognized, and iodine was known to be a remedy for some goiters (5). Baumann then found that most of the iodine was contained in a protein fraction which on hydrolysis yielded a compound he called thyrojodin, iodothyrin in English (6). Furthermore, he demonstrated that iodothyrin was effective in treating myxedema, and reversing the effects of thyroidectomy in animals (6). Eighteen years later, Kendall isolated and crystallized a small amount of a physiologically active compound from the thyroid that he named thyroxin (7). By 1926, Harington had greatly improved the extraction method to increase the yield and was able to characterize and subsequently synthesize the compound. He renamed it thyroxine (T4) (8,9).
The circulating thyroid hormone
Before 1948, the nature of the circulating thyroid hormone was an enigma. Was it T4, thyroglobulin, or something else? Although T4 was known to ameliorate the symptoms of hypothyroidism when administered to humans or animals, in general investigators were hesitant to assert that it was the circulating hormone for several reasons. First, it was not possible to account completely for the biological activity of thyroglobulin by its T4 content. Second, it has a long latent period of action when injected into animals; at that time it was widely assumed that hormones when injected would rapidly induce a response. Third, T4 was inactive in in vitro systems. In 1935, Harington, on the basis of his extensive studies concerning the iodine compounds in the thyroid gland, postulated that the circulating thyroid hormone was a peptide consisting of T4 and diiodotyrosine (10). However, by 1944 he had re-evaluated the evidence and changed his mind deciding that this peptide “represents an unnecessary complication and one which is not justified by the facts,” and he hypothesized that the circulating hormone was in fact T4 (11).
Proving this hypothesis was soon facilitated by the advent of three major technical developments: the availability of radioactive iodine (131I) for research purposes, refined methods for determining small quantities of iodine in tissues, and paper chromatography for the separation of radioactive compounds.
Using two of these techniques, Taurog and Chaikoff obtained strong evidence that in normal animals the circulating thyroid hormone consists of T4 bound reversibly to plasma proteins. They demonstrated that the protein-bound iodine (PBI) in serum behaves like added T4 with respect to extraction with organic and inorganic solvents and its ability to be dialyzed. In addition, the PBI in rat plasma labeled in vivo with 131I follows the T4 carrier quantitatively when the latter is crystallized repeatedly or when it is distributed between two immiscible solvents (12). That the majority of this labeled fraction did indeed comprise T4 was subsequently demonstrated unequivocally using paper chromatographic analysis by Laidlaw (13) and shortly thereafter by Taurog et al. (14), Rall (15), and Gross et al. (16).
