
Abstract
Background:
The innate immune system is the first line of defense and plays a key role in thyroid cancer development. The role of the tumor-infiltrating natural killer (NK) cells is becoming increasingly important in research and potential cancer therapies. NK cell subpopulations, CD3−CD16+CD56dim and CD3−CD16−CD56bright, demonstrate a significant role in the tumor immuno-surveillance process.
Methods:
We investigated the distribution of CD3−CD16+CD56dim and CD3−CD16−CD56bright NK subpopulations in tissue and blood samples from patients with papillary thyroid cancer (PTC) and nodular goiter (NG). Twenty-eight patients with PTC, 13 patients with NG, and 50 healthy donors were included in the study. Tissue and blood samples from all patients and blood samples from healthy donors were analyzed for CD3−CD16+CD56dim and CD3−CD16−CD56bright NK cells by flow cytometry.
Results:
A significant predominance of CD3−CD16+CD56dim cells compared to CD3−CD16−CD56bright NK cells was found in blood samples in all groups (p<0.0001 in PTC, NG, and healthy donors). Increased infiltration by CD3−CD16−CD56bright NK cells was observed in thyroid tissue of patients with PTC, as compared to CD3−CD16+CD56dim NK cells (p=0.046), while CD3−CD16+CD56dim NK cells demonstrated a higher infiltration of NG tissues. CD3−CD16+CD56dim NK cell tissue infiltration positively correlated with advanced stages of PTC. In contrast, the CD3−CD16−CD56bright NK cell population was negatively associated with tumor stage in patients with PTC.
Conclusion:
CD3−CD16−CD56bright NK cell infiltration seems to be associated with PTC progression. These findings contribute to a better understanding of the immune response in PTC and may lead to novel immunotherapeutic approaches in these patients.
Introduction
I
Despite the fact that CD16+CD56dim cytotoxic NK cells are increased in peripheral blood, CD16−CD56bright immunoregulatory NK cells prevail in tissues such as tonsils, lymph nodes, spleen, and placenta (5,7 –9). CD16−CD56bright NK cells mainly produce cytokines, such as IFN-γ, TNF-α), GM-CSF, interleukin (IL)-10, and IL-13 following monokine stimulation (5,10). Although it is believed that only cytotoxic and not immunoregulatory NK cells produce proteases, such as the granzymes and the protein perforin, which cause apoptosis and lysis of target cells (11), a previous report indicated that freshly isolated CD56bright and CD56dim NK cells show similar levels of expression of perforin (12). Furthermore, although it was thought until recently that CD56dim NK cells could not produce cytokines, novel data suggest that they can also produce high quantities of cytokines after engagement of the activating receptors or after short-term cytokine stimulation (13,14). Connection of NK cells with cells that do not express MHC-I, such as tumor cells, makes the targets susceptible to NK mediated lysis (15). Their ability to kill target cells, without prior sensitization, is regulated by the balance between stimulatory and inhibitory signals (16). NK cells express the inhibitory receptors KIRs (killer immunoglobulin-like receptors) that recognize specific HLA class I molecules on target cells, and exert inhibitory signals to NK cells (17). The latter also express activating receptors such as CD16, the NKG2D receptor, and NCR (natural cytotoxicity receptor), involved in tumor lysis (18). Their cytotoxic ability is enhanced by the presence of IL-2 and INF-γ produced by T lymphocytes (19,20). IL-12 seems to enhance NK cell-mediated cytotoxicity and to regulate negatively the proliferation of IL-2–stimulated NK cells (21); IL-15 is the major physiological cytokine present in bone marrow that is important in the development of NK cells (22); and IL-21, secreted by activated CD4 T cells and closely related to IL-15, seems decisive for the end-stage of NK cell activity both by optimizing function and orchestrating elimination (23). The role of interleukins is important in NK cell maturation, but it remains unclear whether CD16+CD56dim or CD16−CD56bright cells represent the precursor cell. Published data suggest that CD56bright cells can be differentiated into CD56dim cells under the influence of IL-2, suggesting that the former may be less mature NK cells (24), while other investigators suggested that CD56dim cells can acquire the CD56bright phenotype upon stimulation with IL-12 (25).
