
Abstract
Background:
Progesterone (P4) is the main steroid secreted by the corpora lutea (CL) and is required for successful implantation and maintenance of pregnancy. Although adequate circulating levels of thyroid hormone (TH) are needed to support formation and maintenance of CL during pregnancy, TH signaling had not been described in this gland. We determined luteal thyroid hormone receptor isoforms (TR) expression and regulation throughout pregnancy and under the influence of thyroid status, and in vitro effects of triiodothyronine (T3) exposure on luteal P4 synthesis.
Methods:
Euthyroid female Wistar rats were sacrificed by decapitation on gestational day (G) 5, G10, G15, G19, or G21 of pregnancy or on day 2 postpartum (L2). Hyperthyroidism and hypothyroidism were induced in female Wistar rats by daily administration of thyroxine (T4; 0.25 mg/kg subcutaneously) or 6-propyl-2-thiouracil (PTU; 0.1 g/L in drinking water), respectively. Luteal TR expression of mRNA was determined using real-time reverse-transcription quantitative polymerase chain reaction, and of protein using Western blot and immunohistochemistry. Primary cultures of luteal cells and of luteinized granulosa cells were used to study in vitro effects of T3 on P4 synthesis. In addition, the effect of T3 on P4 synthesis under basal conditions and under stimulation with luteinizing hormone (LH), prolactin (PRL), and prostaglandin E2 (PGE2) was evaluated.
Results:
TRα1, TRα2, and TRβ1 mRNA were present in CL, increasing during the first half and decreasing during the second half of pregnancy. At the protein level, TRβ1 was abundantly expressed during gestation reaching a peak at G19 and decreasing afterwards. TRα1 was barely expressed during early gestation, peaked at G19, and diminished thereafter. Expression of TRβ1 and TRα1 at the protein and mRNA level were not influenced by thyroid status. T3 neither modified P4 secretion from CL of pregnancy nor its synthesis in luteinized granulosa cells in culture.
Conclusions:
This study confirms for the first time the presence of TR isoforms in the CL during pregnancy and postpartum, identifying this gland as a TH target during gestation. TR expression is modulated in this tissue in accordance with the regulation of P4 metabolism, and the abrupt peripartum changes suggest a role of TH during luteolysis. However, TH actions on the CL do not seem to be related to a direct regulation of P4 synthesis.
Introduction
T
Several endocrinologic abnormalities such as thyroid disease have been implicated as etiologic factors of recurrent pregnancy loss (3 –9). Hypothyroidism has a statistically significant relationship with recurrent pregnancy loss in the first trimester in humans (10). Hypothyroidism also interferes with the formation and function of CL, resulting in pregnancy failure. Experimental findings indicate that the presence of an adequate circulating level of thyroid hormone (TH) is needed to support CL formation and pregnancy (5,6,11).
Previous results from our laboratory demonstrated that hypothyroidism impairs CL function during gestation delaying the onset of parturition in the rat (12,13). On the contrary, hyperthyroidism advances P4 withdrawal and the onset of parturition (14,15). The effects of hyperthyroidism or hypothyroidism on luteal function are mediated by changes in prostaglandin balance. At the luteal level, hyperthyroidism decreased prostaglandin E2 (PGE2) and increased prostaglandin PGF2a (PGF2a), while hypothyroidism increased PGE2 and diminished PGF2a at the end of gestation. Collectively, these alterations result in changes in the moment of induction of 20α-hydroxysteroid dehydrogenase (20α-HSD) expression, the enzyme that metabolizes P4 to an inactive metabolite. Thus, in hyperthyroid rats the expression of this enzyme is advanced and in hypothyroid rats it is delayed, resulting in advanced and delayed parturition, respectively (13,14). These data may indicate that THs participate in the maintenance and function of the CL of gestation in rodents.
THs play critical roles in differentiation, growth, and metabolism. The classic genomic actions of THs are mediated by two main nuclear receptor isoforms, TRα and TRβ, that act mainly as hormone-inducible transcription factors. The isoforms of TRα and TRβ are encoded by the THRA and THRB genes. The TRα1, TRα2, and TRβ1 isoforms are widely expressed whereas TRβ2 is predominantly restricted to the hypothalamus and pituitary. TRα1, TRα2, and TRβ1 mRNA and protein are present in ovarian surface epithelial cells in humans (16), whereas human oocyte, granulosa, cumulus, and stromal cells express TRα1, TRα2, TRβ1, and TRβ2 (17 –19). Binding studies showed that triiodothyronine (T3) binds to CL nuclei in a hormone-specific manner indicating a direct effect of THs on human CL (20). Another investigation demonstrated that T3 stimulates P4 release from human luteal cells in an indirect manner, mediated by a putative proteinaceous factor (21). However, the precise role of TR signaling in the CL of gestation remains elusive.
Although the CL of pregnancy is not an established TH target, the evidence reported indicates that TH signaling plays a role on luteal function during pregnancy, thus becoming an issue for further investigation. Therefore, we hypothesized that THs signal in the CL through their known receptors in a canonical manner allowing the normal function of the CL. In this sense, we set out to determine: (1) which TR isoforms are expressed in CL; (2) whether this family of proteins is differentially expressed throughout pregnancy; (3) whether there is differential expression of the TRs at the mRNA or protein level; (4) and if they are regulated by thyroid status. Finally, because P4 is the main product of CL, we also sought to determine whether exposure of T3 might affect luteal P4 synthesis in vitro.
Materials and Methods
Animals and experimental design
Adult female Wistar rats bred in our laboratory, 3–4 months old, weighing 200–230 g at the onset of treatment and with regular 4-day estrus cycles were used. The estrus cycle was monitored by cytologic examination of the vaginal smears obtained by daily vaginal lavage during at least three consecutive estrus cycles before starting the treatments. They were kept in a light- (lights on 6:00
To determine the pattern of hormonal secretion and TR expression in the CL during pregnancy, groups of 6–8 euthyroid rats were sacrificed by decapitation within 5 seconds after removal from the home cage on G5, G10, G15, G19, or G21 or on day 2 (L2) postpartum between 10:00
Animal maintenance and handling was performed according to the Guiding Principles in the Care and Use of Laboratory Animals of the U.S. National Institutes of Health (NIH publication no. 86-23, revised 1985 and 1991) and the United Kingdom's requirements for ethics of animal experimentation (Animals Scientific Procedures, Act 1986). All procedures were approved by the Institutional Animal Care and Use Committee of the School of Medical Science, National University of Cuyo, Mendoza, Argentina (Protocol approval no. 17/2012).