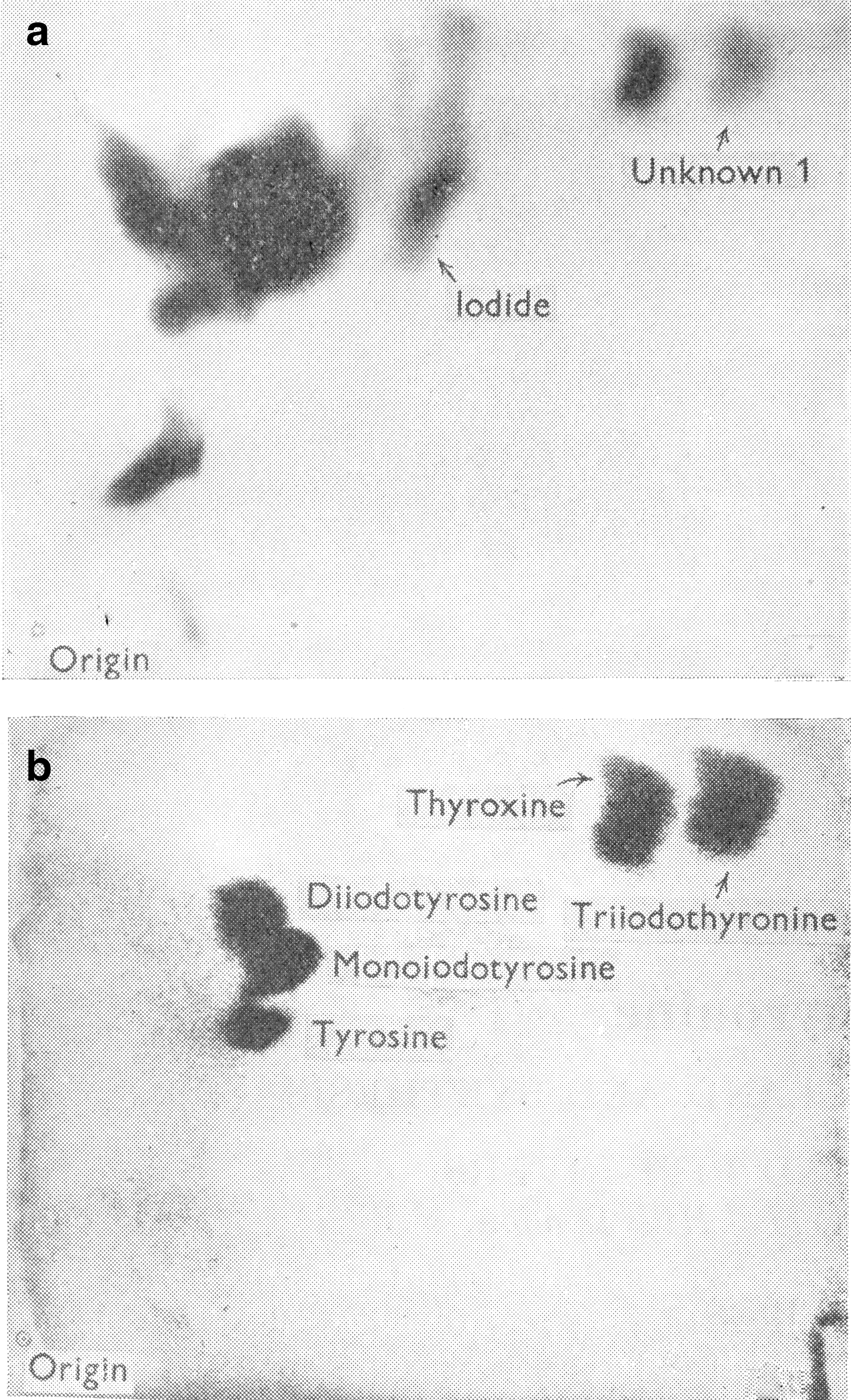
Identification of Unknown 1
The serendipitous outcome of the latter study was the discovery of Unknown 1. Gross et al. (16) were trying to identify the various iodinated amino acids present in plasma and in an extract of unhydrolyzed thyroid gland obtained from iodine-deficient rats injected with carrier-free 131I. The radioactive compounds were separated from each other by two-dimensional chromatography and their location on the paper determined by exposure of the paper to X-ray film. Some compounds were identified by co-chromatographing T4 and several iodotyrosine compounds. After visualizing these carriers on the chromatogram by appropriate staining, their locations were compared with the locations of the radioactive spots as illustrated in Figure 1 (17). Mono- and diiodotyrosine, T4 and iodide were identified in the thyroid extract, and T4 was clearly evident in plasma. In addition, several other compounds were present in the thyroid, and one of them, designated Unknown 1, was also present in plasma (16). In a subsequent study, in which the iodinated compounds present in thyroid and plasma from iodine-deficient and iodine-sufficient rats were compared, it was noted that Unknown 1 was only evident in plasma in the iodine-deficient rats (18). The authors used iodine-deficient rats in the original study to increase the specific activity of the iodinated compounds produced. However, with respect to the detection of Unknown 1, this was a fortuitous decision since it is now well known that the T3/T4 ratio in the thyroid gland increases greatly when iodine supplies are limited.
In 1950, Dr. Jack Gross joined the laboratory of Dr. Rosalind Pitt-Rivers at the National Institute for Medical Research in London, United Kingdom (Fig. 2). The Director of the Institute was Sir Charles Harington, Pitt-Rivers' mentor and an eminent chemist who had been the first to synthesize T4. Together, their primary goal was the identification and characterization of Unknown 1. First, they demonstrated the presence of Unknown 1 in the plasma of humans who had received therapeutic doses of radioactive iodine (19). Next, they synthesized 3,5,3′-triiodothyronine (T3) from 3,5-diiodothyronine (17) and used it to demonstrate that the Unknown 1 present in human plasma (20) and in hydrolyzed thyroid tissue from rats given 131I (16) was indistinguishable chromatographically from T3 (17). Finally, they isolated T3 from ox thyroid tissue and demonstrated that its X-ray crystallographic characteristics were identical to those of synthetic T3 (17).