The important role of NK cells in tumor immunosurveillance depends on their ability to fight and destroy cancer cells before the formation of a solid tumor. As previous studies have shown, a decreased number of NK cells and non-functional NK cells are associated with an increased risk of cancer incidence (26) and tumor progression (27).
Thyroid cancer is the most frequent malignancy of the endocrine system. Papillary thyroid carcinoma (PTC), the most common and well-differentiated type of thyroid cancer, is a disease with relatively good prognosis and a low risk of death (28). The role of NK cells has been widely studied in a variety of malignancies such as lung (29,30), ovarian (31), and breast (32) cancer. However, only a few reports on thyroid cancer exist (33).
In a previous study, we have demonstrated an increased number of the total NK cell population in thyroid tissue of patients with PTC as compared to patients with nodular goiter (NG). We also have shown a negative correlation of NK infiltration with disease stage (34).
Considering NK an important population against tumor development, in the present study, we investigated the distribution of CD16+CD56dim and CD16−CD56bright NK subpopulations in patients with PTC and NG, using tissue specimens and peripheral blood samples.
Materials and Methods
Patients
The present study was approved by the local ethical committee and informed consent was obtained from all patients before surgery. Based on the pathology report, 28 patients with PTC (20 females) and 13 patients with NG (12 females) undergoing total thyroidectomy were included in the study.
Peripheral blood and fresh tissue samples were used for the assessment of NK cell subpopulations. Whole blood and thyroid tissue were collected at the time of the surgical procedure. Blood samples from patients were compared to blood samples of 50 healthy blood donors.
Patients with autoimmune diseases, with the exception of Hashimoto's thyroiditis, or other malignancies were excluded from the study.
Blood and tissue preparation
At the time of surgery, 2 mL of whole blood was collected in EDTA-anticoagulated tubes. Thyroid tissue (fresh tissue) from PTC and NG patients was also collected. The blood samples were immediately processed for flow cytometric analysis.
Following thyroidectomy, thyroid tissue was kept immediately on ice until evaluation by the pathologist who selected areas of the tumor or goiter based on macroscopic inspection. The selected sample of thyroid tissue was placed in phosphate-buffered saline (PBS) and was sent to the flow cytometry laboratory. Tissue was minced into small pieces through mechanical disaggregation. Cell surface staining and ex vivo flow cytometry analysis was performed instantly after surgery.
Flow cytometry analysis
Antibodies used for flow cytometry included CD3 (UCHT1), CD16 (3G8), and CD56 (N901), all from Beckman Coulter (Nyon, Switzerland). Data were analyzed on a Cytomics FC 500 Beckman Coulter analyzer, and in every sample, at least 100,000 gated events were measured.
Selective gating by forward versus side scatter was performed to avoid low forward scatter apoptotic cells. Initially, we plotted CD3 versus side scatter, and we selected the CD3−cells. In order to identify NK cells, we plotted the already gated CD3− with CD16 versus CD56. NK cell subpopulations were determined as CD3−CD16+CD56dim and CD3−CD16−CD56bright.
Statistical analysis
Statistical analysis was performed using the GraphPad version 5.0b software. The Mann-Whitney t test and one-way analysis of variance (ANOVA) were used for statistical comparisons. Significance level was set at p<0.05.
Results
Patient population
Twenty-eight patients with PTC (20 females) and 13 patients with NG (12 females) undergoing total thyroidectomy were included in the study. The median ages in the PTC and NG groups were 38.96 (range 17–68) and 51.92 (range 3674) years respectively. Thyroid cancer patients were classified based on the final histology from stage I to IVa with a median range of tumor size of 1.85 cm (0.8–9; Table 1).
PTC, papillary thyroid carcinoma; NG, nodular goiter.
NK cell screening
Blood and tissue samples of patients with PTC and NG and blood samples of healthy blood donors were analyzed for CD16+CD56dim and CD16−CD56bright NK cells (Fig. 1).