Hormone determinations
Prolactin (PRL) and thyrotropin (TSH) were measured by double-antibody radioimmunoassay (RIA) using materials generously provided by Dr. Parlow and the National Hormone and Pituitary Program, Harbor-UCLA Medical Center, Torrance, CA, as previously described (12).
P4, T3, and T4 concentrations in sera were measured by radioimmunoassay using commercial kits for total hormones (DSL-3400, DSL-3100, and DSL-3200 double-antibody radioimmunoassay, respectively, from Diagnostic System Laboratories (Webster, TX) as previously described (11).
RNA isolation and CL analysis
Reverse-transcription quantitative polymerase chain reaction (RT-qPCR) analysis was used to detect the presence of TR isoforms in CL of pregnancy. Total RNA was extracted from 100 mg of deep-frozen CL, using TRIzol® reagent (Life Technologies, Carlsbad, CA), and following the instructions provided by the manufacturer. The integrity of the total RNA was determined by electrophoresis on 2% (w/v) agarose gels stained with ethidium bromide. Nucleic acid concentrations were measured at 260 nm. Purity of the total RNA extracted was determined as the 260:280 nm ratio with expected values between 1.8 and 2.
Five micrograms of total RNA were reverse-transcribed to cDNA in a 25 μL volume reaction in the PTC-100 Programmable Thermal Controller (MJ Research Inc., Bruno, Canada). The RNA was first denatured at 70°C for 5 minutes in the presence of 2.5 μg of random hexamer primers (Invitrogen Life Technologies, Carlsbad, CA). For the subsequent RT reaction the following mixture was added: RT buffer (50 mM Tris-HCl [pH 8.4], 75 mM KCl, 3 mM MgCl2), 0.5 mM dNTPs, 5 mM DTT, 200 units M-MLV Reverse Transcriptase (Invitrogen Life Technologies). The reaction was incubated at 37°C for 50 minutes and then inactivated by heating at 70°C during 15 minutes. The cDNA was stored at −20°C. PCRs were performed using a Corbett Rotor Gene 6000 Real-Time Thermocycler (Corbett Research Pty Ltd. Sydney, Australia) using Eva-Green (Biotium, Inc., Hayward, CA) in a final volume of 20 μL. The reaction mixture consisted of 2 μL of 10× PCR buffer, 1 μL of 50 mM MgCl2, 0.4 μL of 10 mM dNTP Mix (Invitrogen Life Technologies), 1 μL of 20× Eva Green (Biotium, Inc., Hayward, CA), 0.25 μL of 5 U/μL Taq DNA Polymerase (Invitrogen Life Technologies), 0.1 μL of each 2.5 mM primer (forward and reverse primers) and 10 μL of diluted cDNA. The PCR reactions were initiated with 5-minute incubation at 95°C, followed by 40 cycles of 95°C for 30 seconds, 60°C for 30 seconds, and 72°C for 30 seconds. A melting curve was performed at the end of the PCR run over the range of 55–95°C, increasing the temperature stepwise by 0.5°C every 2 seconds. Gene-specific amplification was confirmed by a single peak in the melting-curve analysis and a single band on a 2% agarose gel stained with ethidium bromide. All pairs of primers listed in Supplementary Table S1 (Supplementary Data are available online at
All qPCR reactions were performed in duplicate and Cq values were averaged; every run also included a nontemplate control and a control for cDNA contamination that contained a pool of RNA of the samples and was subjected to the protocol of retro-transcription exchanging the retrotranscriptase for water in the reaction mixture. All qPCR assays were validated in silico using the RTPrimerDB primer and BLAST evaluation, checking primer specificity. The data acquisition of the expression levels was done employing the second derivative maximum method as computed by the Rotor Gene 6000 Rotary Analyzer Software version 1.7.87 (Corbett Research Pty Ltd., Sydney, Australia). For subsequent data analysis, the values provided by the instrument were imported into a Microsoft Excel spreadsheet and analyzed using the 2−ΔΔCq method (26). Fold change in normalized mRNA expression of the different genes of interest, relative to the expression at early gestation (G5), was calculated for each sample throughout gestation, and relative to the control group when assessing the difference among thyroid status.
A dilution series was created from a pool of cDNA made from aliquots of all of our samples in order to construct standard curves for each primer pair. In brief, qPCR reactions were performed in duplicate for each serial dilution and were plotted against the logarithm of the cDNA dilution factor. The amplification efficiency for each gene evaluated was calculated from the expression [10(1/−S) − 1], in which S represents the slope of the linear regression obtained.
The expression stability of the four candidate reference genes, Gapdh, Rpl19, Hprt1, and Rps16 were estimated using the freely available online software BestKeeper version 1 (
Western blot analysis of TH receptors
TRα1 and TRβ1, the predominant TH binding receptors, were analyzed by Western blot at the luteal level in order to determined their protein expression and regulation during pregnancy. TR proteins were expressed as the ratio of signal intensity for the protein relative to that of α-tubulin. Protein samples used for the detection of TRs during gestation were isolated from the phenol-ethanol supernatant obtained from RNA isolation using TRIzol reagent following the manufacturer's instructions.
Protein samples used for the detection of TR on G19 in hypothyroid, hyperthyroid, and control rats were isolated from CL homogenized in 10 volumes of lysis buffer that contains 50 mM Tris-HCl (pH 7.5), 150 mM NaCl, 0.5% IGEPAL, 50 mM sodium fluoride, and a protease inhibitor cocktail (Sigma P8340, Sigma-Aldrich, St. Louis, MO). The lysates were incubated on ice for 30 minutes and then another 30 minutes on ice on a rocking platform. Lysates were centrifuged at 10,000 rpm for 15 minutes at 4°C, and the supernatant was considered whole cell extract. Protein content was assessed by the bicinchoninic acid method (BCA; Pierce, Rockford, IL). The whole cell extract was aliquoted and stored at −80°C. Before loading, the proteins were boiled for 10 minutes in sample buffer and an amount of 50 μg of proteins were separated in a 12% (w/v) acrylamide gel by sodium dodecyl-polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to polyvinylidene difluoride (PVDF) membranes. The blots were blocked in 5% (w/v) nonfat milk in tris-buffered saline (TBS) containing 0.1% (v/v) Tween-20. Blots were probed overnight with the appropriate dilution in 2.5% bovine serum albumin (BSA) of each of the primary antibodies. The membranes were washed 3×5 minutes in TBS-T and incubated with 1:5000 dilution of a peroxidase conjugate secondary antibody (anti-mouse horseradish peroxidase [HRP] from Cell Signaling [Beverly, MA] and anti-rabbit HRP from Santa Cruz Biotechnology [Dallas, TX]) for 1 hour at room temperature. The blots were washed, developed by chemiluminiscence, using a ChemiDoc XRS+ System (Bio-Rad Laboratories, Hercules, CA). Densitometric analysis of the protein bands was performed using Image Lab Software version 4.0 from Bio-Rad Laboratories expressing the results as the ratio of signal intensity for the different TR proteins relative to that of α-tubulin. The following dilutions were used for the primary antibodies: TRα/β (FL-408; 1/1000), TRβ1 (J52; 1/1000) from Santa Cruz; TRα1 (PA1-211A; 1/1000) from Thermo Scientific (Waltham, MA), and α-tubulin (T6040; 1/12000) from Sigma-Aldrich.