Drs. Rosalind Pitt-Rivers and Jack Gross at the National Institute for Medical Research, Mill Hill, London, 1957 (photo by V.A. Galton).
As so often happens with major advances in research, Gross and Pitt-Rivers were not alone in their pursuit of T3. Concurrently with their investigations, Roche, Lissitsky, and Michel, in France, also synthesized T3 and used it to identify radioactive T3 in the hydrolyzed thyroid tissue from rats given 131I (21,22). Although somewhat different methods were used in the two studies, the results obtained in each study provided mutual confirmation.
Gross and Pitt-Rivers reported that T3 was three times more potent that T4 in a goiter prevention assay, a finding that led them to speculate that T3 was the active form of the thyroid hormone (23). In an expanded study, they demonstrated T3 to be more active than T4 in a wide variety of tests and again suggested that T3 was the active hormone and T4 the precursor (24).
Is T3 the active form of the thyroid hormone?
Although the hypothesis that T3 is the active form of the thyroid hormone was an attractive one, proof was not immediately forthcoming. Several key questions remained unanswered, including: 1. Is T4 converted to T3 in peripheral tissues and, if so, to what extent? 2. What are the relative contributions of T3 to overall thyroid hormone action? 3. Is the action of T4 dependent on its conversion to T3?
Indeed, the 1960s proved to be a decade of disillusionment and confusion with respect to the concept that T3 was the active form of the thyroid hormone. As early as 1955, Pitt-Rivers began to have doubts on this issue, due primarily to the fact that while rats responded more rapidly to T3 than to T4, the latent period was still very long, measured in hours rather than minutes (Galton, personal communication). She was also concerned that T3 appeared to be inactive in vitro. Arguably a more significant problem was that, despite several attempts, the presence of [131I]T3 following injection of [131I]T4 into athyreotic humans could not be demonstrated unequivocally (25,26). Attempts to demonstrate T4-to-T3 conversion in vitro were also largely unsuccessful. Although Albright et al. did obtain some conversion of [131I]T4 to [131I]T3 in both rat and human kidney slices (27,28), these findings were not confirmed (29) and a plethora of other groups were unable to demonstrate [131I]T3 production from [131I]T4, despite the fact that deiodination of T4, as evidenced by the release of 131I, clearly occurred (30). By 1962, Pitt-Rivers had rejected her own hypothesis and she outlined her reasons succinctly in a lecture given at the University of Tennessee in Memphis in 1962 (a digital recording of which is in on file with the American Thyroid Association). First, peripheral conversion of T4 to T3 has not been unequivocally demonstrated. Second, injected [131I]T3 disappears more rapidly from the body than does [131I]T4. This would not be predicted if T3 was the active form of T4. Third, T3 is not effective in in vitro systems. Fourth, since a significant amount of T3 per se is normally secreted from the thyroid, T4-to-T3 conversion is not necessary and may be redundant. In her own words she concludes “So much for the idea that T3 is the active hormone; we can dispose of that theory now and concentrate on T4.” In fact, she felt that both T4 and T3 contribute to thyroid hormone action.
The turning point for T3 came in the early 1970s and it was again largely due to three major technical break throughs. First, the development of competitive binding assays, including radioactive immunoassays for thyroid hormones. Second, the recognition that there are specific enzymes that convert T4 to T3 and the development of sensitive methods for their assay. Third, the discovery of the thyroid hormone receptors and the development of techniques to study their properties.
In 1970, using highly sensitive techniques, which included thyroid hormone displacement technology, Braverman et al. demonstrated unequivocally that T4-to-T3 conversion occurs in athyreotic humans (31). In the same year, Sterling et al. provided additional proof by demonstrating the production of [14C]T3 in euthyroid human subjects injected with [14C]T4. They estimated that as much as a third of the extrathyroidal T4 that was metabolized was converted to T3 (32). These results laid to rest one major contraindication to the theory that T3 was the active form of T4.
