
Abstract
At thyroid hormone response elements on specific genes, complexes of nuclear thyroid hormone receptors (TRs) and 3,5,3′-triiodo-L-thyronine (T3), coactivator or corepressor nucleoproteins, and histone acetylases or deacetylases mediate genomic effects of the hormone. Nongenomic effects of the hormone are those whose initiation does not primarily depend upon formation of the TR–T3 complex. Among the nongenomic effects of thyroid hormone are a set of actions initiated at a cell surface receptor on integrin αvβ3 that are relevant to a) intracellular trafficking of proteins, including TRβ1, b) serine phosphorylation and acetylation of this nuclear receptor, c) assembly within the nucleus of complexes of coactivators and corepressor, and d) transcription of specific genes, including that for TRβ1. These actions initiated at αvβ3 are reviewed here and appear to be adjunctive to the genomic actions of the TR–T3 complex.
Introduction
G
The cell surface receptor for thyroid hormone and hormone analogs has been defined on a structural protein of the plasma membrane, integrin αvβ3 (6 –8). This receptor distinguishes T3 from L-thyroxine (T4), and both of these from the deaminated analog of T4, tetraiodothyroacetic acid (tetrac). The receptor has two hormone-binding domains at which hormone signals are transduced by either phosphatidylinositol 3-kinase (domain S1) or mitogen-activated protein kinase (MAPK) (domain S2) into discrete downstream nuclear events (8,9) (Fig. 1). Among these effects are MAPK-dependent serine phosphorylation of TRβ1 (10,11) and MAPK-mediated acetylation of this nuclear receptor (12). Actions of thyroid hormone at αvβ3 also include trafficking of proteins in the cytoplasm to the nucleus (13 –15). Among these proteins imported by the nucleus in thyroid hormone–treated cells are TRs, the nuclear estrogen receptor-α (ERα), p53, and signal transducing proteins, such as signal transducer and activator of transcription-1α (STAT1α). Finally, T4 may induce internalization of heterodimeric αvβ3 from the plasma membrane and nuclear uptake of monomeric αv (16), where the latter functions as a previously unrecognized coactivator protein involved in the transcription of TRβ1 and several other genes.