Immunohistochemistry
To determine the cellular localization of the TRs, we performed immunohistochemistry (IHC) analysis in paraffin sections of whole ovaries. The tissues were fixed in 10% buffered formalin at room temperature, and processed for paraffin embedding. Five-micrometer sections were cut in a Reichert-Jung Hn 40 microtome and mounted onto 3-aminopropyltriethoxysilane-coated slides (Sigma-Aldrich) for subsequent IHC analysis using the mouse monoclonal antibody TRβ1 (J52).We performed IHC in these samples using the antibody sc-772 that recognizes both TRα1 and TRβ1 isoforms, and PA1-211A that recognizes specifically TRα1, but we did not observe a reliable signal with any of these antibodies (data not shown).
TR antigen unmasking was carried out in 0.01 M citrate buffer (pH 6.0) at 100°C for 25 minutes. The slides were incubated with the primary antibody overnight at 4°C in a humidity chamber at 1/200 dilution. Nonspecific mouse immunoglobulin G1 (IgG1) antibody and purified rabbit preimmune serum (Dako, Kingsgrove, NSW, Australia) were used as isotype controls. Secondary biotin-conjugated goat anti-mouse/rabbit antibodies (Vector Laboratories Homebush, NSW, Australia) were used at 1/250 dilution. Specific primary-secondary antibody complexes were detected using ABC reagent (Vector) and visualized using a DAB peroxidase substrate kit (Vector). Slides were lightly counterstained with hematoxylin to reveal nuclei and observed with a Nikon Eclipse E200 microscope (Nikon Instruments Inc., Melville, NY) and processed with Micrometrics SE Premium Software (Accu-Scope Inc., Commack, NY).
Primary luteinized granulosa cells and luteal cell culture
To determine a possible direct effect of THs on luteal cell P4 synthesis, primary luteinized granulosa cells were cultured as previously described (27). Briefly, 25-day-old immature rats were treated with 10 IU pregnant mare serum gonadotropin (PMSG) intraperitoneally followed by 10 IU human chorionic gonadotropin (hCG) intraperitoneally 2 days later. Eight hours after hCG injection, follicles were gently extruded from the ovaries under a dissecting microscope, incubated sequentially in DMEM/F12 (1:1) containing, respectively, 6 mM ethylenediaminetetraacetic acid (EDTA) and then in 0.5 M sucrose, and luteinized granulosa cells were harvested by needle-pricking the follicles. The cells were cultured for 48 hours at 37°C in an atmosphere of 5% CO2/95% air in DMEM/ F12 (1:1) with 15 mM HEPES, 1% fetal bovine serum (FBS), and 100 IU/mL penicillin G, 100 μg/mL streptomycin, and 0.25 mg/mL amphotericin B. The medium was then replaced and cells were treated for 48 hours with T3 (100 nM) using PRL (1 μg/mL), PGE2 (1 μM), and luteinizing hormone (LH; 100 ng/mL) as stimulators. Each experimental condition was performed in triplicate and the experiment was repeated twice. The concentrations used for T3, PRL, PGE2, and LH were chosen based on previous published data (28 –30). At the end of the experiment, cells were centrifuged, and the culture medium was frozen at −20°C for subsequent RIA of P4.
For primary luteal cell culture, we used the method described by Nelson et al. (29). Briefly, on day 19 of pregnancy, three control or hypothyroid rats were killed by decapitation, ovaries were removed, and CL dissected. The CLs were placed in Hanks balanced salt solution (HBSS) media containing 2% bovine serum albumin (BSA) and 25 mM HEPES, pH 7.4. Any follicle adhering to the CL was carefully removed. CL were incubated at 37°C with 50 U/mL collagenase, 2.4 U/mL dispase, and 200 U/mL DNase in four consecutive incubations (30 minutes each), with stirring at 100 rpm under an atmosphere of 100% O2. After incubation, the cells were centrifuged at 200g, the supernatant was discarded, and fresh medium and enzymes were added. At the end of enzyme treatment, CL were treated for 15 minutes in 10 mL EDTA solution (0.02% EDTA w:v in phosphate buffered saline [PBS]) containing 2% BSA and 25 mM HEPES, pH 7.4. After this treatment, the cells were centrifuged at 200g. The new pellet was resuspended in 10 mL of dissection medium and filtered through nylon mesh. The viability was between 80% and 90% as determined by trypan blue staining. For all experiments, luteal cells were cultured at 37°C under 95% air: 5% CO2 atmosphere, using 105 viable cells per milliliter of culture medium (McCoy's 5A:Ham's F-12, 1:1). The cells were incubated for 6 hours with T3 (1–500 nM) using pregnenolone (100 nM) as precursor for P4 synthesis. At the end of each experiment, the cells were centrifuged, and the culture medium was frozen at −20°C for subsequent RIA of P4.
Statistical analysis
Statistical analysis was performed using one-way analysis of variance followed by Bonferroni post hoc test to compare between groups. When variances were not homogeneous, we performed log transformation of the data. In the cases in which the data do not present a Gaussian distribution after log transformation, the non-parametric Kruskal Wallis test, followed by Dunn post hoc test, was used to compare the groups. Differences between means were considered significant at the p<0.05 level (31).
Results
Hormonal profile of T3, T4, TSH, and P4 levels during gestation and postpartum
The present results (Fig. 1) confirm (12) that the thyroid-pituitary axis is regulated during gestation and postpartum in the rat. T4 decreased significantly throughout gestation, and returned after parturition to levels similar to those found in early pregnancy (G5). Conversely, the levels of T3 increased slightly on G15 and decreased thereafter, remaining at levels similar to those of early pregnancy (G5). TSH surged on G15 and decreased thereafter, but it remained at higher levels than during the first half of gestation. P4 concentration was elevated throughout gestation and decreased on G21 due to the triggering of luteolysis concomitant with the onset of parturition. P4 remained at low levels in the early postpartum period.
