
Abstract
Background:
It is controversial whether persistent signaling by the thyrotropin (TSH) receptor (TSHR) is cell-type specific. We reported persistent TSHR signaling in human embryonic kidney 293 (HEK293) cells expressing human TSHRs (HEK-TSHRs), whereas another group reported persistent signaling in mouse thyroid follicles but not in HEK293 cells. Herein, we test this hypothesis directly.
Methods:
We used two methods to measure persistent signaling in HEK-TSHRs and confirm our previous observations. In Method 1, we used a chemiluminescent immunoassay to measure intracellular cAMP accumulation over 30–60 min by adding a phosphodiesterase inhibitor to the incubation medium. In Method 2, we used an intracellular biosensor to record cAMP levels continuously.
Results:
Using Method 1, we show that TSHR signals persistently in human thyrocytes and human osteosarcoma U2OS-TSHR cells. Using Method 1 in HEK-TSHRs, we show that after 5 min, the phosphodiesterase inhibitor isobutylmethylxanthine (IBMX) increases cAMP to 2.5 pmol/well, TSH increases cAMP to 1.6 pmol/well, but IBMX added 30 min after TSH withdrawal increases cAMP to 105 pmol/well. Using Method 2 in HEK-TSHRs, we confirm that without IBMX, TSH causes a transient increase in cAMP and 30 min after TSH withdrawal, IBMX increases cAMP in cells pretreated with TSH more rapidly and to a higher level than IBMX added to cells not pre-exposed to TSH. Lastly, using Method 2, we show that in HEK-TSHRs phosphodiesterases types 3 and 4 are involved in degrading cAMP as the specific inhibitors Rolipram and Milrinone expose persistent TSHR signaling.
Conclusions:
We conclude that persistent TSHR activation occurs in human thyrocytes, U2OS-TSHR cells and HEK-TSHRs; it is not cell-type specific but is revealed by inhibiting phosphodiesterases.
Introduction
Herein, we show that the TSHR signals persistently in primary cultures of human thyrocytes, in U2OS-TSHR cells, and HEK-TSHRs. We use two approaches to measure persistent cAMP production in HEK-TSHRs and confirm our previous observations that the TSHR signals persistently in HEK293 cells. Moreover, we show that the transient nature of the increase in cAMP levels in HEK-TSHRs is caused by its degradation by phosphodiesterase (PDE) activity, as inhibition of PDEs exposes the persistent activity of TSHRs even 30 min after TSH withdrawal.
Materials and Methods
Cell culture
Primary cultures of human thyrocytes were established and HEK-TSHRs generated as described previously (9). U2OS cells, a human osteosarcoma line, was transfected to express TSHRs stably (U2OS-TSHR cells). All cell types were maintained in Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% fetal bovine serum (FBS), 100 U/mL penicillin, and 10 μg/mL streptomycin (Life Technologies Corp., Carlsbad, CA) at 37°C in a humidified 5% CO2 incubator.
Measurement of cAMP levels
Method 1: Measurement of cAMP production in cell monolayers
Thyrocytes (0.8×105 cells/well), or HEK-TSHRs (2.2×105 cells/well) were seeded into 24-well plates, U2OS-TSHR cells (1.1×105 cells/well) into 12-well plates in DMEM containing 10% FBS. Cells were cultured overnight before incubation for 30 min in HBSS/10 mM HEPES, pH 7.4. To determine the immediate effects of TSH stimulation, cAMP production was measured in cells incubated for 30 min at 37°C in a humidified incubator in HBSS/HEPES containing 1 mM 3-isobutyl-1-methylxanthine (IBMX; SIGMA, St. Louis, MO) in the presence or absence of bovine TSH (bTSH; SIGMA) as described previously (9). To determine the persistent effect of TSH on cAMP production (2), cells were incubated without (control group) or with TSH for 30 min (pretreatment group), washed with HBSS/HEPES three times at RT, and incubated in HBSS/HEPES at 37°C. After 30 or 60 min, cells were incubated in 0.2 mL HBSS/HEPES with 10 mU/mL bTSH or with 1 mM IBMX or 1 mM IBMX and 10 mU/ml bTSH at 37°C for 30 min (treatment group). Incubations were stopped, and the cells were lysed by adding 0.2 mL lysis buffer of the cAMP-Screen Direct™ System (Applied Biosystems, Foster City, CA) at the times indicated. The cAMP content of the cell lysate was determined using the method described in the manufacturer's protocol. The chemiluminescence signal was measured in a VICTOR3™ V 1420 Multilabel Counter (Perkin Elmer, Waltham, MA). In some experiments, the PDE3- and PDE4-specific inhibitors, milrinone and rolipram respectively, were used instead of IBMX. (A line graph of this protocol is presented as fig. 2 in ref. 4.)