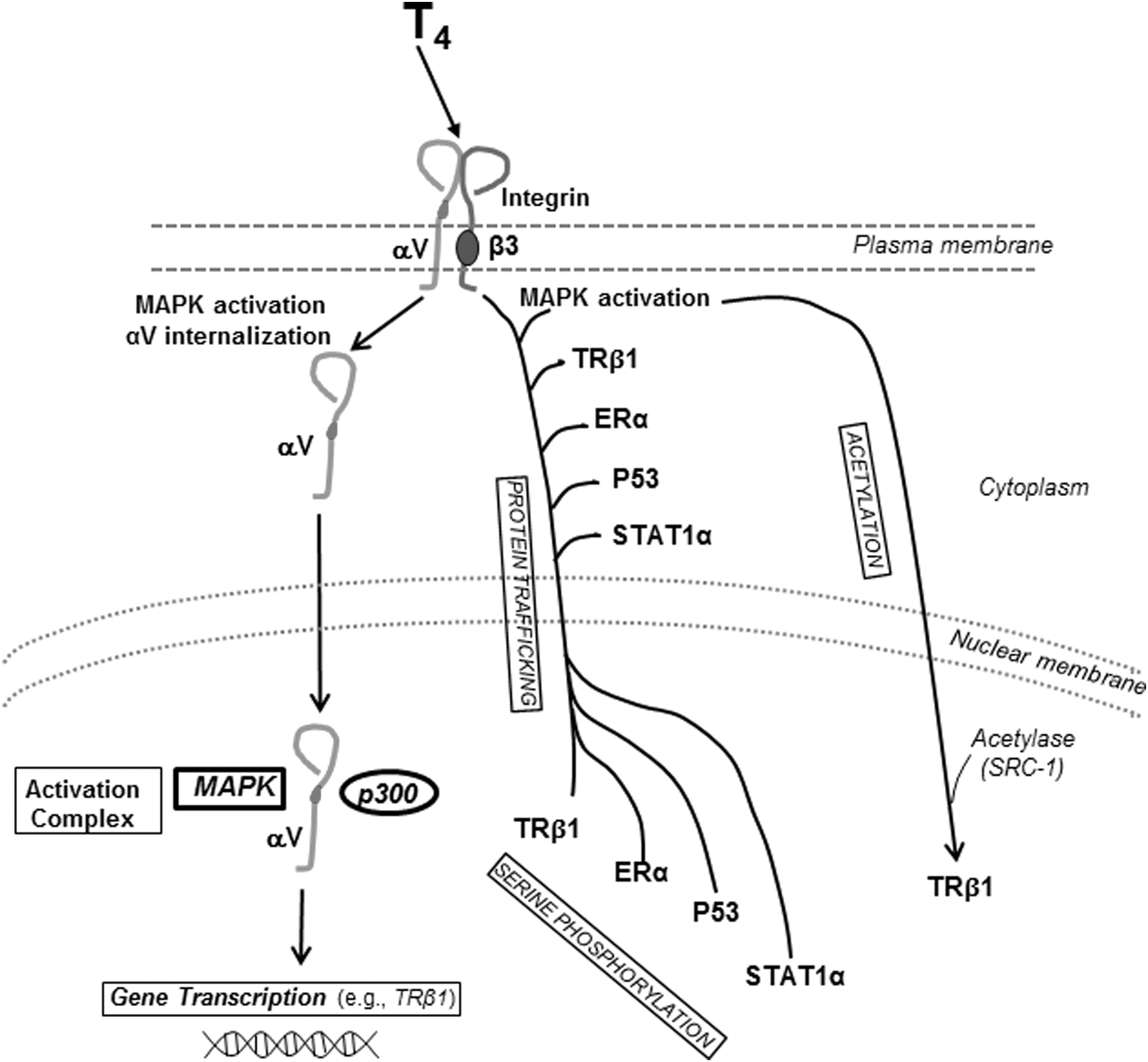
Diagram of nongenomic actions of thyroid hormone initiated at heterodimeric integrin αvβ3 that modify state or function of nucleoproteins, including TRβ1. The integrin is a structural protein of the plasma membrane. In pathway 1, thyroid hormone (L-thyroxine, T4) via activation of mitogen-activated protein kinase (MAPK) causes internalization of and nuclear uptake of monomeric αv. In the nucleus, αv forms a coactivator complex with p300, activated MAPK, and other proteins that leads to transcription of genes that include TRβ1 (16) (see Assembly in the cell nucleus of coactivator complexes). In pathway 2, T4-activated MAPK promotes nuclear importation from the cytoplasm and concurrent serine phosphorylation of TRβ1 and other proteins—estrogen receptor-α (ERα), p53, and STAT1α. These proteins are activated by specific serine phosphorylation. In pathway 3, thyroid hormone–directed MAPK causes acetylation via steroid receptor coactivator-1 (SRC-1) of TRβ1. Acetylation may be required for recruitment of p300 to the MAPK–αv complex in the nucleus (pathway 1). T3 also binds to the hormone receptor on αvβ3 and is capable of activated MAPK, but the affinity of the receptor for T3 is lower than that for T4 (6), and thus its contributions to these pathways are less clear.
This review summarizes the effects on thyroid hormone on TR that originate at the plasma membrane and αvβ3. These actions are nongenomic in mechanism and appear to be adjunctive to genomic effects of T3. Thyroid hormone at the receptor on integrin αvβ3 is largely T4, because the affinity of the receptor is higher for T4 than for T3 (6) and near-physiologic for free T4. We have elsewhere reviewed the actions of thyroid hormone and tetrac at αvβ3 that relate to cancer cell proliferation and angiogenesis (1,17,18). These actions do not primarily involve TRs.