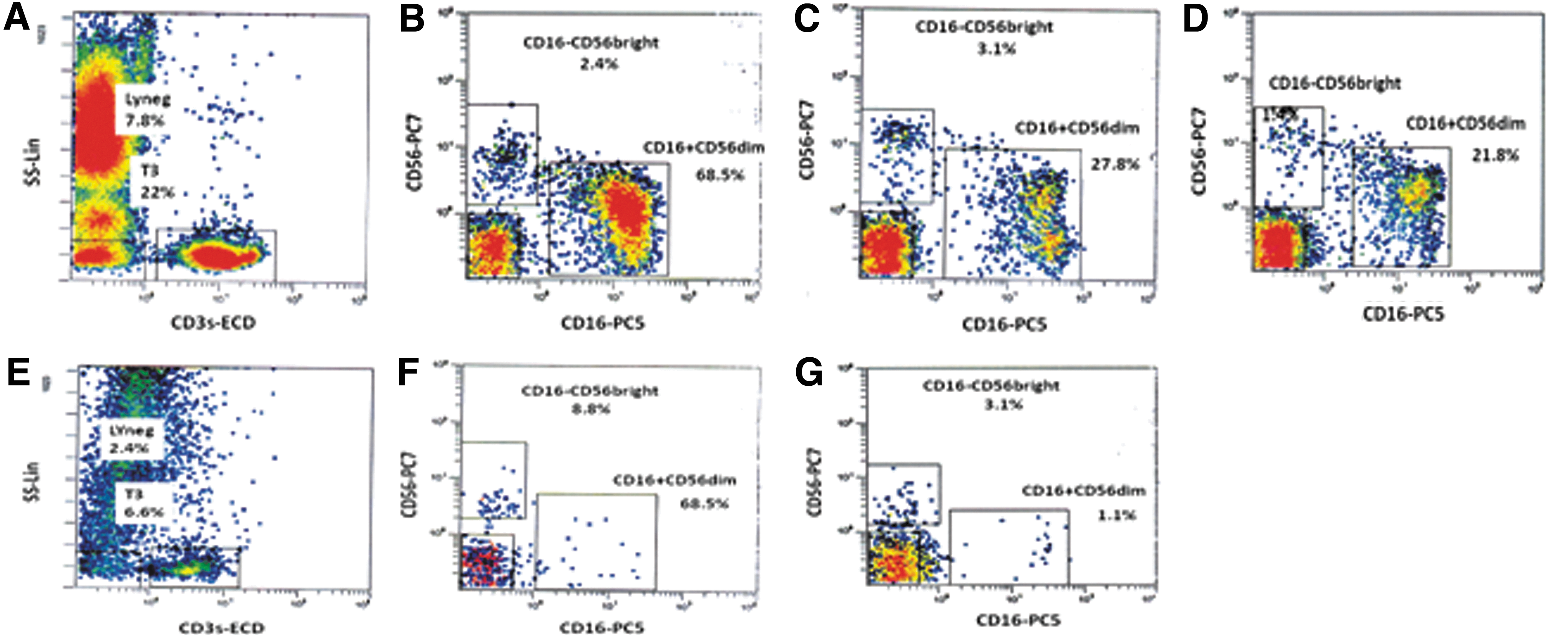
Representative flow cytometric charts are given for natural killer (NK) cell subpopulations in thyroid tissue and peripheral blood from patients with papillary thyroid carcinoma (PTC) and nodular goiter (NG) and peripheral blood from healthy donors:
Flow cytometry results
Both NK cell subpopulations analysis did not show any statistical difference as far as sex and age are concerned, although there was a statistical trend toward decreased infiltration in older patients.
Distribution of NK cell subpopulations in the peripheral blood
The distribution of CD16−CD56bright and CD16+CD56dim NK cell subpopulations in the peripheral blood were similar in patients with PTC, NG, or healthy donors; CD16+CD56dim was the prevailing population as compared to CD16−CD56bright NK cells (p<0.0001, Fig. 2A and B). The distribution of the CD16+CD56dim or CD16−CD56bright populations did not show statistically significant difference among the groups (ANOVA; Fig. 2C and D). Mean percentages and standard deviation (SD) of each group are shown in Table 2.

CD16+CD56dim and CD16−CD56bright NK cells in blood samples:
Values are expressed as mean percentage±standard deviation (SD) of NK cells. Comparisons were made using Mann–Whitney t test.
