
Abstract
Background:
RET/PTC1 is the most prevalent type of gene rearrangement found in papillary thyroid carcinoma (PTC). Previously, we introduced a new noncationic nanosystem for targeted RET/PTC1 silencing by efficient delivery of small interfering RNA (siRNA) using the “squalenoylation” approach. With the aim of improving these results further, we designed new squalenoyl nanostructures consisting of the fusogenic peptide GALA-cholesterol (GALA-Chol) and squalene (SQ) nanoparticles (NPs) of siRNA RET/PTC1.
Methods:
The siRNA RET/PTC1–SQ bioconjugate was synthesized. The corresponding NPs were prepared with or without GALA-Chol by nanoprecipitation and then characterized for their size and zeta potential. The effects of NPs on BHP 10-3 SCmice and TPC-1 cell viability (MTT assay), gene and protein silencing (reverse transcription–quantitative polymerase chain reaction [rt-qPCR], Western blot), and cellular uptake (fluorescent microscopy) were studied. In vivo gene silencing efficiency of siRNA RET/PTC1–SQ NPs was assessed by administration in nude mice via either intratumoral (i.t.) or intravenous (i.v.) routes. Tumor growth was followed for 19 days. Tumors were then collected, and RET/PTC1 gene and protein inhibitions were assessed by RT-qPCR and Western blot.
Results:
The combination of siRNA RET/PTC1–SQ bioconjugate and GALA-Chol leads to stable NPs of ∼200 nm diameter. In vitro, the results revealed that combining GALA-Chol with siRNA RET/PTC1–SQ NPs decreased cell viability, enhanced cellular internalization, and induced gene silencing efficiency in both human PTC (BHP 10-3 SCmice and TPC-1) cell lines. On the contrary, in vivo, the siRNA RET/PTC1–SQ GALA-Chol NPs were not found to be efficient either in gene silencing or in tumor growth inhibition, compared to siRNA RET/PTC1–SQ NPs both via i.t. and i.v. routes (p<0.001).
Conclusions:
Conversely to siRNA RET/PTC1–SQ NPs, the siRNA RET/PTC1–SQ GALA-Chol NPs are efficient in vitro but not in vivo. Finally, NPs of siRNA RET/PTC1–SQ were found to be efficient silencers of the RET/PTC1 fusion oncogene in in vivo applications even at a concentration lower than used in a previously published study.
Introduction
RET/PTC1
Since 2009, our aim has been to introduce a personalized therapy for cancers carrying junction oncogenes, such as PTC, by targeting them with siRNAs. As a first step, an siRNA for RET/PTC1 was designed and found to be efficient in silencing the gene and protein expression in vitro in human BHP 10-3 SCmice and TPC-1 cell lines harboring the RET/PTC1 fusion oncogene (6). Then, to overcome the hurdles of siRNA instability in vivo, we developed an original noncationic nanosystem for efficient in vivo delivery by coupling the siRNA RET/PTC1 to squalene (SQ) (7,8). Squalene, a natural and biocompatible lipid, is a precursor in the biosynthesis of cholesterol in mammalians. It was selected as a transporter moiety because it was found to increase the pharmacological activity of bioactive drugs such as gemcitabine, paclitaxel, and penicillin in several malignancies and resistant intracellular infections (9 –12). Moreover, these bioconjugates have the unique property of self-organizing spontaneously in aqueous solution as nanoparticles (NPs), irrespective of the nucleoside or bioactive molecule used (13,14).
Then, the in vivo efficiency of NPs siRNA RET/PTC1–SQ was tested in the BHP 10-3 SCmice cells, which is the only PTC cell line with a RET/PTC1 rearrangement and tumorigenic properties (15). We found that NPs siRNA RET/PTC1–SQ were efficient in gene and protein inhibition in vivo once injected intravenously in mice bearing BHP 10-3 SCmice tumor xenografts (7). However, these NPs did not induce similar in vitro gene inhibitory effects due to their failure to cross cellular membranes (7). Therefore, to manage the cellular entrance and intracellular trafficking of siRNA, the fusogenic peptide glutamic-alanine-leucine-glutamic acid (GALA) composed of 30 amino acids (16 –18) was specifically combined with siRNA RET/PTC1–SQ NPs to enhance their translocation into the target. GALA has, indeed, extremely low binding affinity to membranes at neutral pH, but undergoes a structural transition from a random coil at pH 7.5 to an amphipathic α-helix at pH 5.0 due to neutralization of the negatively charged glutamic acid residues (19 –21). Thereby it attains a high binding affinity to neutral and negatively charged membranes (22,23). Taking advantage of the mildly acidic condition in the tumor cell environment and in the endosomes/lyosomes compartment, acid-responsive delivery carriers may cause facilitated cellular entry as well as intracellular cytosolic release of oligonucleotides (ONs) from the endosome. This strategy could overcome the intracellular delivery hurdle of ONs and the impossibility for these molecules to escape from the lysosomal trap (24). These properties made GALA an attractive candidate for investigation. Indeed, it has already been used successfully to improve target delivery of siRNA (25 –29) and antisense-ONs (30). The hypothesis was that GALA might interact with the cell membrane in the target tissue and would lead to enhanced transmembrane and intracellular penetration of GALA-combined siRNA–SQ NPs into their destination.
The GALA peptide without attachment to the cholesterol moiety was reported to interact only slightly with lipid membranes, and exhibits a random structure (17,18). Once the cholesteryl moiety was attached, the peptide is probably forced to lie on the membrane to form a helical structure at low pH. Possible loss of repulsion between the negative charge of glutamic acid residues also changes the mode of interaction of the peptide with the membranes due to its structural modification, and eventually facilitates membrane fusion (19). Thus, a cholesterol (Chol) derivative was incorporated in GALA to facilitate its anchoring to siRNA-squalene NPs.
Thus, the aim of this study was to assess if the combination of siRNA RET/PTC1–SQ NPs with a 10% GALA-Chol ratio could enhance the in vitro and in vivo efficiency of siRNA RET/PTC1 in the cell models of human PTC carrying the RET/PTC1 fusion oncogene. This GALA-Chol concentration was previously used in another study and found to be efficient for enhancing luciferase activity in several tissues (31).
We found that siRNA RET/PTC1–SQ GALA-Chol NPs were efficient in vitro but not in vivo. In addition, the present study demonstrates that siRNA RET/PTC1–SQ NPs without GALA-Chol was found to be an efficient silencer in vivo of the RET/PTC1 fusion oncogene, even at a dose lower than that used in our previous study (7).
Materials and Methods
siRNAs synthesis and reagents used
Previously, we described the siRNA RET/PTC1 sequence as well as the irrelevant siRNA control (siRNA CT), a scrambled sequence (6). All single-stranded siRNAs were synthesized by Eurogentec (Belgium) as 21-mer with two 3′ overhanging 2′-deoxynucleotide residues to provide stabilization against nucleases, as described by Tuschl et al. (32). For experiments of cellular uptake, a FAM-labeled antisense strand of siRNA RET/PTC1 was synthesized by Eurogentec. For functionalization, a 3-mercaptopropyl phosphate group was introduced at the 3′-end of the sense strand of each siRNA sequence. GALA-Cholesterol was kindly provided by Dr. Harashima (Hokkaido University, Japan) having the sequence: WEA ALA EAL AEA LAE HLA EAL AEA LEA LAA and is in the form of cholesteryl-GALA (GALA-Chol). Squalene, dimethyl sulfoxide (DMSO), and 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT) reagents were purchased from Sigma-Aldrich Chemical Co. (Saint Quentin Fallavier, France). Dulbecco's modified Eagle's medium (DMEM), Opti-MEM (reduced-serum minimum essential medium), fetal calf serum (FCS), and Lipofectamine™ 2000 (a transfecting reagent) were purchased from Invitrogen (St.-Aubin, France). Water was purified using a Milli-Q system (Millipore, St.-Quentin-en-Yvelines, France). All the chemicals used were of analytical grade.