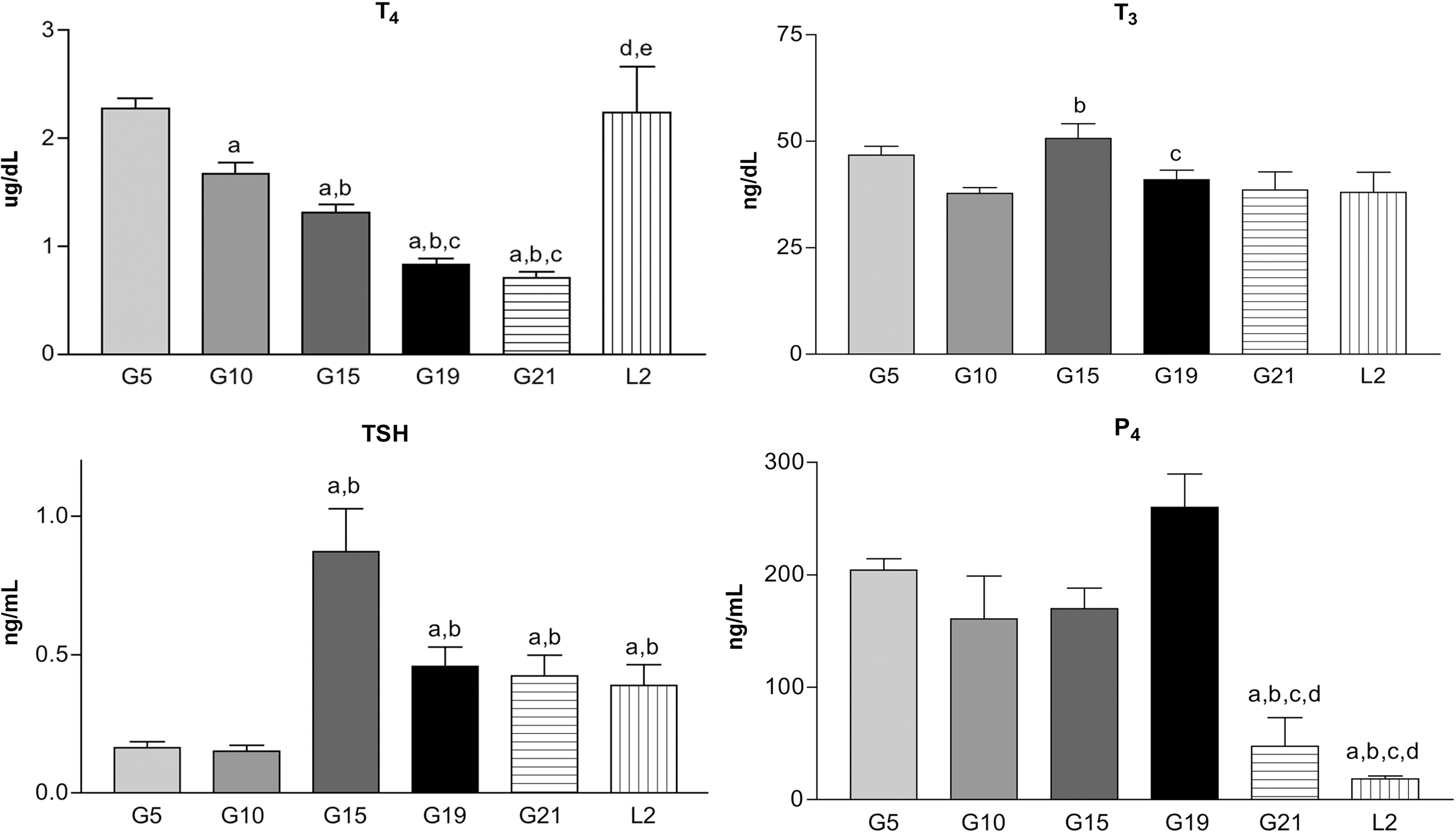
T4, T3, TSH, and P4 circulating concentrations during gestation and postpartum in the rat. Serum concentration of T4, T3, TSH, and P4 assessed by RIA on G5 (n=6), G10 (n=6), G15 (n=6), G19 (n=6), G21 (n=9), and L2 (n=6). Results are shown as means±SEM of groups of 6–9 rats. To compare T3 and TSH after log transformation, one-way ANOVA and Bonferroni posttest was used to compare between groups. To compare P4 and T4, Kruskal Wallis and Dunn posttest was used to compare between groups. Statistical significance (p<0.05) is indicated: avs. G5, bvs. G10, cvs. G15, dvs. G19, and evs. G21. T4, thyroxine; T3, triiodothyronine; TSH, thyrotropin; P4, progesterone; RIA, radioimmunoassay; G#, gestational day #; L2, day 2 of lactation; SEM, standard error of the mean; ANOVA, analysis of variance.
TRs mRNA expression at luteal levels during gestation and postpartum
In order to evaluate for the first time a possible direct action of THs on CL function during pregnancy, the expression of thyroid hormone receptors was assessed. Figure 2 shows that in the rat, the CL of pregnancy express TRα1, TRα2, and TRβ1 mRNA, however, the presence of TRβ2 was not detectable (data not shown). TRα1 mRNA expression increased during the first two-thirds of gestation reaching a peak at G15, and decreased significantly at G19, remaining at levels similar to those of G5 at the end of gestation and after parturition. TRα2 mRNA expression was similar on all the days of pregnancy studied, with the exception of a peak observed on G10, although the differences were only significant compared with G19 and G21. TRβ1 mRNA expression tended to increase in the first part of gestation; on G19 the expression decreased significantly compared with G10, and it increased gradually thereafter reaching significant differences in the postpartum period with respect to G19.