Method 2: Measurement of intracellular cAMP levels in living cells
HEK-TSHR cells were seeded into glass-bottomed MatTek dishes (200,000 cells per dish) in growth media (DMEM, 10% FBS, 1% penicillin/steptomycin). After 20 h, cells were transiently transfected by FuGENE 6 (Roche Applied Sciences, Indianapolis, IN) with a pCDNA3 mammalian expression plasmid carrying the gene for the FRET-based, cAMP biosensor YR-ICUE (10,11). After 20 h, HBSS supplemented with 20 mM HEPES, pH 7.5 replaced growth media, and cells were imaged on a Zeiss 510 NLO/Meta system, using a Plan-Neofluor 40×/N.A. 1.3 objective (Carl Zeiss, Oberkochen, Germany). The pinhole was completely open to generate epifluorescent images. YFP was excited with a 515 nm laser at 30% transmission. YFP and YFP to RFP FRET (YR FRET) fluorescence were detected via the Meta detector with the following settings: YFP (533–554 nm), FRET (597–650 nm). Detector gains were set at 350 for both YFP and YR FRET channels. Detector gains and microscope parameters remained unchanged throughout all experimental conditions. Images were acquired at 30 s intervals, and a baseline FRET ratio was established at which point IBMX and/or bTSH were added by complete discontinuous buffer exchange as indicated.
Measurement of PDE activity
PDE activity was measured using the Cyclic Nucleotide Phosphodiesterase assay kit (Enzo Life Sciences Inc., Farmingdale, NY) according to the vendor's instructions. We measured the specific PDE activity by quantifying phosphate levels after 15 and 30 min and subtracting the phosphate levels found in the absence of 5′-nucleotidase conversion of cAMP to inorganic phosphate.
Statistical analysis
Analysis was performed by t test, with a p value of <0.05 considered significant.
Results
Persistent cAMP production in human thyrocytes and U2OS-TSHR cells was measured by Method 1. Figure 1 illustrates that thyrocytes from three different donors exhibited persistent cAMP production. cAMP production 60 min after bTSH withdrawal was 3.8-fold higher than in the control group. By contrast, bTSH acutely stimulated cAMP production 45-fold. In U2OS-TSHR cells, persistent cAMP production was 5.4-fold higher than in the control group, whereas acutely stimulated cAMP production was 7.1-fold higher than in the control group (Fig. 1). These findings show that human thyrocytes exhibit persistent TSHR activation as was previously found in mouse thyroid follicles (3), as do U2OS cells made to express TSHRs.