Synthesis, preparation, and characterization of siRNA RET/PTC1–SQ NPs with and without GALA-Chol
siRNA RET/PTC1 and siRNA control squalene bioconjugates were synthesized, and the corresponding NPs were prepared by nanoprecipitation as previously described (7). Hence, in a typical nanoprecipitation method, NPs of siRNA RET/PTC1–SQ and NPs of siRNA control-SQ were prepared at a molar ratio of 10% GALA-Chol. Briefly, lyophilized siRNA RET/PTC1–SQ bioconjugate (6.12 μM) was synthesized as previously described (7), dissolved in 0.5 mL of ethanol, and then 0.612 μM (10% ratio) of GALA-Chol was added accordingly. The corresponding mixtures were added dropwise under centrifugation at 500 g into 1 mL of 5% aqueous dextrose solution. Subsequently, ethanol was distilled using a rotary evaporator to obtain aqueous suspensions of pure siRNA RET/PTC1–SQ GALA-Chol NPs or siRNA control-SQ GALA-Chol NPs. Double-stranded siRNA were generated by adding equimolar amounts of both forward and reverse strands (labeled by FAM or not) in annealing buffer (30 mM HEPES-KOH (pH 7.4), 2 mM Mg acetate, 100 mM K acetate) for 2 min at 95°C and then kept for 1 h at room temperature. The duplex formation was assessed by 4% agarose gel electrophoresis. The hydrodynamic diameter (nm) of the NPs was measured at 20°C by quasi-elastic light scattering using a Nanosizer 4 (Malvern Instruments SA, Orsay, France) operating at 90°. Zeta potential (mV) was measured using a Zetasizer 4 (Malvern Instruments).
Stability of NPs
In order to assess the stability of NPs, the suspension of siRNA RET/PTC1–SQ GALA-Chol NPs was stored at room temperature. Size and zeta potential were measured at different time points—at 0, 15, 30, 90, and 180 days.
Determination of nanoassembly structure by transmission electron microscopy
Transmission electron microscopy (TEM) was performed using a Philips EM208 electron microscope with a large-format CCD camera AMT (FEI Co., Eindhoven, The Netherlands) at the Joint Center for Electron Microscopy (CCME; Orsay, France). The samples were deposited on a pre-ionized Formvar carbon-coated electron microscope grid (FEI). The excess sample was removed after 5 min and stained with 1% neutral aqueous phosphotungstic acid as a contrasting agent. After 30 s, samples were observed under a microscope.
Cell culture
Both human TPC-1 (gift from Dr. C. Dupuy, Institut Gustave Roussy, Villejuif, France) and BHP 10-3 SCmice (kindly provided by Dr. G. Clayman, MD Anderson Cancer Center, Houston, TX) cell lines harbor the RET/PTC1 fusion oncogene. TPC-1 cells did not colonize in soft agar and were found to be nontumorigenic when injected in nude mice, while BHP 10-3 SCmice cells, derived from the BHP 10-3 cell line, colonized in soft agar and produced tumors in 100% of injected mice (15). Both cells lines were maintained at 37°C in an atmosphere of 5% CO2 and 95% humidity in DMEM supplemented with 10% FCS, penicillin (100 U/mL), and streptomycin (10 μg/mL). Before further experiments, polymerase chain reaction (PCR) analysis was performed to demonstrate that the cells were free of mycoplasma.
Viability assay
Cell viability was evaluated using the MTT (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide) assay, as previously reported (33). Briefly, BHP 10-3 SCmice and TPC-1 cells were seeded into 96-well culture plates at 3×103 cells/well. After 24 h, the culture medium was removed and replaced with 100 μL of fresh medium containing siRNA RET/PTC1–SQ GALA-Chol (10%) NPs at 50 nM final concentration either alone or in the presence of the transfecting agent (Lipofectamine 2000). After 24 h, 48 h, and 72 h post-transfection incubation at 37°C, MTT reagent (5 mg/mL in PBS) was added to the culture medium (Sigma-Aldrich) followed by 2 h incubation at 37°C. The cells were lysed by sodium dodecyl sulfate (SDS) solution (SDS 10%, 10 mM HCl) overnight at 37°C. The optical density was measured at 570 nm wavelength using a microplate reader (MRXII photometer; Dynex Technologies, Thermo Scientific, Courtaboeuf, France). All experiments were performed in triplicate. The results are expressed as a percentage after normalization over untreated cells.
Determination of nanoassembly gene silencing effect by reverse transcription quantitative PCR
BHP 10-3 SCmice or TPC-1 cells were seeded in 6-well plates at 4×105 cells/well density. After 24 h, cells were (i) transfected with 50 nM of siRNA RET/PTC1 or siRNA CT by using Lipofectamine 2000, as described by Gilbert-Sirieix et al. (6); and (ii) treated with siRNA CT-SQ NPs, siRNA RET/PTC1–SQ NPs, or siRNA RET/PTC1–SQ GALA-Chol NPs (10% ratio) either alone or in the presence of the transfecting agent. Cells were incubated for 24 h or 48 h before the analysis of RET/PTC1 gene knockdown.
For the determination of RET/PTC1 gene expression, the total RNA of cells was extracted using an RNeasy mini-kit (Qiagen Ltd., Crawley, United Kingdom). RNA quantity and purity was assessed using NanoDrop ND-1000 spectrophotometer (Wilmington, NC). M-MLV Reverse Transcriptase buffer pack (Promega, Lyon, France) was used for reverse transcription as previously described (7). Primers were designed based on the human RET/PTC1 sequence (H4/RET mRNA; GenBank: AF395885.1); oligonucleotide forward: 5′-AGA TAG AGC TGG AGA CCT AC-3′ and reverse: 5′-CTG CTT CAG GAC GTT GAA-3′, to produce PCR fragments of 290 bp. Primer sequences for ribosomal protein L13A (RPL13A, used as an internal gene control) were: 5′-CAT GAG GCT ACG GAA ACA GG-3′ (forward) and 5′-TCG GGA GGC AGT GAC TAA GAC-3′ (reverse). The amplification was monitored on a StepOnePlus PCR System (AB Applied Biosystems, Perkin-Elmer, Courtaboeuf, France) using SYBR Green Reagent (Invitrogen) according to the manufacturers' instructions. Experiments were performed in triplicate, and samples were run in triplicate with primer sets of the RET/PTC1 and the RPL13A genes. Gene regulation was determined by the quantification-comparative ΔΔCT method (34), normalized to RPL13A expression and compared to nontreated cells (35).
Determination of nanoassembly protein silencing effect by Western blot analysis
For the determination of RET/PTC1 protein expression, total protein was extracted from the cells using Mammalian protein extraction reagent (M-PER®; Thermo Scientific, Rockford, IL) supplemented with complete mini EDTA-free protease inhibitor cocktail (Roche Diagnostics, Meylan, France). Proteins were titrated by the BCA method using Micro BCA protein assay kit (Thermo Scientific). An amount of 30 μg of cell protein extract was heated at 70°C for 10 min with 1× sample reducing buffer (NuPAGE® sample reducing agent; Invitrogen) and 1X sample loading buffer (NuPAGE® LDS sample agent; Invitrogen) in a mixture of M-PER and protease inhibitor. The samples were then run on polyacrylamide gels (NuPAGE® Bis-Tris Mini Gels 10%; Life Technology, St.-Aubin, France), and the proteins were transferred to nitrocellulose membrane using the iBlot dry blotting system (Invitrogen). The membranes were saturated with 10% nonfat dry milk (Bio-Rad, Marnes-la-Coquette, France) in PBS with 0.1% Tween-20, and then were incubated overnight at 4°C under agitation with the primary antibodies specific for RET (rabbit Ret EPR2871; 1:1000, abcam Biochemicals®) or for GAPDH (mouse Abcys®, 1:1000; Sigma-Aldrich; used as an internal control) in 10% nonfat dry milk solution. Blots were then washed and incubated with anti-rabbit or anti-mouse antibodies conjugated with horseradish peroxidase. Bands were visualized by enhanced chemiluminescence (Invitrogen). For quantification of relative protein inhibition, the films were scanned with Gel Doc™ XR+ Systems (Bio-Rad) using a calorimetric method. Images were then analyzed using Bio-Rad Image Lab software, and the RET protein was quantified after being normalized to GAPDH. The results are expressed as relative protein expression compared to nontreated cells.