Statistical significance between the groups is as follows: blood (a/b/c p>0.05, d/e/f p>0.05, a/d p<0.0001, b/e p<0.0001, c/f p<0.0001); tissue (a/b p=0.03, a/c p=0.046, b/d p=0.131, c/d p=0.047).
Distribution of NK cell subpopulations in tissue samples
Distribution of CD16−CD56bright and CD16+CD56dim in PTC and NG
CD16−CD56bright NK cells were increased in the tumor microenvironment of PTC. Analysis of the NK cell subpopulations in the PTC tissue samples revealed that CD16−CD56bright cells were found in higher numbers compared to CD16+CD56dim cells (p=0.046; Fig. 3A). The mean percentage and SD values of CD16+CD56dim and CD16−CD56bright were 35.70±18.79 and 53.71±26.26 (Table 2).
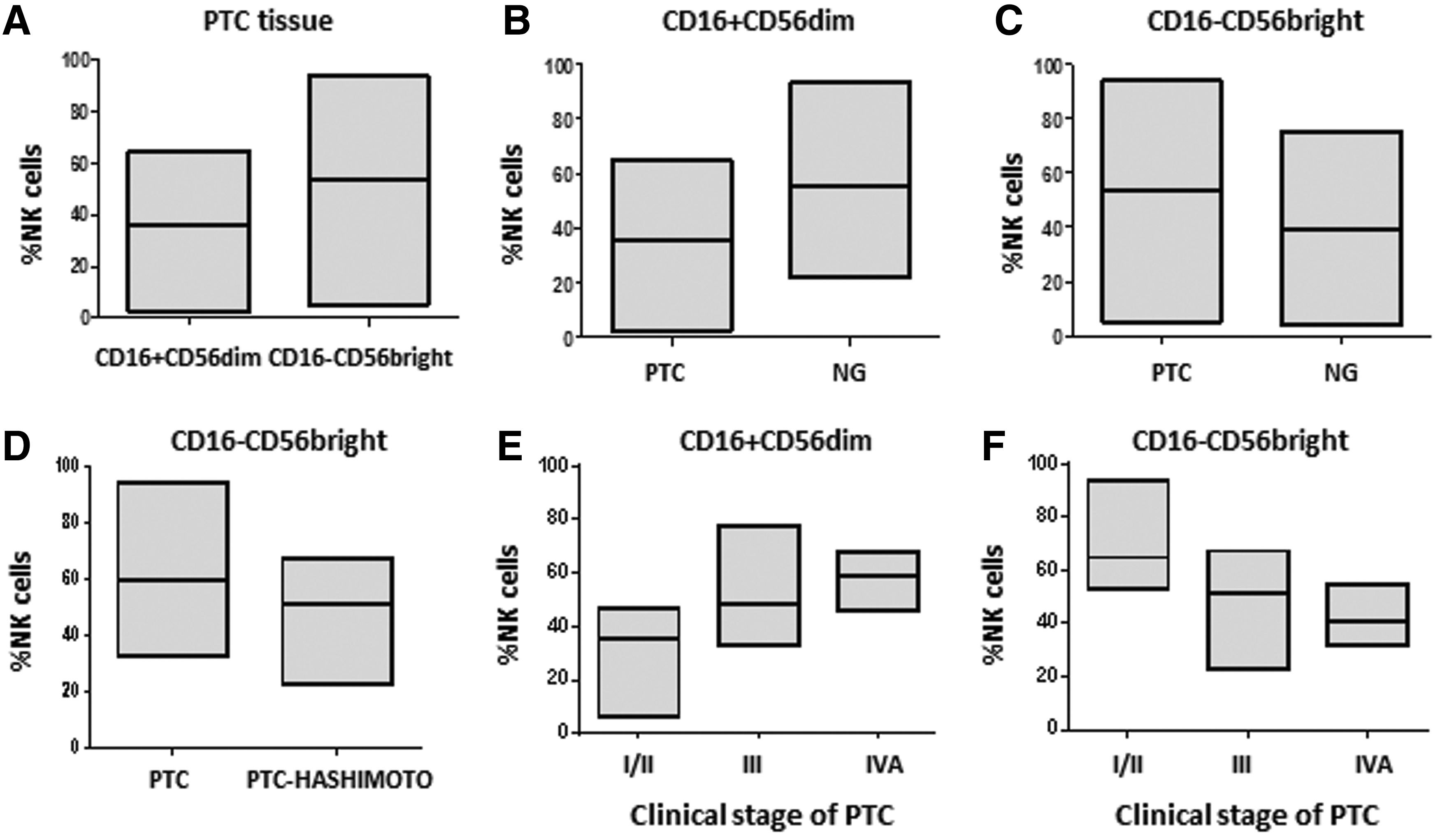
CD16+CD56dim and CD16−CD56bright NK cells in tissue samples of PTC and NG patients:
Analysis of the NK subpopulations in NG tissue samples did not show statistical differences (p=0.131; Table 2).