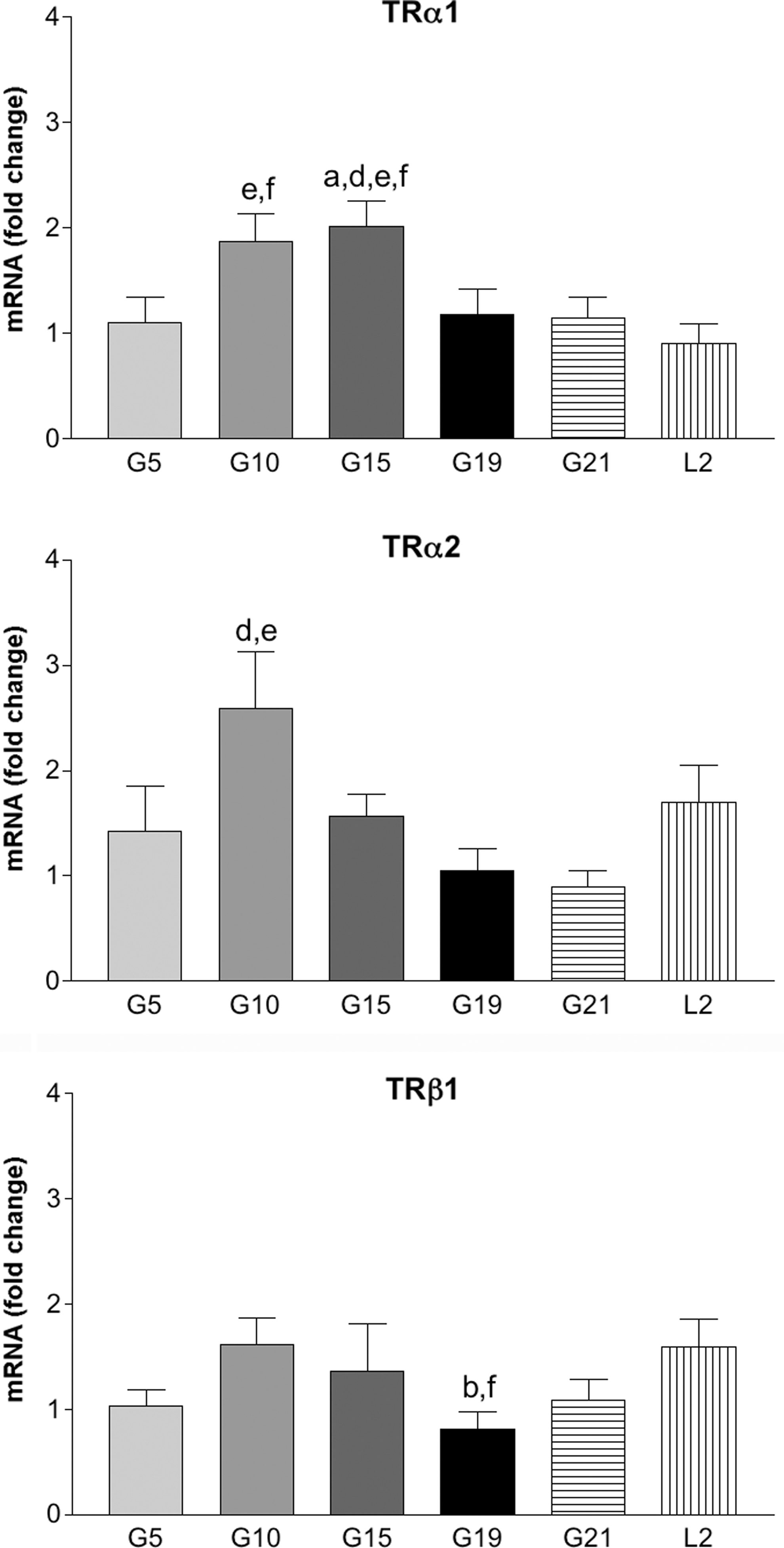
TRα1, TRα2, and TRβ1 luteal mRNA expression. mRNA fold change with respect to G5 and normalized to gapdh estimated by real-time RT-PCR during gestation and postpartum in the rat. The results are shown as means±SEM of groups of 5 rats. One-way ANOVA and Bonferroni posttest were used to compare between groups to estimate TRβ1 expression, and nonparametric Kruskal Wallis and Dunn posttest were used to compare between groups for TRα1 and TRβ2. Statistical significance (p<0.05) is indicated: avs. G5, bvs. G10, cvs. G15, dvs. G19, and evs. G21. RT-PCR, reverse transcription-polymerase chain reaction.
TRα1 and TRβ1 protein expression in CL during gestation and postpartum and its regulation by thyroid status in the rat
The two most widely expressed and best characterized isoforms that bind T3 are TRα1 and TRβ1. Because the normal expression of these proteins has never been identified in the CL, we decided to determine their luteal expression at protein level during gestation and postpartum. We identified the presence of TRβ1, which is abundantly expressed during gestation and postpartum in CL (Fig. 3). The expression of this receptor slightly increased during gestation reaching a peak on G19 and decreasing significantly on the last day of gestation and postpartum. On the contrary, TRα1 is barely expressed at the beginning of gestation and increased significantly with respect to the expression on G19 as well as compared to TRβ1; its expression diminished thereafter (Fig. 3).