Cellular uptake of NPs
The BHP 10-3 SCmice cells (1.5×105 cells/well) were seeded in 12-well plates containing a cover glass one day prior to transfection. Cells were transfected with 50 nM of FAM-labeled (i) siRNA RET/PTC1 alone or (ii) with Lipofectamine 2000, or treated with (iii) siRNA RET/PTC1–SQ NPs or (iv) siRNA RET/PTC1–SQ GALA-Chol NPs. After 4 h of incubation at 37°C, the medium was removed, and the cells were washed twice with PBS and then fixed with formaldehyde (4% in PBS) for 20 min at room temperature. After final rinses in 0.1% PBS-Tween and PBS, slides were stained with Vectashield containing 4′,6-diamidino-2-phenylindole (DAPI), and fluorescence was assessed by fluorescence microscopy (Zeiss LSM 510/Axiovert 200 M, Carl Zeiss SAS, Marly le Roi, France).
Animal experimentation
All animal experiments were carried out according to the French laws of animal welfare and were approved by the Ethics Commission of the official veterinary authorities. Six-week-old female nude nu/nu mice were purchased from Harlan Laboratory and housed in a sterilized laminar flow caging system, and food, water, and bedding were autoclaved before putting in cages. BHP 10-3 SCmice cells (2×106) were suspended in 200 μL PBS and injected subcutaneously into the mice toward the upper portion of the right flank.
In vivo effects of nonvectorized siRNAs
Mice (n=3 per group) were treated either with nonvectorized siRNA RET/PTC1 or with siRNA CT when the tumor volume reached 50 mm3 (day 0). Five injections of each treatment (siRNA RET/PTC1 or siRNA CT) were given at three different doses, that is, 25 μg/kg, 100 μg/kg, or 500 μg/kg (cumulative doses=0.125 mg, 0.5 mg, or 2.5 mg/kg respectively) on days 0, 2, 4, 7, and 10 intravenously in the lateral tail vein, and tumors were collected at the end of the experiment (day 20).
In vivo efficiency of NPs siRNA RET/PTC1–SQ with or without GALA-Chol
When the tumor volume reached about 50 mm3, mice (n=6 per group) were treated intravenously either with normal saline (NaCl 0.9%), siRNA CT-SQ NPs, siRNA RET/PTC1–SQ NPs, or siRNA RET/PTC1–SQ GALA-Chol NPs dispersed in 100 μL of 0.9% NaCl solution at the rate of 0.1 mg/kg/injection (cumulative dose=0.5 mg/mouse). Similarly, for the i.t. route, mice (n=6 per group) were treated with the same four conditions with an additional condition of empty 1,1′,2-tris-nor-squalenic acid NPs (SQ NPs) at the rate of 0.1 mg/kg/injection (cumulative dose=0.5 mg/mouse). The injections were given on days 0, 2, 4, 7, and 10, and the tumor growth was followed starting from day 0 to the end of the experiment (day 19). Mice were sacrificed at the end of the experiment on day 19 (n=6), when the mice treated with normal saline reached the ethical tumor size limit.
Tumor length and width were measured with calipers, and the volume was thus calculated using the equation: Tumor volume (V)=length (l)×width2/2 (36). Mice were monitored daily for changes in tumor size and body weight. At the end of the experiments, mice were sacrificed; and tumors were collected and immediately frozen in liquid nitrogen for reverse transcription quantitative PCR (RT-qPCR) analysis and Western blotting.
RNA extraction and RT-qPCR from tumors
For the determination of the RET/PTC1 expression, tumors were grinded and mixed well to avoid any heterogeneity in tumor composition. Total RNAs were extracted from specified tumor quantities using an RNeasy mini-kit according to the manufacturer's instructions. RNA quantity and quality control, reverse transcription, and RT-qPCR were performed as explained above. The results are presented as relative mRNA expression compared to the 0.9% saline-treated group.
Protein extraction and Western blotting from tumors
Protein extraction and Western blot from ground tumors were performed as described in the section Determination of nanoassembly protein silencing effect by Western blot analysis. The RET protein was quantified after being normalized with GAPDH, and the results are expressed as relative protein expression compared to tumors treated with 0.9% NaCl.
Statistical analysis
Data are presented as mean±standard deviation (SD). Comparisons between multiple treatments were made using one-way analysis of variance (ANOVA) followed by Bonferroni's or Newman–Keuls multiple comparison test. All pairwise comparisons between tumor growths of different treatment groups at different days were done by least significant difference (LSD) post-hoc test using InVivoStat software (Cambridge, United Kingdom). The level of significance was set at p<0.05.
Results
Characterization of siRNA RET/PTC1–SQ GALA-Chol NPs
The attachment of the squalene was performed at the 3′-end of the 21-mer sense strand siRNA RET/PTC1 to protect it against 3′-exonucleases. This modification prevents steric hindrance toward recognition of the targeted mRNA by the antisense strand, and allows fixation of proteic RISC complex at the 5′-end required for siRNA phosphorylation (7). The bioconjugate induced the spontaneous formation of NPs (siRNA CT-SQ, siRNA RET/PTC1–SQ, and siRNA RET/PTC1–SQ GALA-Chol) in water as measured by laser light scattering. The NPs were found to be negatively charged, which favors establishing repulsive forces allowing the stabilization of the NPs suspension. The shape of siRNA RET/PTC1–SQ GALA-Chol NPs was found to be spherical by TEM (Fig. 1A). The addition of the GALA-Chol during the nanoprecipitation led to a slight increase in the NP diameter (∼190 nm for siRNA RET/PTC1–SQ GALA-Chol NPs vs. 160 nm for siRNA RET/PTC1–SQ NPs; Fig. 1A, B). Both size and zeta potential of siRNA RET/PTC1–SQ GALA-Chol NPs remained nearly constant during six months' incubation at room temperature (Fig. 1B).