Comparison of the NK cell subpopulations between PTC and NG revealed that CD16+CD56dim cell numbers were significantly higher in NG than in PTC (p=0.03) (Fig. 3B). In contrast, CD16−CD56bright cells showed a higher infiltration in PTC than NG tissues (p=0.047; Fig. 3C).
Distribution of NK cell subpopulation in relation to tumor stage
CD16+CD56dim and CD16−CD56bright NK cell numbers in patients with stage I/II, III, and IVa are shown in Table 3.
Values are expressed as mean percentage±SD of NK cells. Comparisons were made using Mann–Whitney t test.
Statistical significance between the stages is as follows: CD16+CD56dim NK cells (a/b p=0.016, a/c p=0.0003, b/c p=0.31) and CD16−CD56bright NK cells (a/b p=0.029, a/c p=0.0001, b/c p=0.31).
The number of CD16+CD56dim NK cells positively correlated with disease stage. The mean cell number in stage IVa was doubled compared to that of stages I/II (p<0.0003; Fig. 3E). CD16−CD56bright NK number were inversely correlated with the stage of the disease, with the infiltration of thyroid tissue being greater in stages I/II (p<0.0001; Fig. 3F).
Distribution of CD16−CD56bright and CD16+CD56dim NK infiltrating cells in relation to autoimmune thyroid disease
No significant difference was observed either for CD16+CD56dim (p=0.99) or for CD16−CD56bright (p=0.98) NK cell number in tissue samples of patients with NG versus NG and Hashimoto's thyroiditis. However, a greater infiltration of CD16+CD56dim NK cells was noted in tissue samples of patients with PTC with coexistent Hashimoto's thyroiditis in comparison to patients with PTC alone (mean cell numbers: 39±16.6 and 52±14.6 respectively; p=0.043). In contrast, CD16−CD56bright NK cells were more prevalent in tissue samples of patients with PTC without thyroiditis (mean cell numbers: 60±16.5 vs. 45±14.2 respectively; p=0.044; Fig. 3D).
Discussion
The role of tumor-infiltrating NK cells in cancer microenvironment is evident in many malignancies (35), but NK cell subpopulations have not been extensively studied. More specifically, data analyzing NK cell subpopulations in thyroid cancer, and particularly in PTC, have not been published to date. The goal of the present study was to analyze the distribution of NK cell subsets in thyroid tissue from patients with PTC and NG.
We show for the first time the diversity of NK cell subpopulations in blood and thyroid tissue of patients with PTC and NG. We observed that there is an increased infiltration of CD16−CD56bright NK cells in the PTC tumor microenvironment. In accordance with our study, Carrega et al. (29) showed that tumor-infiltrating NK cells in non-small-cell lung cancer carry the CD16−CD56bright type. In the same study, the authors showed a greater infiltration of CD16−CD56bright NK cells in larger tumors compared to smaller ones. In the present study, a lower infiltration of CD16−CD56bright NK cells correlated with advanced stages of the disease, which may explain the attenuated immune response as the tumor grows (27). In accordance with our data, Gulubova et al. have demonstrated an inverse correlation of CD56+cell infiltration with advanced disease stage (36). Similar results were reported by Ishigami et al. in gastric carcinomas, in which patients with a high level of NK infiltration had a higher rate of early gastric carcinoma and fewer metastases (37). Furthermore Takanami et al. (30) showed that bright NK cell infiltration in lung tumor tissues correlates with a favorable prognosis. However, Jusufovic et al. have reported that increased CD56+NK cells in advanced stages cannot inhibit disease progression (38).