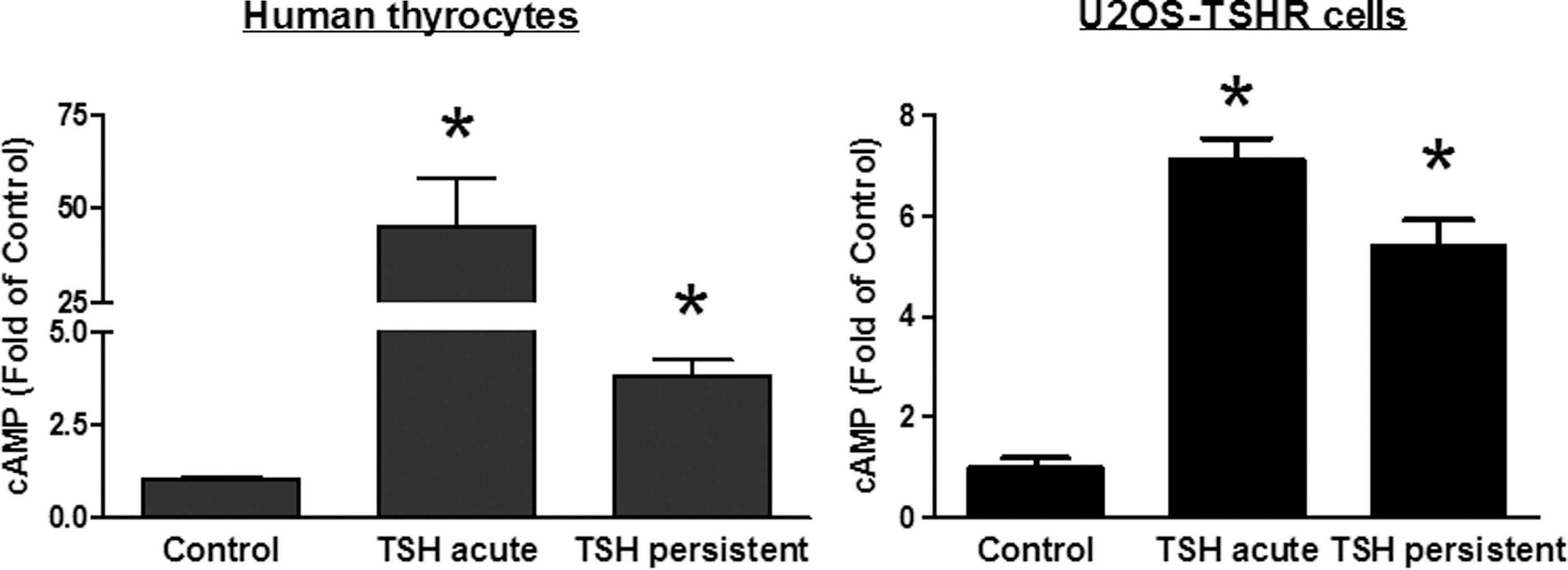
Persistence of thyrotropin (TSH)-stimulated cAMP production in primary cultures of human thyrocytes and U2OS-TSHR cells. Left panel: Thyrocytes were incubated without or with 10 mU/mL bTSH. After 30 min, the cells were washed three times with buffer and then incubated in buffer at 37°C. After an additional 30 min, cells not pretreated with TSH were incubated in HBSS/HEPES with 0.5 mM IBMX (control) or IBMX and 10 mU/mL bTSH (TSH acute) and then incubated at 37°C. Cells pretreated with bTSH were incubated in HBSS/HEPES with 0.5 mM IBMX (TSH persistent). After an additional 60 min, the cells were lysed and cAMP was measured in the cell lysates. The bars are the mean±SE of two to four measurements in experiments with thyrocytes from three different donors. Right panel: U2OS-TSHR cells were incubated without or with 10 mU/mL: bTSH. After 30 min, the cells were washed three times with buffer and then incubated in buffer at 37°C. After an additional 60 min, cells not pretreated with TSH were incubated in HBSS/HEPES with 0.5 mM IBMX (control) or IBMX and 10 mU/mL bTSH (TSH acute) and then incubated at 37°C. Cells pretreated with bTSH were incubated in HBSS/HEPES with 0.5 mM IBMX (TSH persistent). After an additional 60 min, the cells were lysed and cAMP was measured in the cell lysates. The bars are the mean±SE of three measurements in two experiments. *p<0.05 compared to control.