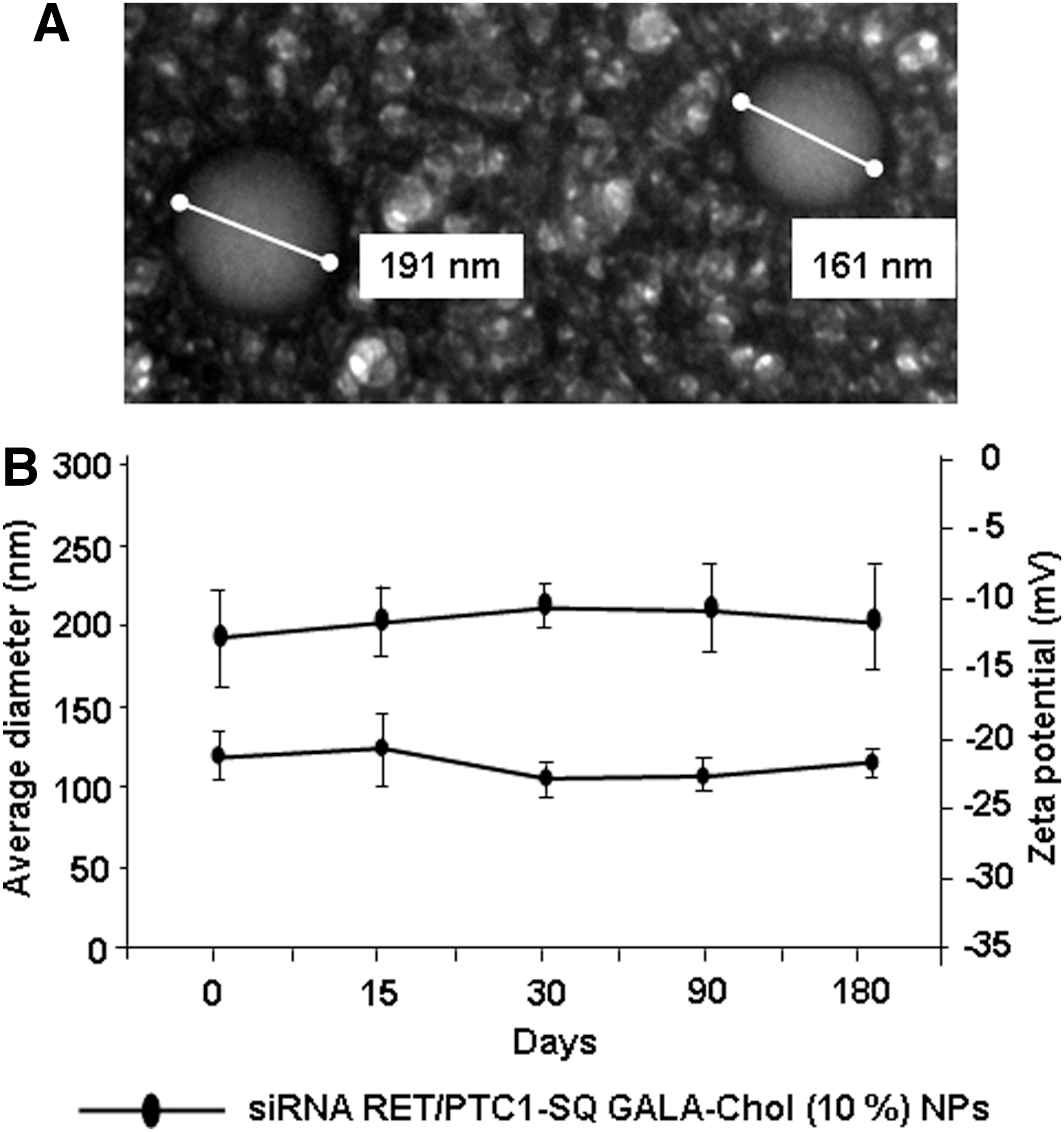
Characterization and stability of siRNA RET/PTC1–SQ GALA-Chol nanoparticles (NPs).
Effects of GALA-Chol combined with siRNA RET/PTC1–SQ NPs on cell viability
Effects of siRNA RET/PTC1–SQ GALA-Chol NPs on cell viability were assessed by MTT assay on two different cell lines both expressing the RET/PTC1 fusion oncogene (BHP 10-3 SCmice and TPC-1) at 24, 48, and 72 h at 50 nM siRNA RET/PTC1 concentration and 10% GALA-Chol ratio. An inhibition (p<0.01) in growth rate (∼10% at 24 h and 20% at 48 h and 72 h) of both cell lines treated with siRNA RET/PTC1–SQ GALA-Chol NPs was observed compared to nontreated cells, which increased significantly (p<0.001) with the use of Lipofectamine 2000 (±50% inhibition; Fig. 2A and B).
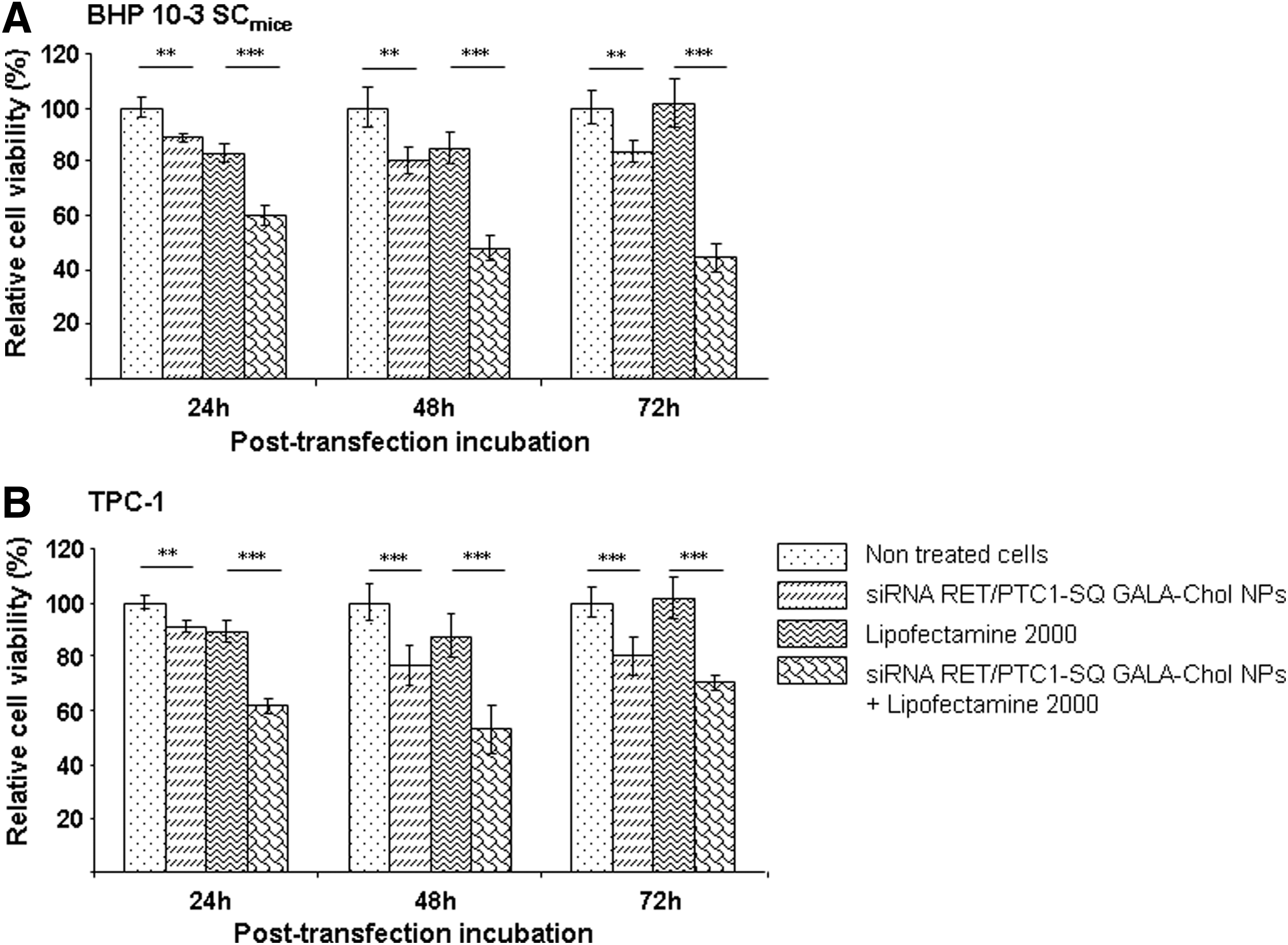
Effects of siRNA RET/PTC1–SQ GALA-Chol NPs on cell viability. Effects of siRNA RET/PTC1–SQ GALA-Chol NPs (10%) at 50 nM siRNA concentration were tested on cell viability by MTT assay at 24, 48, and 72 h with and without Lipofectamine 2000 in both BHP 10-3 SCmice
Effects of GALA-Chol combined with siRNA RET/PTC1–SQ NPs on RET/PTC1 gene and protein expressions
We tested the effects of GALA-Chol combined with siRNA RET/PTC1–SQ NPs on RET/PTC1 gene (RT-qPCR, n=3) and protein (Western blot, n=3) silencing. As shown in Figure 3, siRNA RET/PTC1–SQ GALA-Chol NPs significantly inhibited RET/PTC1 gene (Fig. 3A-i, B-i) and protein (Fig. 3A-ii, B-ii) expression in both cell lines at 24 h and 48 h.