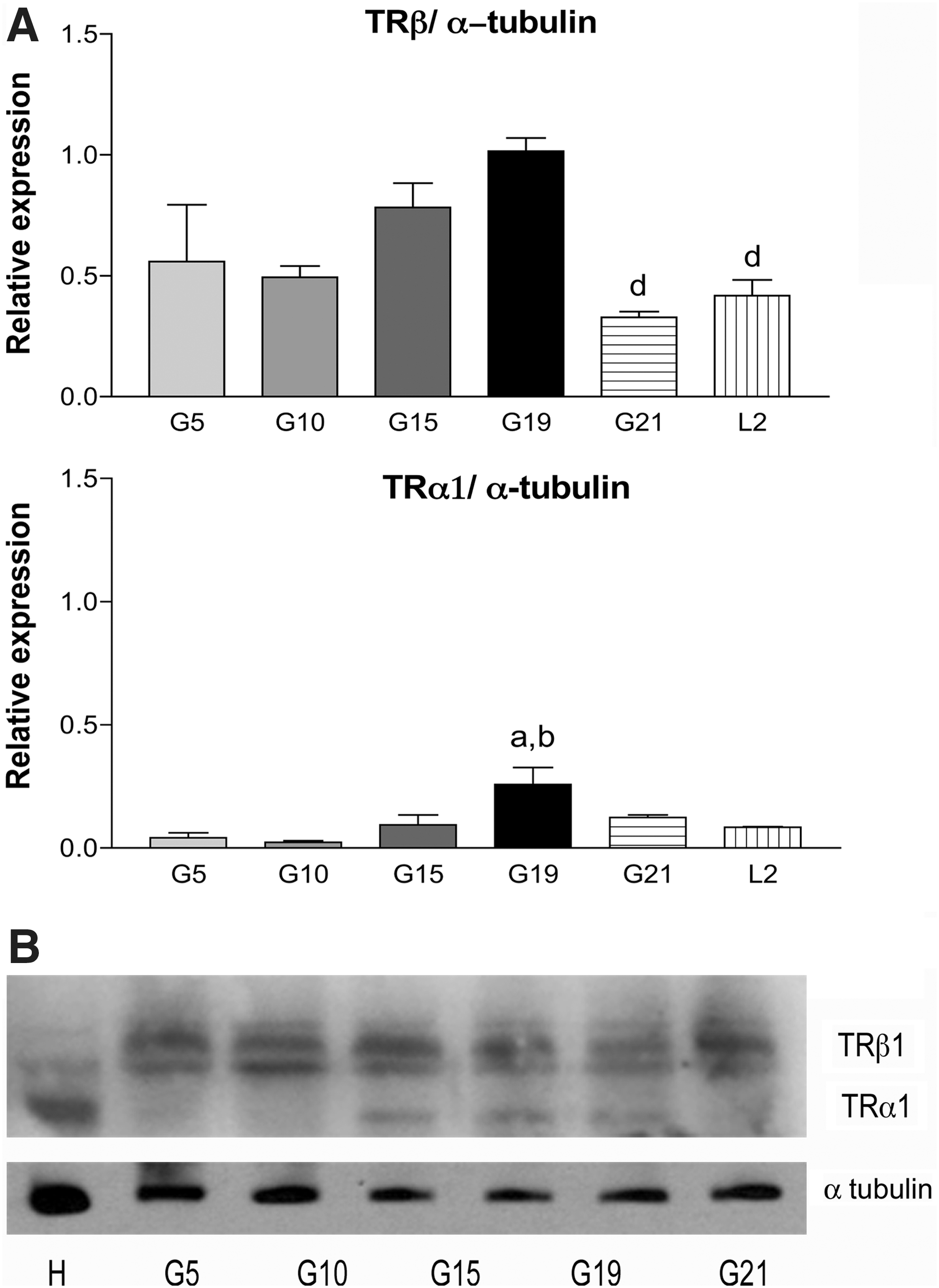
Luteal TR protein expression throughout gestation in the rat.
Due to the increased expression of both TR isoforms found on G19 we explored the influence of thyroid status on their expression at luteal level on this day. To further corroborate the specificity of the signals, we used specific antibodies for TRβ1 (J-52) and TRα1 (PAI-211) that confirmed the results shown in Figure 3 and demonstrated that CL express TRβ1 and TRα1 (Fig. 4). To determine if thyroid status can modulate the expression of TRs at luteal level, as has been shown for other rat tissues such as myocardium and epididymis (22,32), we determined the expression of TRβ1 and TRα1 at the protein and mRNA level at G19 in control, hypothyroid and hyperthyroid rats. Our results show that thyroid status does not affect the expression of luteal TR at this stage of pregnancy since the expression of TRβ1 and TRα1 at the protein (Fig. 4) and mRNA level (Supplementary Fig. S1) remained unchanged. Hyperthyroid and hypothyroid status were confirmed measuring circulating T4 and TSH. Hyperthyroid rats had significantly increased T4 and decreased TSH, while hypothyroid rats had decreased T4 and increased TSH serum concentrations compared to the control group (Supplementary Fig. S2).

Luteal TR protein expression at the end of pregnancy in euthyroid, hypothyroid, and hyperthyroid rats.
Immunolocalization of TRs in CL of gestation in hypothyroid, hyperthyroid, and control rats
In order to confirm the presence of the receptor and to evaluate its cellular localization we determined the presence of TRα1 in the CL of gestation by IHC (Fig. 5). The immunostaining signal observed was moderate in CL and intense in granulosa cells of follicles within the ovary. The immunolocalization was predominantly nuclear as expected. When comparing the presence and localization of the mentioned isoform in CL of gestation in hypothyroid, hyperthyroid and control rats, we did not observe any significant difference due to thyroid status. Although we performed the experiments evaluating the immunolocalization of TRα1 and α2, the antibodies tested did not give a reliable signal and were not considered for this study.