bTSH-stimulated cAMP levels in HEK-TSHRs were measured using Method 1, which is the method we used in our previous report (2), and Method 2, which is similar to that used by Werthmann et al. (3). Figure 2 compares acute and persistent bTSH-stimulated cAMP levels measured by Method 1. Basal cAMP in these cells was 0.1 pmol/well. In the absence of IBMX, acute administration of TSH stimulated a transient increase in cAMP that was 1.6 pmol/well after 5 min, peaked at 6.1 pmol/well at 30 min and decreased by 60 min (Fig. 2, upper left). IBMX alone increased cAMP to 2.5 pmol/well after 5 min, peaked at 8.0 pmol/well at 30 min, and decreased by 60 min (Fig. 2, upper right). Acute administration of bTSH in the presence of IBMX stimulated a robust increase in the level of cAMP that was 23.7 pmol/well after 5 min and continued to increase to 241 pmol/well by 60 min (Fig. 2, lower left). Administration of IBMX 30 min after the withdrawal of bTSH (persistent signaling) also caused a marked increase in the cAMP level to 105 pmol/well after 5 min and continued to increase to 175 pmol/well by 60 min (Fig. 2, lower right). The cAMP level stimulated by bTSH in the presence of IBMX during acute stimulation is not very different from the levels attained when persistent signaling is revealed by administration of IBMX, although the kinetics of accumulation are different. cAMP accumulated at a faster rate during the first 5 min of persistent signaling than during the same period of acute stimulation. These data show that TSHRs continue to produce cAMP at a high rate 30 min after bTSH withdrawal when measured in the presence of IBMX, thereby confirming that TSHRs remain activated after bTSH withdrawal.
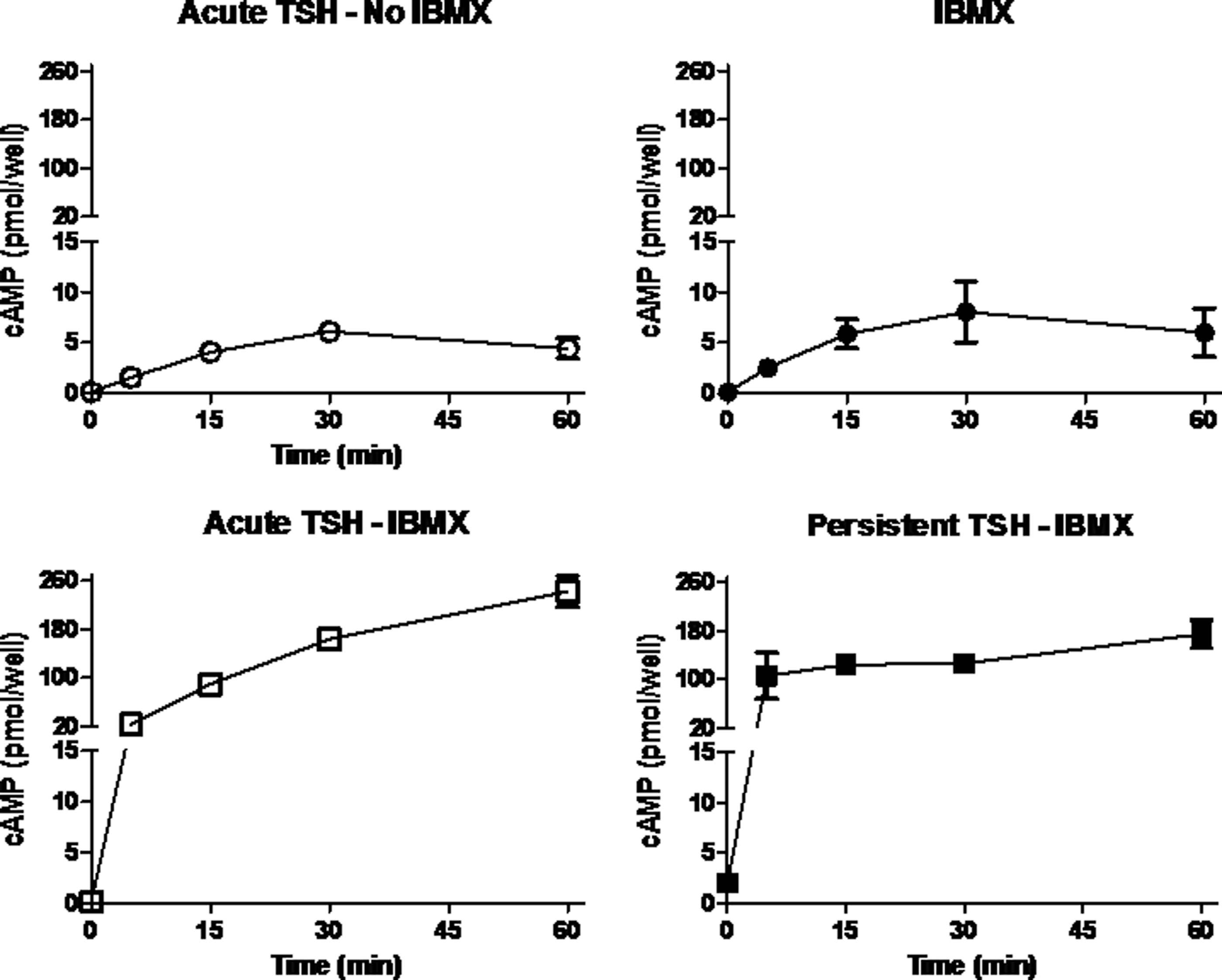
Persistent TSH-stimulated cAMP production in HEK-TSHRs using Method 1. HEK-TSHR cells were incubated without or with 10 mU/mL bTSH (TSH pretreatment). After 10 min at 37°C, the cells were washed three times with buffer and then incubated in buffer at 37°C. After 30 min, cells not pretreated with bTSH were incubated in HBSS/HEPES 10 mU/mL bTSH (acute TSH—no IBMX) or in HBSS/HEPES with 0.5 mM IBMX (IBMX) or in HBSS/HEPES with bTSH and IBMX (acute TSH—IBMX), and cells pretreated with bTSH were incubated in HBSS/HEPES with 0.5 mM IBMX (persistent TSH—IBMX) at 37°C. After 30 min, the cells were lysed and cAMP was measured in the cell lysates. The data (mean±SE) are from two experiments performed in triplicate.