Literature Review
Trafficking of TR from the cytoplasm to the nucleus
Zhu et al. (14) and Maruvada and co-workers (13) demonstrated the residence of nuclear TRβ1 in the cytoplasm and transfer of the receptor to the nuclear compartment. The model system consisted of the expression of a green fluorescent protein TR chimera into cells that lack endogenous TR. Zhu et al. also showed that T3 drove cytoplasmic TR into the nucleus. We subsequently reported that shuttling of transfected TR from the cytoplasm to the nucleus directed by thyroid hormone in CV-1 cells (15) was a process initiated at the cell surface receptor for thyroid hormone on integrin αvβ3 (6). Anti-αvβ3 and tetrac, an inhibitor of the binding of T4 and T3 to the integrin (8), blocked nuclear uptake of TRβ1 in hormone-treated cells. T4 was used in these studies in the presence of propylthiouracil that inhibited its deiodination to T3. The S2 domain within the hormone receptor on αvβ3 binds both T4 and T3, and the S1 domain recognizes only T3 (9).
A number of studies have shown that thyroid hormone can activate MAPK (ERK1/2) via plasma membrane αvβ3 (8,10). When activated by T4 or T3, the enzyme has a variety of intracellular functions, including serine phosphorylation of TRβ1 (10,11). In studies of trafficking of TR stimulated by T4, we found TR and MAPK to be complexed in the nucleus. Pharmacologic inhibition of MAPK with PD98059 prevented nuclear uptake of both TR and MAPK that was caused by thyroid hormone, and the inhibitor expectedly blocked phosphorylation of the receptor. In these experiments, the concentration of free T4 was within the physiologic range. Thus, activation of MAPK is essential to the shuttling of TRβ1 from the cytoplasm to the nucleus, and to the formation of a kinase–TR complex that appears to be required for importation. The complex of activated MAPK and TRβ1 is found in the cytoplasm (19), and thus we presume that nuclear uptake involves the preformed complex. In the course of this cytoplasm-to-nucleus trafficking, TR is phosphorylated. Interestingly, T3, apparently acting at S1 on heterodimeric αvβ3, drives TRα1 into the nucleus (9) by a mechanism that involves phosphatidylinositol 3-kinase. Thus, independent control exists at αvβ3 for the nuclear importation of TRβ1 and TRα1. This is not surprising, given the discrete roles of the two receptors (1). Of course, additional pathways may exist in cells for driving nuclear uptake of these two TRs.
It is not known whether other proteins of cytoplasmic origin participate in the complex of MAPK and TRβ1 that is directed from the integrin in T4-exposed cells. However, it is clear that enhanced protein trafficking from the cytoplasm to the nucleus in thyroid hormone–treated cells includes nuclear receptors for other hormones, such as ERα (20), as well as STAT-1α (21) and STAT3 (22). Once the intranuclear compartment is reached by the MAPK–TRβ1 complex in T4-treated cells, however, nuclear coactivator proteins are a part of the complex, as is, surprisingly, monomeric integrin αv (16) that can have coactivator activity.
Modulation of transcription of specific genes directed from αvβ3 by thyroid hormone
The genomic regulation of gene transcription by nuclear T3 and nuclear TRs in conjunction with coactivator and corepressor proteins is regarded as the principal mechanism by which thyroid hormone influences gene expression (1 –5). However, activation of integrin αvβ3 is known to regulate expression of a varied spectrum of genes, including epidermal growth factor receptor (23), tissue inhibitor of metalloproteinase-1 (24), vitronectin (25), transforming growth factor-β (26), and integrin-linked kinase (27). The ligand binding by αvβ3 of thyroid hormone and hormone analogs, particularly, tetrac—molecules with molecular weights of less than 1000 Da—has revealed a distinctive panel of gene transcription effects (8,16). These effects have been profiled in tumor cells that generously express integrin αvβ3, for example, human breast cancer (28) and glioblastoma cells (9), and in endothelial cells (29). The gene profiles are relevant to cancer cell survival pathways and to angiogenesis. Nonmalignant cells and nondividing blood vessel cells have reduced αvβ3 expression/activity compared with tumor cells or rapidly dividing endothelial cells.