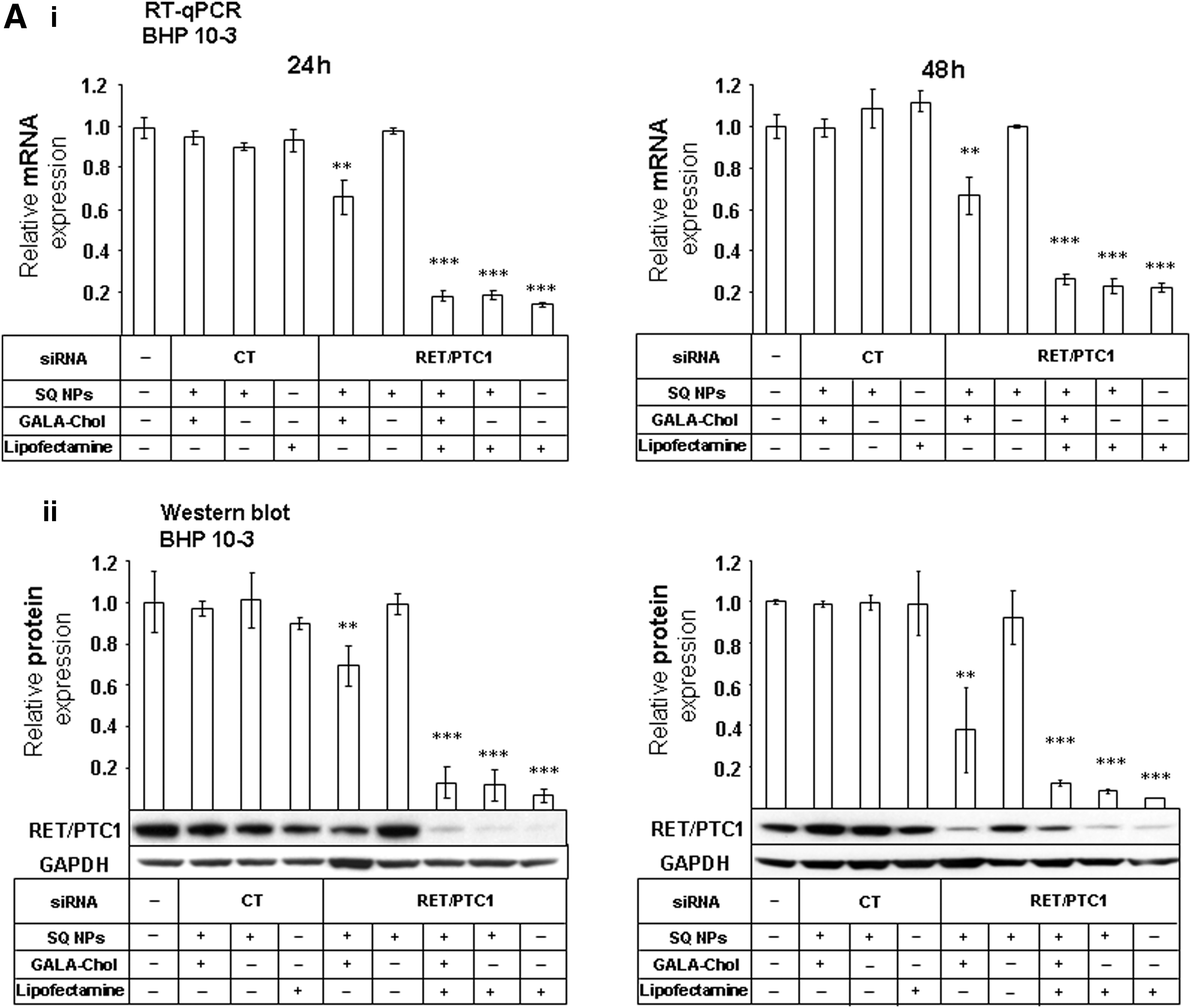
Gene and protein silencing efficiency of NPs in BHP 10-3 SCmice and TPC-1 cell lines. To assess the gene silencing efficiency, siRNA RET/PTC1–SQ NPs and siRNA CT-SQ NPs with and without GALA-Chol (10%) were added to BHP 10-3 SCmice
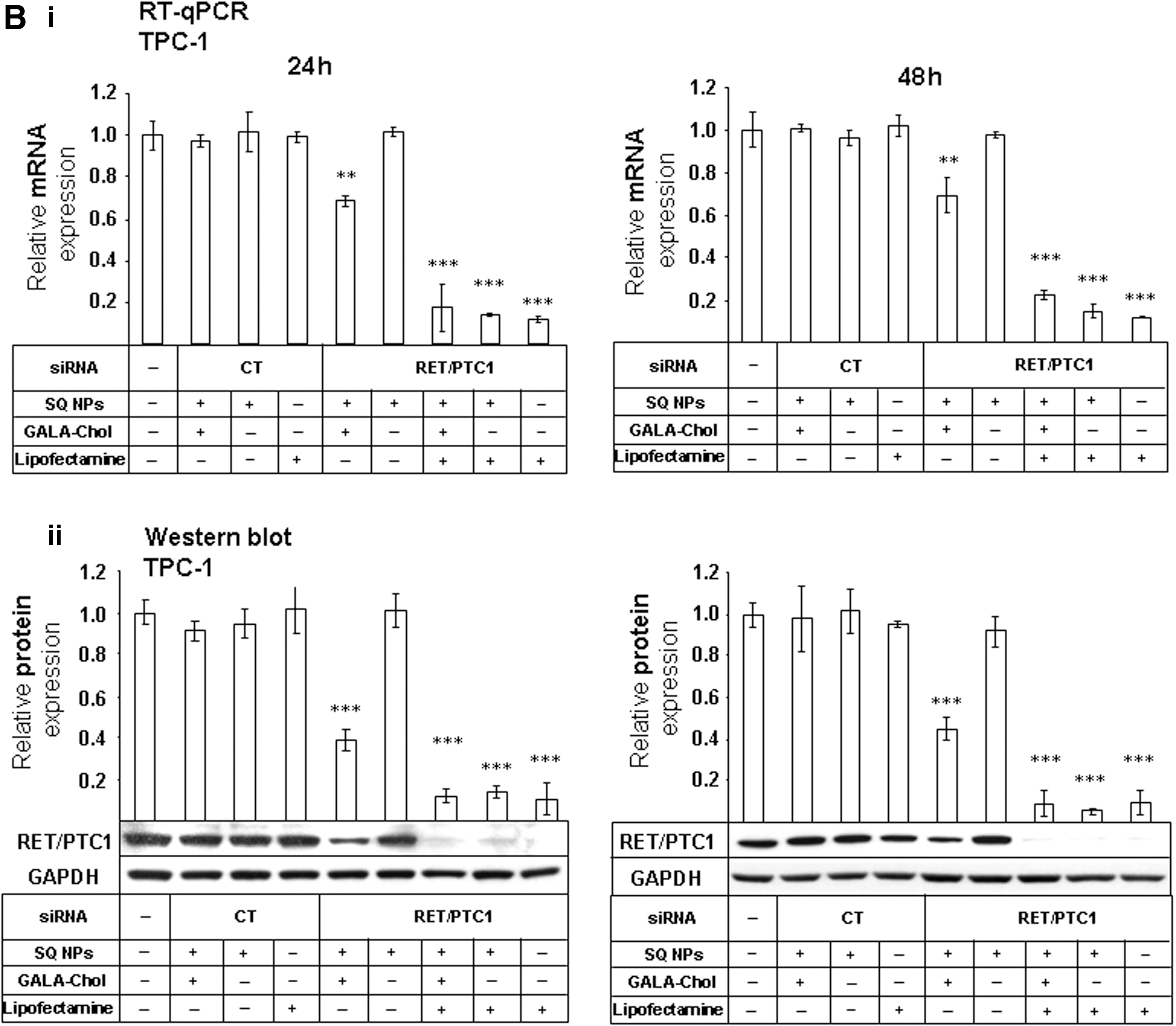
On the contrary, siRNA RET/PTC1–SQ NPs alone were unable to inhibit gene and protein expression (Fig. 3). Similar to the results obtained for the nonvectorized siRNA RET/PTC1 transfected with Lipofectamine 2000, siRNA RET/PTC1–SQ NPs transfected with the same agent showed a significant inhibition of RET/PTC1 mRNA and protein expression (Fig. 3).