Figure 3 illustrates the effect of acute addition of bTSH in the absence of IBMX, followed by a washout and then addition of IBMX, and compares it to the effect of IBMX alone using Method 2. We used HEK-TSHRs transiently expressing the FRET-based cAMP biosensor as previously described (12). The concentration of IBMX used in these experiments was chosen after determining the lowest IBMX concentration that shows no increase in cAMP in parental HEK293 cells (not shown) and a modest increase in cAMP in HEK-TSHR cells, which is almost certainly caused by the basal signaling of TSHRs (Fig. 3, lower tracing). In contrast to the results obtained with Method 1 that measures cAMP levels accumulated over a period of time, this method allows detection of intracellular cAMP dynamics within living cells in real time. Moreover, this method allows for the measurement of changes in cAMP levels stimulated by bTSH in the absence of IBMX followed directly by the effects of administered IBMX within the same group of cells. Figure 3 (upper tracing) illustrates the results of an experiment in which cells were stimulated by 10 mU/ml bTSH for 3 min. The bTSH was then “washed out” and the cells incubated in buffer followed by addition of 0.06 mM IBMX. The effect of 0.06 mM IBMX in cells that had not been incubated with bTSH is shown (Fig. 3, lower tracing). Stimulation by bTSH in the absence of IBMX caused a transient elevation of cAMP that peaked after 11 min. Addition of IBMX 30 min after withdrawal of bTSH caused a second increase in cAMP that exhibited a much more rapid rise and was of greater magnitude (Fig. 3, upper tracing) than that obtained in cells not treated with bTSH (Fig. 3, lower tracing) that was sustained for at least 80 min. These data show that persistent cAMP production could be revealed by adding IBMX in Method 2 as in Method 1.