Soon after this, it was found that T4-to-T3 conversion could be readily demonstrated in vitro. Between 1955 and 1976, the study of T4 deiodination in vitro was hampered, especially in broken cell preparations, because considerable nonspecific, heat-stable deiodination occurred in a reaction that did not produce detectable amounts of T3 (30). However, the problem was resolved in 1976 by the demonstration that thiol groups were essential cofactors for the deiodinases, and thiol-containing compounds, such as dithiothreitol (DTT), needed to be included in broken cell preparations to support the enzymatic deiodination. The presence of these compounds also greatly reduced the nonenzymatic breakdown of T4. Thus, it was shown that in the presence of DTT, subcellular fractions of rat liver can readily convert T4 to T3 (33).
The finding that T4 can be converted to T3 in extrathyroidal tissues is certainly compatible with the concept that T3 is the active form of the thyroid hormone. Nevertheless, it does not prove it. While the physiological actions of T4 might be dependent on its conversion to T3, it is possible that both T4 and T3 contribute to overall thyroid hormone action, in which case follow-up questions concern the relative contribution of the two hormones and whether their actions mimic or complement each other?
The identification and characterization of the thyroid hormone receptors has provided extremely strong evidence that T3 is indeed the major player in thyroid hormone action. These receptors, located in the nucleus, were first reported in 1972 by Oppenheimer et al. (34) and Schadlow et al. (35), and they were found to have a much higher affinity for T3 than for T4 (36,37). Tata et al. had discovered previously that T3 can regulate gene transcription (38), an effect now widely accepted to be the primary mechanism of thyroid hormone action. Thus, the finding that more than 80% of the total iodothyronine bound to the nuclear receptors in liver and kidney from euthyroid rats was T3, strongly substantiated a major role of T3 in thyroid hormone action (39). Indeed, there appears to be no justification for denying that T3 is responsible for the majority of thyroid hormone action at the level of gene expression.
However, the question of whether T3 is responsible for all of thyroid hormone action and whether T4 functions only as a prohormone has yet to be answered. Supporting this concept are several published observations indicating that 6-n-propylthiouracil antagonizes the physiological effects of T4, but not of T3; this has been shown to be due to a marked reduction in the fractional rate of deiodination of T4 to T3 (40). In addition, it has been demonstrated that the suppression of pituitary TSH secretion that occurs following administration of T4, is due to its intrapituitary conversion to T3 (41). Nevertheless, some of the receptor-bound hormone in tissues is T4 (39), and it is possible that the T4/receptor complex can also modulate gene transcription and, thereby, contribute to total thyroid hormone action. If so, then it would be of considerable interest to know whether it merely mimics the effects of T3 and/or has some specific effects of its own?
In addition to the genomic actions of the thyroid hormone, a number of thyroid hormone effects have been described that occur rapidly, are unaffected by inhibitors of transcription and translation, and involve extra nuclear systems. The mechanisms of several of these nongenomic effects are understood, at least in part, and involve either plasma membrane receptors or nuclear receptors located in the cytoplasm [see review (42)]. The physiological relevance of these findings is, however, still controversial. Although T3 can initiate many of these effects, it appears that in some cases T4 is the preferred hormone, and in these instances, it is acting as a hormone rather than a prohormone (42).
In summary, the history of T3 began in 1950 when a radioactive spot than did not correspond to any known iodothyronine or iodotyrosine was noted on an autoradiograph of a two-dimensional chromatogram of thyroid extract from a rat injected with 131I. Two years later the spot was identified as T3 and, primarily because it was more physiologically active than T4, it was postulated to be the active form of the thyroid hormone. Proof of this was not immediately forthcoming and it was not until the early 1970s that it was accepted that T3 is responsible for most of the direct genomic effects of the thyroid hormone. Future studies will determine if it is responsible for all the genomic effects and whether it plays an equally dominant role in initiating the nongenomic actions of the thyroid hormones.
Footnotes
Disclosure Statement
The author has nothing to disclose.