Intracellular uptake of siRNA RET/PTC1–SQ NPs with and without GALA-Chol in BHP 10-3 SCmice cells
The visualization of NP internalization in BHP 10-3 SCmice cells was realized by fluorescent microscopy. The fluorescence emitted from the FAM-labeled NPs was localized in intracellular compartments, indicating that the siRNA RET/PTC1–SQ GALA-Chol NPs were efficiently internalized into the cells (Fig. 4A). As expected, nonvectorized siRNA RET/PTC1, once transfected with Lipofectamine 2000, was also internalized (Fig. 4B), but neither siRNA RET/PTC1–SQ NPs (Fig. 4C) nor the untransfected nonvectorized siRNA RET/PTC1 (Fig. 4D) were able to cross the membrane barrier in the BHP 10-3 SCmice cells.
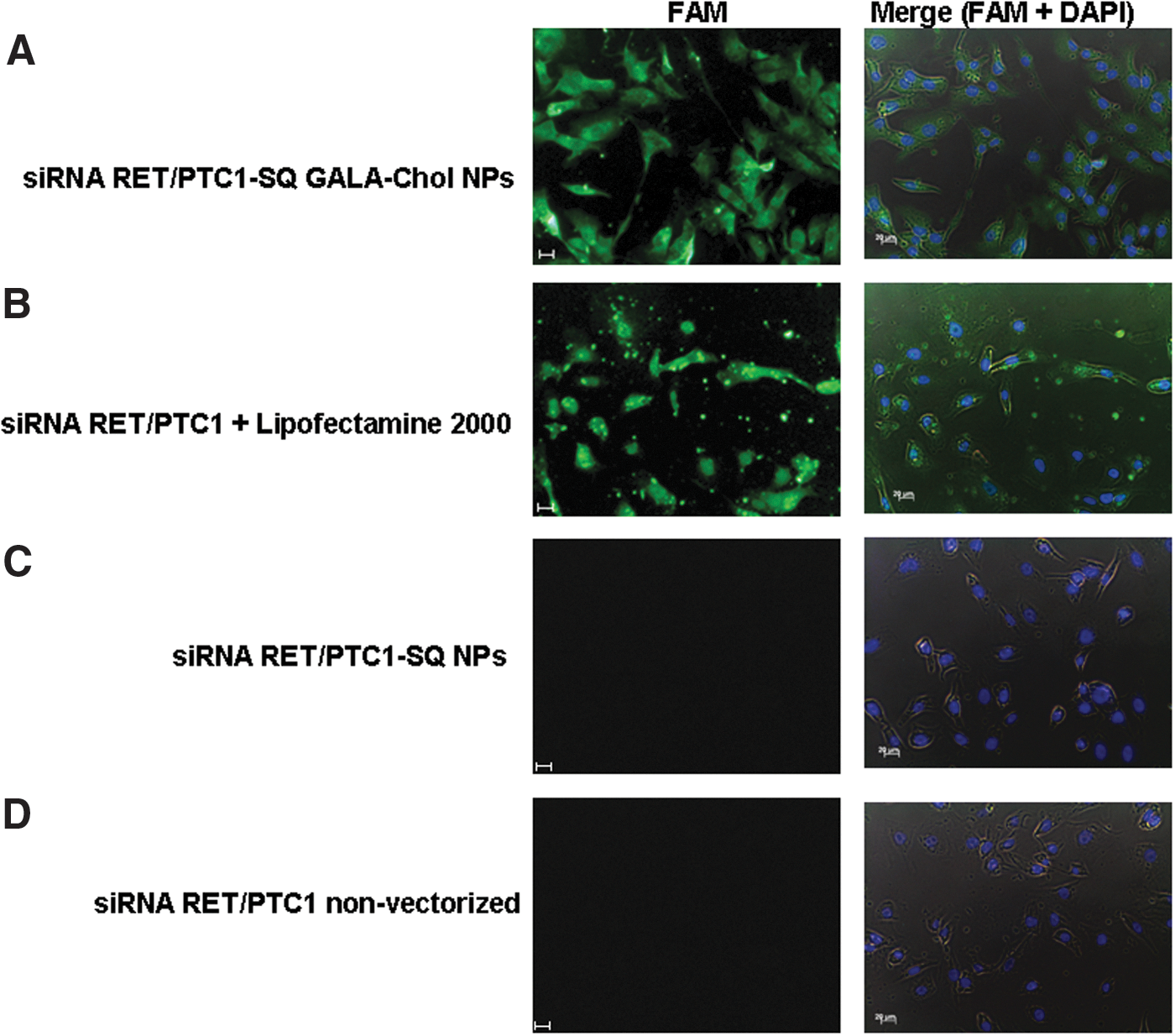
Uptake of NPs in BHP 10-3 SCmice cell line. The intracellular trafficking of FAM-labeled NPs was assessed by using fluorescence microscope. 1.5×105 cells/well were incubated at 37°C for 24 h prior to transfection and then left for 4 h either with 50 nM of FAM-labeled siRNA RET/PTC1–SQ GALA-Chol NPs
Effect of nonvectorized siRNAs on mice bearing BHP 10-3 SCmice xenografts
TPC-1 cells were found to be nontumorigenic in mice. Thus, only the BHP 10-3 SCmice cell line was used to develop a solid tumor model. In order to establish the dose of siRNA RET/PTC1 able to provide effective gene inhibition without side effects, we first followed the tumor growth after injection of different doses of nonvectorized siRNA CT or target siRNA RET/PTC1. The cumulative dose of 2.5 mg/kg (i.e., 500 μg/kg in each injection) of either control or target siRNA indistinctly inhibited tumor growth (p<0.001). However, both siRNAs did not influence gene or protein levels (Supplementary Fig. S1; Supplementary Data are available online at
Efficacy of siRNA RET/PTC1–SQ NPs alone or combined with GALA-Chol via the i.t. route
We tested the efficiency of siRNA NPs after i.t. injection in nude mice (n=6). Squalenoylated siRNA RET/PTC1 NPs as well as squalenoylated siRNA CT NPs with and without GALA-Chol (10%) were injected at a cumulative dose of 0.5 mg/kg. Moreover, in this experiment, 1,1′,2-tris-nor-squalenic acid NPs (SQ NPs) were injected as a control to assess if this terpenoid could also affect tumor growth. Noteworthy, no weight loss or overall toxicity in all treated groups were observed. Interestingly, after i.t. injection, a significant inhibition of tumor growth was observed when tumors were treated with siRNA RET/PTC1–SQ NPs (LSD test, p<0.001). In contrast, siRNA RET/PTC1–SQ GALA-Chol NPs did not significantly affect tumor growth (Fig. 5A).