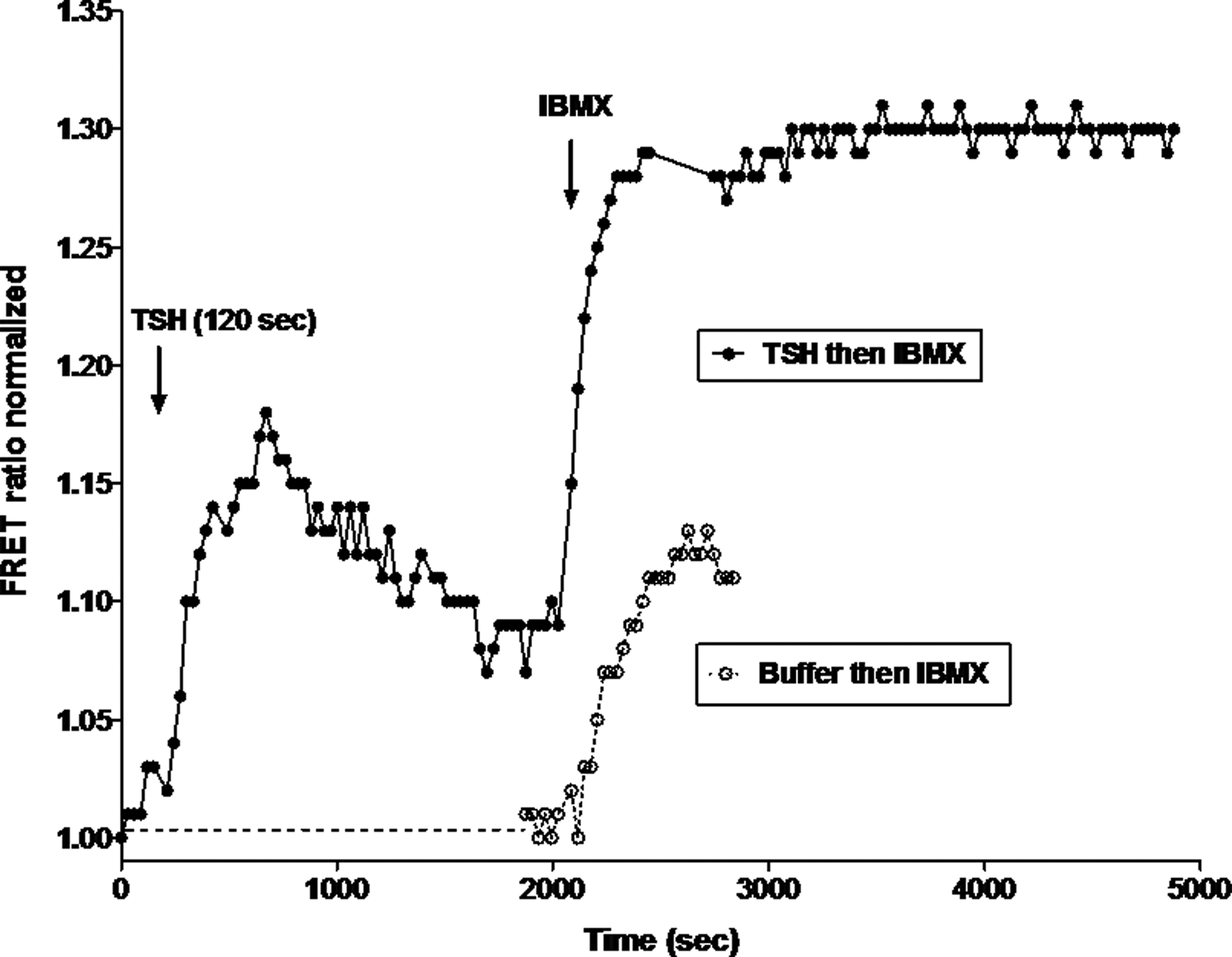
Persistent TSH-stimulated cAMP levels in HEK-TSHRs using Method 2. Continuous measurement of cAMP levels was made in HEK-TSHRs transiently expressing the FRET-based cAMP biosensor ICUE-IR (10). Upper tracing: Representative time course of YFP to RFP emission ratio changes in cells stimulated with 1 mU/mL bTSH for 3 min followed by a washout for 30 min and then addition of 0.06 mM IBMX (upper tracing). TSH stimulated an increase in cAMP that was followed by a decrease after washout. Addition of IBMX 30 min after washout caused a second increase in cAMP levels. Lower tracing: Representative time course of the addition of 0.06 mM IBMX to cells that had not been exposed to bTSH.