Furthermore, our data demonstrate that the percentage of cytotoxic NK cells (CD16+CD56dim) is lower in tissue samples of patients with PTC and that it correlates with the disease stage. The cytotoxicity of CD16+NK cells is stimulated in the tumor microenviroment, but it seems unable to control tumor proliferation. The activation of NK-cell cytotoxicity is thought to be mediated by a balance of inhibitory and activating NK receptors, as well as various adhesion and co-stimulatory molecules (39).
In peripheral blood, the prevalence of CD16+CD56dim NK cells has been widely established. In our study, the percentage of CD16+CD56dim cells in blood was similar in PTC and NG patients. Our results are in accordance with Belisle et al. (31) who demonstrated similar percentages of CD16+CD56dim NK cells and CD16−CD56bright NK cells in peripheral blood of patients with ovarian cancer in comparison to healthy donors. In contrast, other studies have shown an increased percentage of CD16dim NK cells in the peripheral blood of patients with breast cancer (32) and melanoma (40), as compared to healthy donors. The tumor burden may play a crucial role in these findings.
Hashimoto's thyroiditis is a chronic autoimmune thyroid disease characterized by lymphocytic infiltration, and in many cases, it coexists with PTC. Many previous studies suggest that patients with PTC and coexistent Hashimoto's thyroiditis seem to have a better prognosis and a reduced recurrence risk (33). The proliferation of T and B lymphocytes and the destruction of thyroid tissue is the result of a CD16−NK-cell decrease (41). In the present study, increased numbers of CD16+CD56dim NK cells were observed in patients with PTC and Hashimoto's thyroiditis, although a greater infiltration of CD16−CD56bright NK cells was found in patients with PTC alone. In patients with autoimmune thyroid disease, total CD16+CD56+ NK cells possibly harbor a defect in their cytotoxic effect, and this impairment may lead to expansion of T/B-cell autoimmunity (42).
NK cells migrate from the bone marrow to the peripheral blood and secondary lymphoid tissue (9). The NK cells that migrate to lymphoid organs are CD56bright NK cells and seem to be immature in comparison to CD56dim NK cells (43). Recent studies confirm that CD56bright NK cells are precursors of the CD56dim subset (44
PTC patients who underwent total thyroidectomy and received radioiodine as initial therapy are considered as having a good prognosis with a 10-year survival rate greater than 95% (48). However, there are patients who present recurrence of the disease in a range of 20–30% after initial therapy, depending upon the patient's age and tumor stage at the time of initial treatment (49). The majority (75%) of patients with recurrence develop locoregional neck disease, while the remaining 25% develop distant metastasis (lungs 50%, bones 25%, lungs and bones 20%, or other sites 5%) (50). PTC, when relapsed, has a 10-year survival rate of 49–68% considering neck disease (51 –53), and 36–42% considering distant metastasis (54 –56). Therefore, new treatment modalities should be investigated in patients with advanced cancer, where conventional treatment with surgery and radioiodine therapy has nothing to offer (57), and perhaps NK cell modulation could be amenable for immunotherapies. For instance, activation of endogenous NK cells in a tumor-bearing host and transfer of ex vivo activated NK cells might be beneficial for those patients.
In summary, further studies are needed to investigate the mechanisms underlying the potential role of NK cell subpopulations in thyroid cancer. Moreover, the positive correlation of CD16+CD56dim NK cells with disease stage suggests that these cells are unable to suppress tumor progression. A novel approach in cancer immunotherapy includes NK cell expansion or stimulation by other immune-related cells. Although PTC is generally considered a tumor with a favorable prognosis, a better understanding of immune responses and their impact on tumor progression may contribute to the development of effective and less toxic immunotherapeutic options.
Footnotes
Acknowledgment
This study was financially supported by a grant of the Hellenic Endocrine Society.
Author Disclosure Statement
The authors declare that no competing financial interests exist.