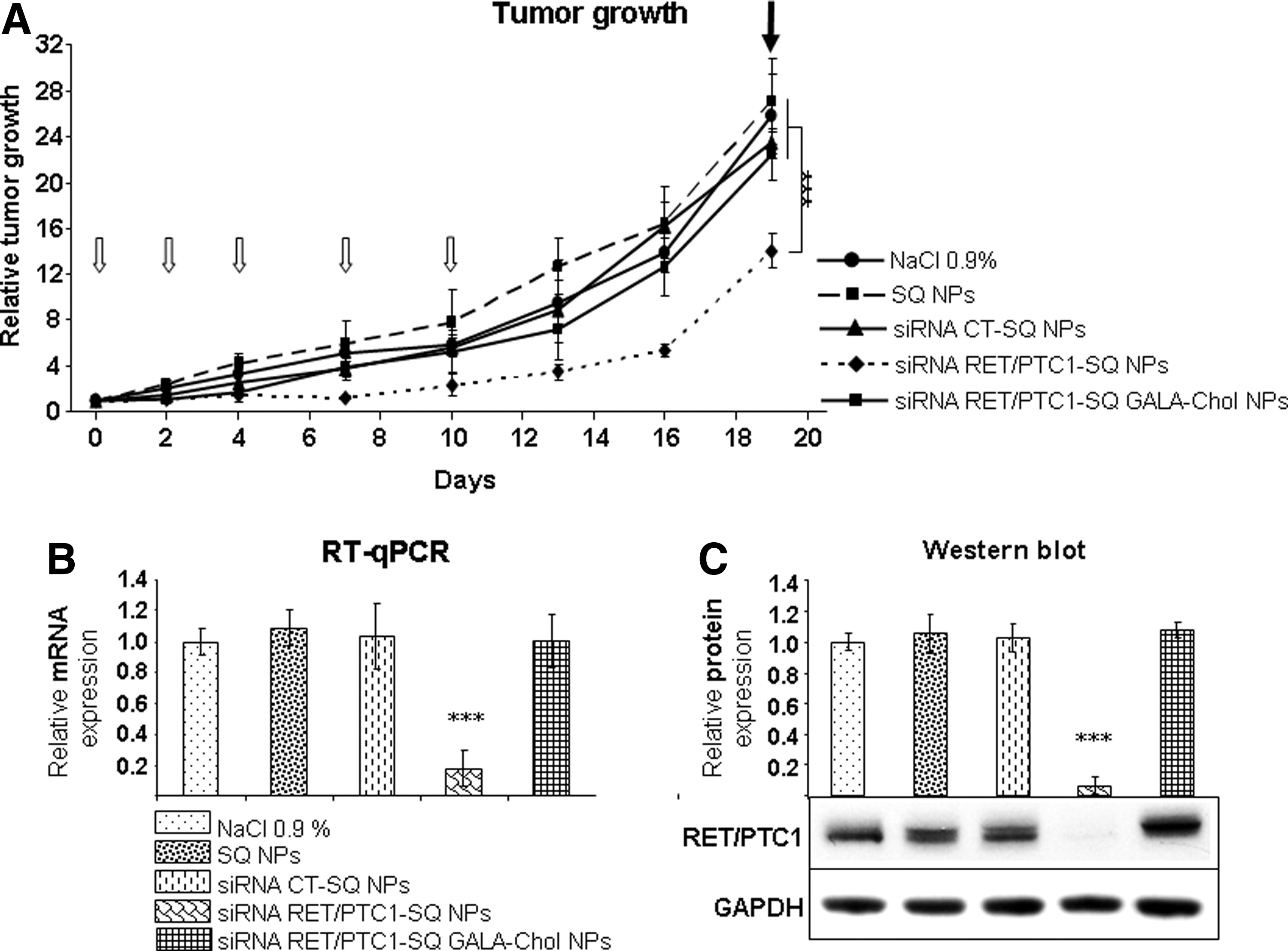
Antitumor effects of siRNA RET/PTC1–SQ NPs with or without GALA-Chol through the intratumoral (i.t.) route.
Concerning the RET/PTC1 gene expression, tumors treated with siRNA RET/PTC1–SQ NPs showed a significant inhibition of ∼80% at the end of the experiment (Fig. 5B). In addition, a complete inhibition of protein expression was observed by Western blot (Fig. 5C).
Efficacy of siRNA RET/PTC1–SQ NPs alone or combined with GALA-Chol via the i.v. route
Because not all tumors can be treated by local administration, we decided to assess the ability of siRNA NP formulations to inhibit tumor growth after i.v. injection. As described in Materials and Methods, squalenoylated siRNA RET/PTC1 NPs (with and without GALA-Chol, 10%) and siRNA CT NPs were injected in nude mice (six per group) intravenously at 0.5 mg/kg cumulative dose. By using the LSD test and as observed in Fig. 6A, a highly significant (p<0.001) inhibition of tumor growth was observed only in tumors treated with siRNA RET/PTC1–SQ NPs. The growth curve of siRNA RET/PTC1–SQ GALA-Chol NPs followed that of siRNA CT-SQ NPs and was not very different from the group receiving saline solution (0.9% NaCl).
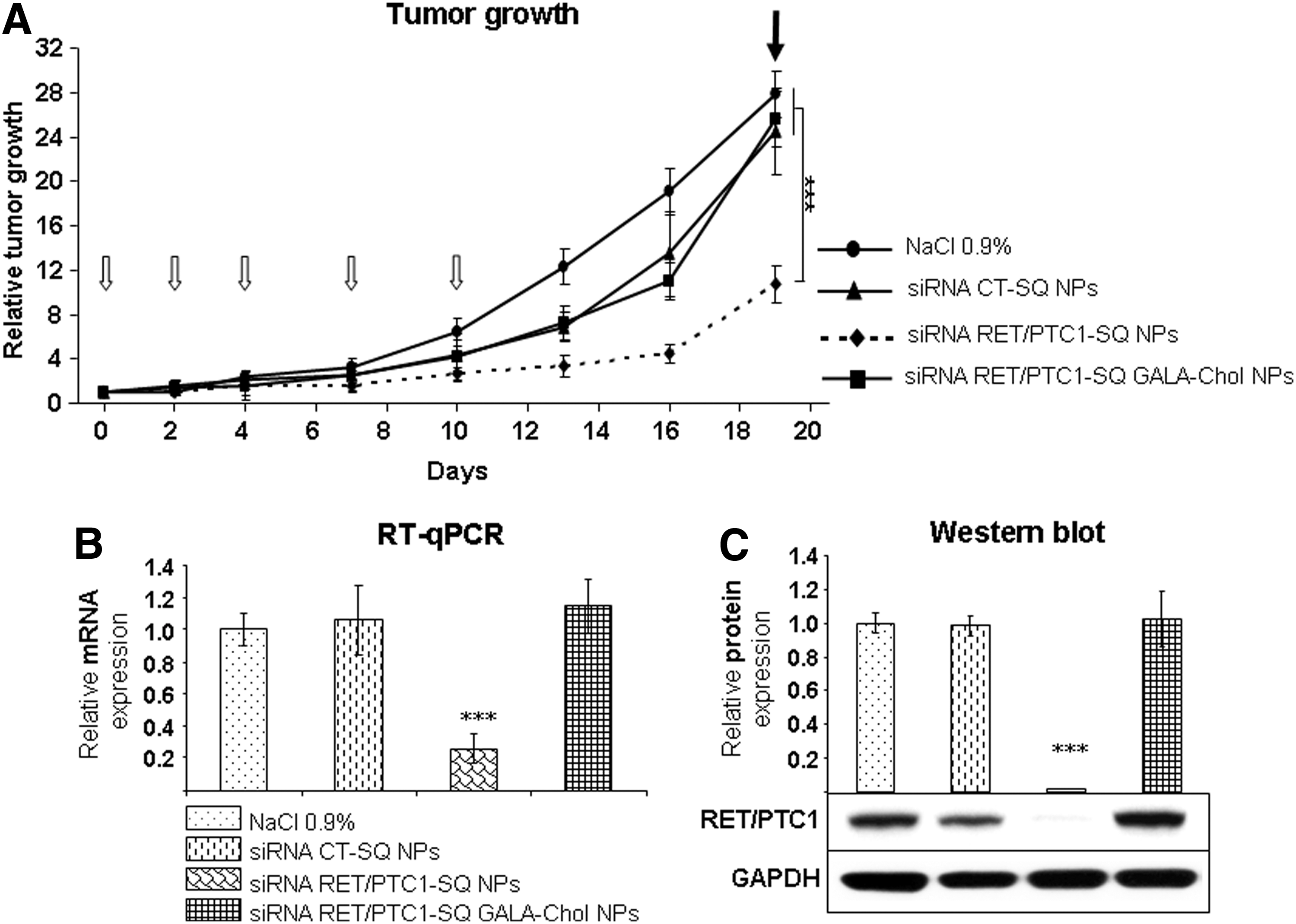
Antitumor effects of siRNA RET/PTC1–SQ NPs with or without GALA-Chol through the intravenous (i.v.) route.