To gain insight into which PDEs may be active in HEK-TSHR cells and to confirm that the effect of IBMX was due to its inhibition of PDE activity, we determined whether milrinone or rolipram would expose persistent signaling. Figure 4 shows that both milrinone and rolipram at the doses used expose persistent TSHR-mediated cAMP signaling. Of note, milrinone had a greater effect than rolipram on basal cAMP production whereas the reverse was found for TSH-stimulated cAMP production, and the effects of milrinone and rolipram were additive.

Effects of milrinone and rolipram to expose persistent TSH-stimulated cAMP production in HEK-TSHRs using Method 1. The experiment was performed as in the legend to Figure 2, except the PDE inhibitor was either IBMX, milrinone, rolipram, or milrinone+rolipram (all at 1.0 mM). The data (mean±SE) are from two experiments performed in triplicate.
Discussion
In this report, we have attempted to resolve the controversy as to whether persistent signaling by the TSHR is cell-type specific—in particular, whether it occurs in thyroid cells but not in HEK293 cells or U2OS cells. We first show that TSHRs signal persistently via the cAMP pathway in primary cultures of human thyrocytes, thus confirming the finding reported by Calebiro et al. in mouse thyroid follicles (1), and in U2OS cells made to express TSHRs. We used two approaches to determine whether the TSHR signals persistently in HEK293 cells, even though we had reported previously that TSHR signals persistently via both the cAMP (2) and phosphoinositide pathways in these cells (4). It is important to note that in both of our previous reports, we measured persistent activation of the TSHR by quantifying the production of cAMP or inositol phosphates after TSH withdrawal by inhibiting their degradation by PDEs using IBMX or inositol phosphatases using LiCl respectively. This is Method 1 for cAMP signaling in this report. This approach allows for measurement of receptor activation more sensitively than the approach employed by Calebiro et al. (1) because the readout for receptor activation is not affected by the activities of the degrading enzymes, PDEs for cAMP or inositol phosphatases for inositol monophosphates. In Method 2, we used an approach similar to that employed by Calebiro et al. (1) in which we measured cAMP levels by a FRET-based cAMP biosensor (10,11) in real time in the absence of a PDE inhibitor. We show persistent cAMP elevation in the absence of IBMX. In contrast to the measurements made by these other investigators, we added administration of IBMX to our protocol at the end of the real-time measurements to overcome any reduction in cAMP levels caused by PDE-mediated degradation. We show that addition of IBMX caused a robust increase in cAMP in cells previously exposed to TSH. In both methods, addition of IBMX revealed the persistent activation of TSHRs in HEK293 cells as shown by continued increases in cAMP levels when cAMP degradation was inhibited. Furthermore, we show that the PDE3-specific inhibitor milrinone and the PDE4-specific inhibitor rolipram both revealed persistent TSHR cAMP signaling, thus confirming that the effect of IBMX was due to PDE inhibition. The effects of the specific PDE inhibitors suggest that degradation of cAMP in HEK-TSHR cells is mediated by both PDE3 and PDE4. Thus, we can conclude that TSHRs exhibit persistent signaling in HEK293 cells.