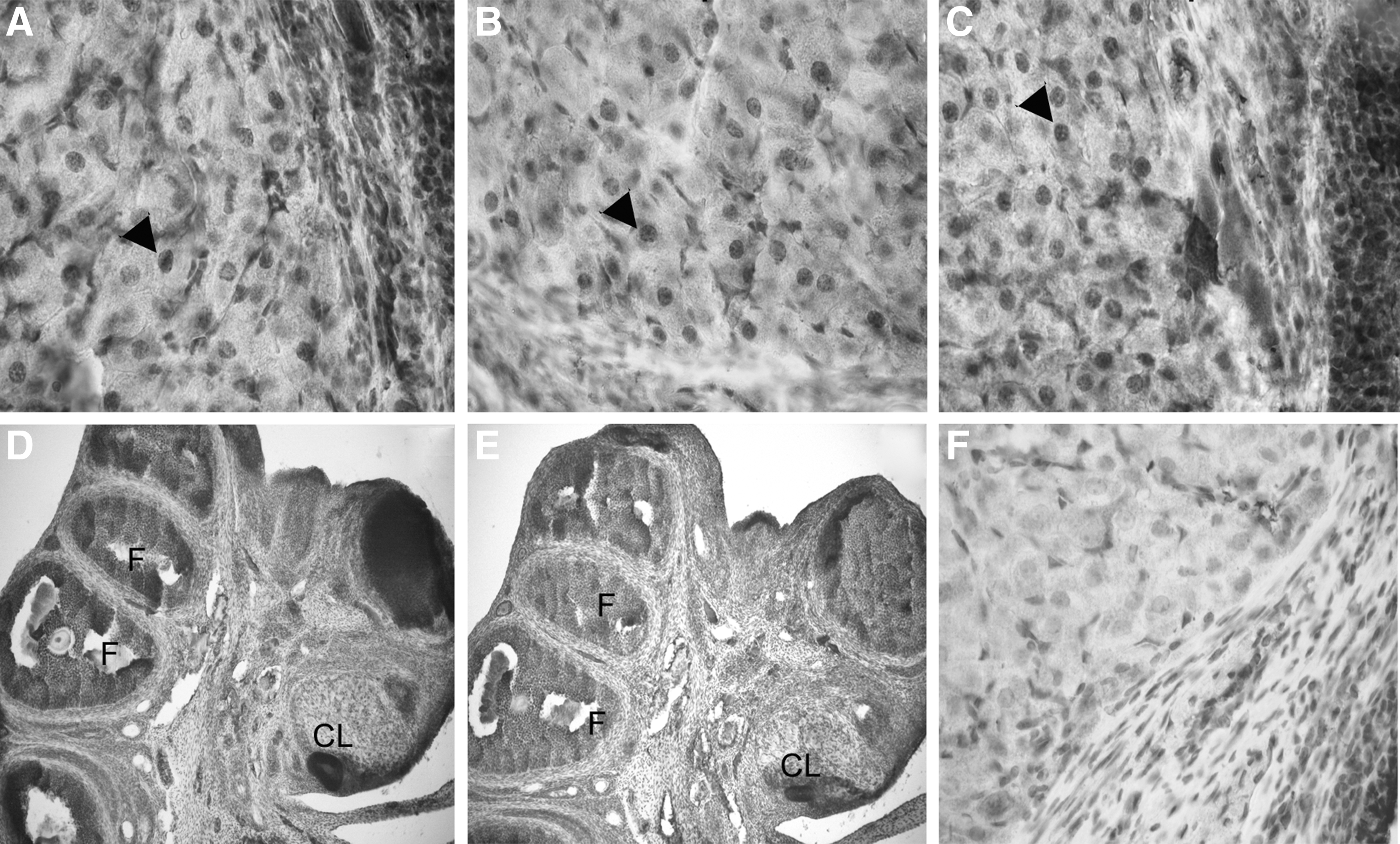
Ovarian localization of TRβ1 on day 19 of gestation in euthyroid, hypothyroid, and hyperthyroid rats. Immunostaining of a transversal section of the CL at 60× magnification in
Progesterone synthesis stimulation in vitro
As a first step to understand the action of T3 on CL function during pregnancy, we studied whether they are able to stimulate P4 output in vitro using two different approaches. First, we assessed the capacity of the CL of pregnancy to produce P4 in a primary culture in the presence of T3, using as substrate pregnenolone (P5) and comparing the capacity of normal CL and those from hypothyroid rats (Fig. 6). The experiment shows that T3 alone did not modify the output of P4 by the CL of pregnancy. Although, the percentage of P4 output was always higher in the hypothyroid CL of pregnancy, the difference never achieved significance. Since CL of pregnancy cultures are difficult to maintain for long periods of time, we decided to attempt a second approach. We evaluated the action of T3 on P4 output using luteinized granulosa cell culture; these cells are more resistant in culture and therefore we could test their basal and stimulated capacity to synthesize P4 for 48 hours, using different hormones that are established P4 synthesis stimulators like PRL, LH and PGE2. The analysis of P4 in the culture media showed that PGE2 and LH significantly increased the synthesis of P4 compared to basal condition and under the stimulation with PRL. Although PRL stimulated P4 synthesis, it did not achieve a significant difference compared to the controls. Using this model, we did not observe any significant effect of T3 addition on basal nor stimulated P4 synthesis (Fig. 6).
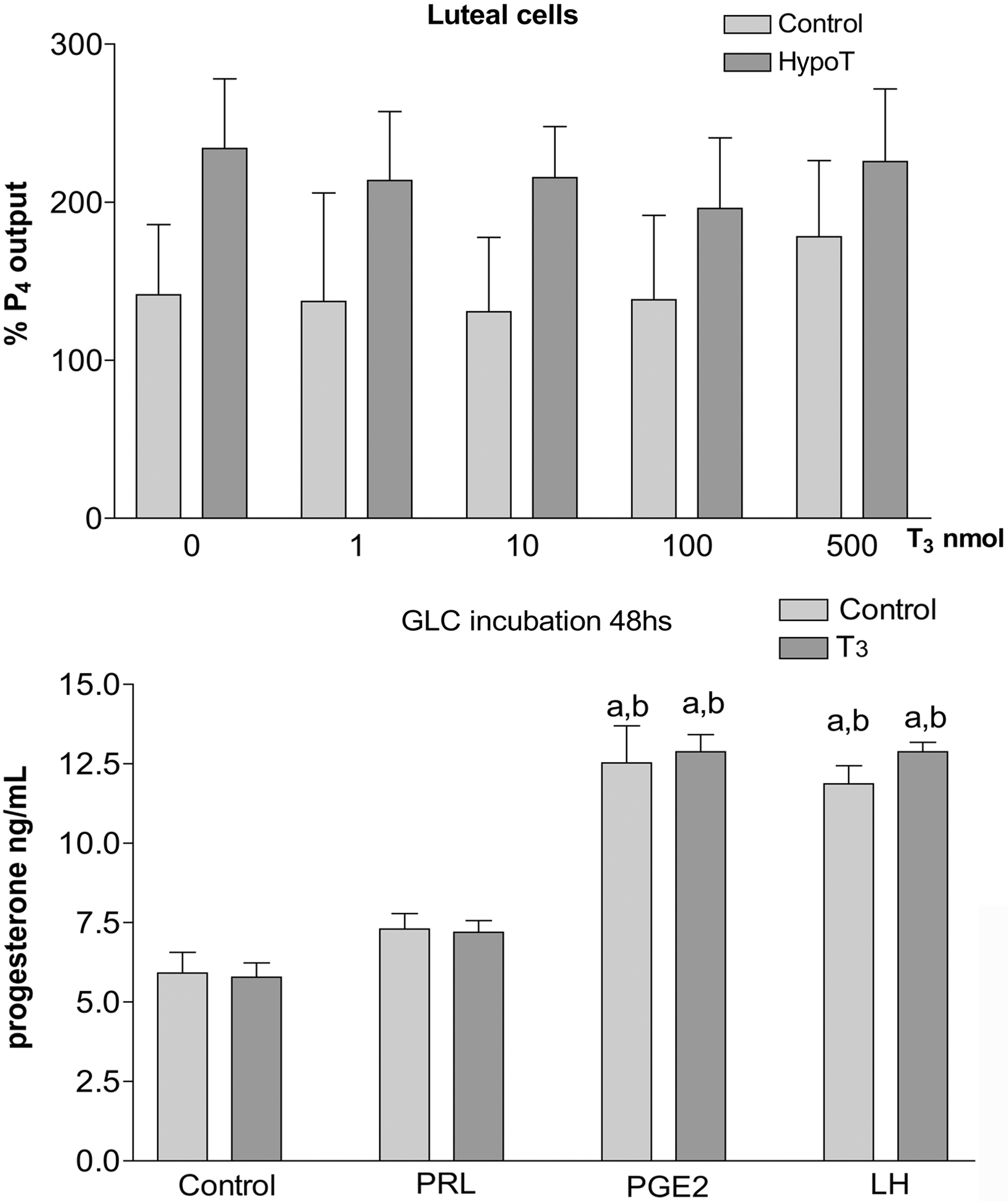
In vitro effect of T3 on luteal P4 synthesis.