Tumors treated with siRNA RET/PTC1–SQ NPs showed a highly significant inhibition (∼70%) of RET/PTC1 mRNA expression compared to tumors treated with other conditions (p<0.001; Fig. 6B). Similarly, a decreased protein expression was observed by Western blot in the tumors treated with siRNA RET/PTC1–SQ NPs compared to other groups (p<0.001; Fig. 6C).
Discussion
In a previous study, we have applied the “squalenoylation” concept for the administration and transport of siRNA against the RET/PTC1 fusion oncogene and found that the siRNA-squalene bioconjugate formed NPs of about 170 nm. These nanoassemblies were able to inhibit tumor growth and RET/PTC1 expression in vivo but were found inefficient in vitro (33). In this study, we hypothesized that because GALA-Chol might interact with the cell membranes in the target tissue, it would improve the transmembrane and intracellular penetration of siRNA-SQ NPs into their destination.
Thus, GALA-Chol was added at a 10% ratio to siRNA RET/PTC1 squalene and submitted to nanoprecipitation conditions. The siRNA RET/PTC1–SQ GALA-Chol bioconjugate was able to form spherical NPs with a slightly larger size than those synthesized with siRNA RET/PTC1–SQ alone. The zeta potential remained negative, which favors establishing repulsive forces allowing the stabilization of the NP suspension, as observed for six months in this study.
In vitro biological assessment showed that siRNA RET/PTC1–SQ GALA-Chol nanoassemblies displayed effects on cell viability in two different cell lines (BHP 10-3 SCmice and TPC-1, both expressing the RET/PTC1 fusion oncogene) along with efficient inhibition of RET/PTC1 mRNA and protein expression. In contrast, siRNA RET/PTC1–SQ NPs without GALA-Chol did not silence mRNA and protein expression. The in vitro inefficiency of siRNA RET/PTC1–SQ NPs can be explained by our experiments on cellular uptake of NPs. We believe that GALA-Chol plays a role not only in endosomal escape and cytoplasmic delivery but also in intracellular penetration of NPs. Indeed, transmembrane penetration of siRNA RET/PTC1–SQ NPs is probably improved by the GALA positive charge, whereas siRNA/PTC1–SQ NPs alone were unable to be internalized unless transfected with positively charged agents (Lipofectamine 2000). Moreover, a GALA conformational change, due to the slight acidic pH of the tumor cell environment, might help cellular penetration. The inefficiency of the siRNA RET/PTC1–SQ NPs in gene silencing might not be due to siRNA degradation, but rather result from the inability of NPs to enter the cells.
As siRNA RET/PTC1–SQ GALA-Chol NPs were found to be efficient in vitro, and siRNA RET/PTC1–SQ NPs, as previously described, were able to reduce tumor growth in vivo (33), we tested the antitumor activity of the former in mice bearing BHP 10-3 SCmice xenografts. However, as described by several authors (36 –38), high doses of siRNA can induce side effects. In order to avoid nonspecific effects and also to prevent toxicity, we first determined the suitable dose of nonvectorized siRNAs (RET/PTC1 and CT). We found that the highest concentration (2.5 mg/kg) significantly inhibited tumor growth no matter which siRNA was administrated. On the contrary, gene expression analysis did not show any RET/PTC1 inhibition, which is clear evidence of side effects, rather than a specific ability of gene inhibition. Therefore, for further experiments, the cumulative dose of 0.5 mg/kg was selected to ascertain the therapeutic effects of siRNA RET/PTC1.
Nanoassemblies of siRNA RET/PTC1–SQ NPs with or without GALA-Chol were administrated via intratumoral or intravenous injections. Surprisingly, only siRNA RET/PTC1–SQ NPs induced a regression of tumor growth and inhibition of mRNA and protein RET/PTC1 expression after both routes of administration (i.t. and i.v.). A possible explanation is that a 10% GALA-Chol concentration might cause aggregates in serum containing large quantities of nonspecific proteins in vivo compared to the serum-deficient conditions used for in vitro experiments. In fact, Sakurai et al. (28) showed that GALA-Chol concentration may affect tumor siRNA delivery, and Walkey et al. (39) demonstrated that variations in serum protein adsorption correlate with differences in the mechanism and efficiency of NP uptake by macrophages. Indeed, Khalil et al. (31) proved that GALA accumulates preferentially in the liver, thus reducing the distribution of the therapeutic siRNA in tumors.
These data demonstrate once again that prediction of in vivo pharmacological efficacy through cell culture experiments is uncertain, even when experiments are performed in exactly the same tumor cell line. We have already observed such a phenomenon with siRNA loaded into chitosan-coated nanospheres (40). The possible reasons are numerous: (i) cells in culture may exhibit different pattern of gene expression and differentiation than in vivo; (ii) the physicochemical properties of the NPs may change in vivo: aggregation, dissociation, or reorganization due to physiological salts, opsonins, or lipids, among others; (iii) the cell culture experiments that are usually performed for 24 hours differ from in vivo experiments that last for weeks; and (iv) enzymatic contents are entirely different in in vitro compared to in vivo conditions. This should significantly influence the kinetic release of the oligonucleotide from the squalenoylated NPs.
The observed results show that GALA-Chol abolished SQ NPs efficiency, excluding its use for further investigations. In contrast, siRNA RET/PTC1–SQ NPs could have an interesting medical application, since their effects on gene silencing are persistent. They are able to maintain gene knockdown up to nine days after the end of treatment, which means that the system is highly stable and able to maintain the effects of siRNA, which are usually transitory.
For further studies, we suggest performing a longer treatment schedule and combining siRNA RET/PTC1–SQ NPs with 131I radioiodine therapy, which is the most commonly used treatment for PTC.
In conclusion, we have demonstrated that the use of GALA-Chol improves gene and protein silencing efficiency of siRNA RET/PTC1–SQ NPs in vitro but not in vivo. On the other hand, siRNA RET/PTC1–SQ NPs showed a significant inhibition of the target gene in vivo, using a fivefold lower dose than previously published. Finally, we suggest that siRNA RET/PTC1–SQ NPs can be an efficient therapy for RET/PTC1 fusion oncogene inhibition, and this observation opens the way for further pharmacological studies intended for in vivo applications.
Footnotes
Acknowledgments
The research leading to these results has received funding from the Agence Nationale de Recherche ANR, Programme P2N, Grant No. NANO 00301 and from the European Research Council, Programme FP7/2007-2013 Grant No. 249835.
Author Disclosure Statement
The authors declare no conflicts of interest.