Table 1 is a partial list of genes whose expression is affected by a nanoparticulate formulation of thyroid hormone analog tetrac that cannot enter cells and acts exclusively at αvβ3 (28 –30). The products of these genes are relevant to control of the cell cycle, apoptosis, angiogenesis, and cell-to-cell adhesion. As shown in Table 2, action of T4 at the cell surface receptor includes regulation of expression of genes for nuclear hormone receptors for TRβ1 and ERα and the COX-2 and HIF-1α genes, as noted above. When we compare Tables 1 and 2 with gene expression profiles compiled for liganded nuclear TRs (4,5), little or no overlap is apparent; this suggests that the panels of genes regulated from the integrin by thyroid hormone/hormone analogs and by the nuclear receptor–T3 complex are largely discrete. However, expression of the TRβ1 gene—and, as noted above, the trafficking of this receptor—can be influenced by thyroid hormone from the cell surface integrin to determine abundance and location of the nuclear TR protein. It should be noted that T3 can also act genomically to transcribe TRβ1 (31). Thus, even though there is little concurrence of transcriptional events initiated at the TR on the integrin and transcriptional events initiated within the nucleus by thyroid hormone, control of TRβ1 expression is one example of transcriptional redundancy.
Tetrac, tetraiodothyroacetic acid.
Assembly in the cell nucleus of coactivator complexes relevant to TR
Genomic actions of thyroid hormone include assembly in the cell nucleus of coactivator or corepressor protein complexes interacting with the TR with or without bound T3 so that transcription may be initiated or suppressed (1 –3). The system requires cellular uptake and transport into the nucleus of T3.
When we sought to determine whether the cycling of integrin αvβ3 to and from the plasma membrane might be regulated by thyroid hormone, we found that T4-treated tumor cells did internalize the integrin (16). Internalization of the heterodimeric integrin, however, did not include transport of the ligand into the cytoplasm. The αv monomer surprisingly was imported into the nucleus in hormone-exposed cells, whereas the β3 monomer remained in the cytoplasm. In the nucleus, monomeric αv was also found to be phosphorylated and complexed with activated MAPK (pERK1/2) (16). Monomeric αv was not subject to phosphorylation by MAPK in vitro or in cells in which there was β3 knockdown. This suggests that internalization of the heterodimer is required in order to obtain monomer phosphorylation.
After immunoprecipitation, the αv monomer–MAPK complex was found to include p300 and STAT1α. These nuclear coactivator proteins have been associated with genomic actions of thyroid hormone that begin with ligand binding of T3 by TRβ1. The coactivators are subject to phosphorylation. In our studies of T4-treated cells, the monomeric αv–MAPK–p300 complex in the nucleus bound to the promoter region of several genes, including, as expected, TRβ1. Quantitative polymerase chain reaction confirmed increased transcription of this gene (16). The αv–MAPK–p300 complex was shown to bind to the promoters of several other genes, including ERα, and to be result in increased expression of these genes. It was also apparent in these studies that the αv–MAPK could transfer to the nucleus as a complex that contained two corepressors, as well as coactivators. This raised the possibility that nuclear αv–MAPK organizes these transcription-modifying nucleoproteins that are subject to phosphorylation. From the standpoint of this review, however, the conclusion is that T4 can act at the intact integrin on the cell surface as a hormone rather than a prohormone, and regulate the availability of nuclear TR.