We chose to employ two methods to measure persistent signaling because they measure this activity in different ways. In Method 1, we measure the increase in the accumulation of cAMP over a set time frame in a population of cells. By inhibiting cAMP degradation with a PDE inhibitor, we were able to quantify the increase in cAMP accumulation that is stimulated by an activated TSHR. For example, the level of cAMP increased from less than 1 pmol/well in control cells to 3 pmol/well in cells 5 min after addition of IBMX. In contrast, the level of cAMP increased from 2.5 pmol/well in cells that had been exposed to TSH for 3 min and from which TSH had been withdrawn 30 min before to 105 pmol/well 5 min after addition of IBMX. The difference in the magnitudes of the increases with addition of IBMX revealed the continuing activity of TSHRs. Method 1 thus allowed us to measure the magnitude of the increase in cAMP production produced by persistently activated TSHRs. By contrast, Method 2 measures the level of cAMP in compartments within individual cells in which the FRET biosensor is expressed in real time. Thus, Method 2 allows measurement of differences in individual cells and provides a continuous analysis of these changes. However, it does not allow for quantitation of the magnitude of the changes in cAMP levels. Addition of IBMX to cells that had been treated with TSH led to increases in cAMP that were more rapid and robust than addition of IBMX to control cells. Thus, we conclude that these two methods are complementary.
An uncertainty regarding persistent TSHR signaling that has not been resolved pertains to whether persistently active TSHRs remain in a complex with TSH. In their recent paper (3), Werthmann et al. showed that there is a correlation between the loss of a FRET signal between TSH and the TSHR and a decrease in cAMP levels, and the authors interpreted this to mean that a persistently active TSH/TSHR complex is necessary for continued stimulated production of cAMP. We do not think this shows that the active signaling complex must contain TSH bound to the TSHR because we think there is persistent TSHR activation in their experiments even though they have not revealed it. In a previous report (2), we used a specific small molecule TSHR inverse agonist (13), and were able to show that an activated TSHR was needed for persistent cAMP signaling. Our previous data, however, could not be used to determine whether the activated TSHR remained bound to TSH because the inverse agonist could have inhibited signaling by an activated receptor that no longer had TSH bound. A previous study using a neutral antagonist was inconclusive (4). A future study using a more suitable neutral antagonist might provide evidence that would allow this question to be resolved.
A third aspect of persistent TSHR signaling that remains controversial pertains to whether TSHRs must be internalized in order for them to exhibit persistent activation. Both we (2,4) and Werthmann et al. (3) find that TSHRs are poorly internalized in HEK293 cells and that the level of internalization can be increased by co-expressing β-arrestin 2. Using their method, Werthmann et al. found no persistent cAMP signaling even after TSHR internalization was increased. In contrast, we found that neither persistent TSHR cAMP signaling (2) nor persistent phosphoinositide signaling (4) was affected by increasing internalization by co-expressing β-arrestin 2 or inhibiting internalization by co-expressing a dominant-negative dynamin. We therefore concluded that persistent TSHR signaling is independent of β-arrestin 2-mediated internalization.
We conclude that TSHRs exhibit persistent signaling in HEK293 cells, U2OS cells, and human thyrocytes. Thus, persistent TSHR signaling does not appear to be cell-type specific and may occur in all cell/tissues in which TSHRs are expressed.
Footnotes
Acknowledgments
This research was supported by the Intramural Research Program of the NIH (1 Z01 DK011006). We thank Bernice Samuels for her excellent technical assistance and Alisa Boutin for generating U2OS-TSHR cells.
Author Disclosure Statement
The authors have nothing to disclose.