Phosphorylation and acetylation of nuclear TR
In studies conducted before the specific receptor for thyroid hormone on integrin αvβ3 was described (6), it was apparent that tetrac blocked rapid-onset nongenomic actions of thyroid hormone in membranes (8,32) and that agarose–T4—a formulation excluded from the cell interior—initiated effects that terminated in the cell nucleus (10). Using these probes of plasma membrane–initiated effects of the hormone, we showed that T4 caused complexing of TRβ1 and MAPK in the nucleus of noncancer cells and serine phosphorylation of TR within 10–20 minutes (10). The serine likely to be phosphorylated was identified, as was the docking site on TRβ1 for MAPK (11). Dissociation of TR and the corepressor, silencing mediator of retinoid and thyroid hormone, resulted from serine phosphorylation, a step required to relieve silencing and to permit consequent activation of the receptor (10).
TRβ1 is also subject to T4-directed acetylation (12). Interestingly, this modification of the receptor requires MAPK activation, but not serine phosphorylation of TR, itself. This is consistent with an integrin-mediated mechanism involving MAPK activation from the S2 domain of the TR on integrin αvβ3, as described above. Acetylation is carried out by steroid receptor coactivator-1, a coactivator of nuclear steroid and TRs, and the acetylation step is associated with recruitment of coactivator p300 by TRβ1 (12). Tetrac blocks the acetylation process initiated by T4.
Thus, phosphorylation and acetylation of TR are modifications of the receptor that alter receptor function. Both are rapid-onset consequences of T4 action that prepare TR for activation (phosphorylation) or are a part of activation (acetylation) with recruitment of p300. The susceptibility of these hormone-directed changes in the state of TR to inhibition by tetrac or reproduction with agarose-T4 that acts exclusively at the cell membrane is consistent with involvement of αvβ3 in the hormone action. However, these processes have not yet been subjected to a second probe of inhibition of actions originating at the receptor on the integrin, namely, the addition of anti-αvβ3.
Discussion
The presence of a plasma membrane receptor for thyroid hormone on integrin αvβ3 provides a molecular basis for the distinction between genomic and nongenomic actions of iodothyronines. At the same time, recognition of effects at the integrin of T4 as a hormone, rather than as a prohormone source of intracellular T3, integrates nongenomic and genomic mechanisms of hormone action, contributing to the function of the genomic nuclear TR–T3 complex. Via αvβ3, T4 supports (i) nuclear uptake of TRβ1 from the cytoplasm and uptake of activated MAPK in a complex with TR, (ii) phosphorylation and acetylation of TR that are important to coactivator–corepressor recruitment, (iii) assembly of intranuclear complexes via monomeric αv of coactivators or corepressors that are relevant to transcriptional activity of TRβ1, and (iv) transcription of the TRβ1 gene.
Integrin αvβ3 interacts with a large number of extracellular matrix (ECM) proteins (33 –35), with growth factors and with adjacent growth factor receptors on the cell surface (36 –38). These interactions are transduced into complex cellular responses, including motility and adhesion and cell division and into effects on angiogenesis (12,36,39). The responses involve integrated transcription of genes. The integrin has relatively recently been appreciated to contain specific receptor sites for small molecules (40,41), in addition to that for thyroid hormone (6,42). The capacity of the integrin to modulate the transcription of a number of genes in response to microenvironmental ECM inputs via downstream kinases is a context in which to understand the complex actions of thyroid hormone that are initiated at αvβ3 (23 –27). In addition, the modulation of transcription of several genes, including TRβ1, ERα, and COX-2, may be a function of internalization of integrin αv monomer in thyroid hormone–exposed cells (16). Studies that disclosed this previously unrecognized function of αv were prompted by reports that internalized αv may be homogeneously distributed throughout the cytoplasm (43,44), rather than localized in endosomes from which heterodimeric αvβ3 may be recycled to the plasma membrane. These reports suggested the existence of additional functions for the monomer. Interestingly, intensity of the diffuse intracellular monomer signal was increased in metastatic lung cancer cells, compared with cells from primary tumors (44).
The affinity of the iodothyronine receptor on integrin αvβ3 for T4 is high (K d=3×10−10 M) (6) and compares favorably with affinities for T3 of nuclear receptors described in several human (45 –47) and animal (48 –50) tissues. The affinity of the TR on αvβ3 for T3 is lower than that for T4 (6), whereas the avidity of nuclear hormone receptors for T4 is 10–100-fold lower than that for T3 (45,47). Thus, it is unlikely that the functions of T4 that have been described to start at αvβ3 reflect conversion of T4 to T3 and actions of T3 within the cell, for example, in the nucleus. Further, T4 covalently bound to agarose—a nanoparticulate formulation of T4—does not gain access to the cell interior, but reproduces a number of nongenomic actions of unmodified hormone known now to be mediated by integrin αvβ3 (20,32,51 –53). These factors are consistent with function of T4 at the integrin as a hormone, rather than as a prohormonal source of T3. In addition, actions of thyroid hormone initiated at αvβ3 that we have described in vitro have been confirmed to occur when conversion of T4 to T3 is inhibited by propylthiouracil (10,52). Human platelets respond to T4 via αvβ3 with agglutination, but there is no response to T3 (53), and the ability of the S1- and S2-binding domains of the hormone receptor on αvβ3 to distinguish among thyroid hormone analogs has been described (9).
The possibility must be considered that hormone uptake systems (transporters) in the plasma membrane compete with the TR on αvβ3 for iodothyronines. The existence of such competition would serve to reduce the actions of thyroid hormone at the integrin. That actions of agarose-T4 (51 –53) or agarose-T3 (32) reproduce actions of unmodified thyroid hormone suggests that such competition, if it does exist, is modest. Indeed, nanoparticulate tetrac cannot enter the cell, but is somewhat more potent than unmodified tetrac (54). The actions initiated at αvβ3 by thyroid hormone are rapid in onset in cultured cells (55), indicating that receptor site occupancy of thyroid hormone on the integrin need not be prolonged in order to cause a response.
Is it possible that the integrin is a transporter? No such function has been attributed to αvβ3 or other integrins. Integrin αvβ3 is a structural protein of the plasma membrane, primarily responsible for interpreting the presence of specific ECM proteins in the immediate cellular microenvironment and for transducing ECM protein–integrin interactions into intracellular signals at the cytoskeleton and in the nucleus (56 –58). Other features of integrin αvβ3 also make it unlikely that the protein functions as a hormone transporter. For example, (i) the binding of T4 and T3 by the integrin can modify signaling between αvβ3 and nearby vascular growth factor receptors on the cell surface, so that angiogenesis is stimulated (8,36). (ii) Acting at the integrin, T4 also stimulates fibroblast and granulocyte migration toward a chemical cue (S.A. Mousa, unpublished observations). (iii) The expression of specific genes not previously reported to be susceptible to thyroid hormone or hormone analogs such as tetrac via nuclear TR can be modulated from αvβ3 (28,30). (iv) Integrin αvβ3 is internalized and recycled, but does not bind thyroid hormone within the cell (16). (v) Within thyroid hormone–treated cells, the imported αv monomer (16) is widely distributed (43,44), is taken up by the nucleus, and becomes a coactivator (16). Coactivator proteins may reside in the cytoplasm (59,60) and be imported by the nucleus as needed, but integrin αv appears to be the first integrin recognized to be distributed periodically throughout the cell and to be subject to involvement in coactivator complex formation in the nucleus.
The relatively stable concentration of free T4 at the cell surface in the intact organism implies that actions of the hormone at αvβ3 contribute to the setting of rates of basal activities of processes controlled from the integrin, including plasma membrane ion transport (61) and certain genomic functions reviewed here. From the standpoint of critical genomic actions of T3 within the nucleus, it is obvious that stable availability of TR—via trafficking and receptor gene expression at least in part regulated from the integrin—is desirable.
Footnotes
Acknowledgments
The authors acknowledge the generous support from M. Frank and Margaret D. Rudy (Los Angeles) and from Richard Liebich (Albany) who supported a number of studies reviewed here.
Author Disclosure Statement
No competing financial interests exist.