Discussion
Experimental findings indicate that adequate circulating TH levels are needed to support CL formation and pregnancy (5,6,11). Previous studies from our laboratory demonstrate that hypothyroidism impairs CL function during gestation, delaying the onset of parturition in the rat (12,13). On the contrary, hyperthyroidism advances P4 withdrawal and the onset of parturition (14,15) suggesting that TH participates in the maintenance and function of CL of gestation in the rat.
The physiologic variation in TH levels during gestation responds to the demands of the fetus. The thyroid status during gestation, in this study, confirms the results we have reported before (12). The concentration of T4 diminished as gestation progressed in order to maintain adequate T3 levels to assure a proper TH environment for both maternal and fetal tissues. This regulation may be possible since it has been demonstrated that uterine deiodinase D2 expression increases during gestation favoring the metabolism of T4 to T3 (33).
To test the possibility of a direct action of TH on the CL, we determined the presence of TR mRNA and protein. As expected, our results demonstrated that the CL of pregnancy express mRNA for TRα1, TRα2, and TRβ1 but not TRβ2, as has been reported in other rat tissues such as placenta, mammary gland, liver, and kidney (34 –36). The expression profiles of TRα1, TRα2, and TRβ1 mRNAs are similar to the results previously described in placenta at the end of gestation (34). Our results show the expression profile of TRs during the entire lifespan of the CL, since the beginning of gestation until the moment of functional and structural luteolysis, immediately before and after parturition respectively. The relative expression of TRα1, TRα2, and TRβ1 transcripts increased accompanying the growth in size of the CL during the first half of gestation, and decreased significantly thereafter. TRβ1 expression remains elevated after parturition, while TRα and TRα2 decline during luteolysis along with the size of the CL (37). The increase observed in the relative expression of these receptors during early gestation suggests a physiologic and positive regulation, and consequently a possible function during this period. The expression of the TRα2 transcript followed a similar pattern as TRα1 confirming that both isoforms are generated by the same promoter and may be under the same type of regulation during gestation at luteal level, as has been demonstrated in other tissues (38). Despite that TRα isoforms are expressed under the control of the same promoter their actions are dissimilar. The TRα2 isoform does not bind T3 but interacts with the same DNA response elements as TRα1 and TRβ1 (38). This type of interaction may allow TRα2 to attenuate the effect of TRα1 and TRβ1 receptors bound to their ligands (39). Taking all together, this evidence shows that TRα1 and TRβ1 may function as promoters of CL development and survival during gestation and TRα2 may participate as a regulator of this process.
To further understand the functions of the TRs, we also evaluated the protein expression and regulation of the predominant receptors that mediate TH actions, TRα1 and TRβ1, during gestation and postpartum. The upregulation of TRα1 and TRβ1 expression on G19 together with the diminution of T4, could respond to the necessity of maintaining adequate TH action in those tissues critical for the maintenance of pregnancy, among them, the CL function that drives the timing of parturition. The diminution of the luteal expression of TRs on day 21, a few hours before parturition, reinforces the hypothesis of a participation of TH on CL function and on the timing of parturition. Moreover, the presence of TRβ1 on G19, as shown by immunohistochemistry, confirms its presence specifically in luteal cells and its nuclear localization, suggesting that such participation is mediated by a direct action as a transcription factor in this tissue.
The expression of TRα1 and TRβ1 protein throughout gestation showed a pattern similar to the profile of circulating P4. Furthermore, on G21, when functional luteolysis has proceeded, evidenced by the prompt decline in P4, the expression of TRα1 and TRβ1 descend concomitantly. The parallelism between P4 and TRs expression allows us to speculate that P4 may regulate the luteal expression of TR at the protein level but not at the posttranscriptional level, since the mRNA expression remains unchanged. At the end of gestation, when luteolysis is established, TRα1 and TRβ1 protein levels diminished significantly without significant changes in mRNA, suggesting that degradation of these proteins may be part of the several processes that take place during functional and structural luteolysis (37). On the other hand, luteal weight loss and the increase in number of apoptotic cells occur in the postpartum regressing CL, and are considered hallmarks of this process (40). T3 has strong trophic actions in the ovary, since it promotes cell proliferation and survival in human granulosa cells, and prevents cell cycle arrest and induction of apoptosis in rat ovarian granulosa cells in vitro (41,42). These actions of TH on the ovary overlap with the prosurvival actions of P4 on the ovary during gestation, which may be indicative of a possible interaction between both pathways. Thus, the fall in TRα and TRβ protein between G19 and G21 may be an early signal of luteolysis, and through the withdrawal of trophic signals, favor the subsequent regression and apoptosis of the luteal cells. Thus, TH action may not be further needed after functional luteolysis and consequently, the expression of TRs is downregulated.
The role of TH signaling on the reproductive process has been explored in other animal models. For example, knockin mice with the PV mutation in TRα1, which abolishes T3 binding and inhibits transactivation of the wild-type TRs, show dwarfism, increased mortality, reduced fertility, and mild thyroid failure (43). Reduced fertility was associated with decreased frequency of successful pregnancies and litter size, and this phenotype was attributed to the alteration in the receptor, since T3 and T4 showed concentrations slightly above normal values. Conversely, mice with a TRβ1 with the PV mutation display a normal reproductive phenotype (44). Female mice lacking TRα and TRβ receptors rarely become pregnant and were deficient in nurturing any pups born (45). On the other hand, mice lacking either TRα1 or TRβ1 display normal fertility (46). The lack of concordance between the double knockout versus TRα or TRβ knockout suggests that both isoforms may compensate for each other and that they may have overlapping functions in the reproductive process. The discrepancies between the knockin and knockout models may be due because, at least in vitro, the mutation of the receptor in the knockin model exhibits a potent dominant negative activity inhibiting the wild-type TRs transcriptional activity (44). Unfortunately, none of these studies identify which are the target organ/s affected by TH signaling that may be, at least in part, responsible for the alteration in the reproductive performance.
To our knowledge, this is the first study demonstrating expression and regulation of TR mRNA and protein in the CL of pregnancy in mammals. Despite TR presence and regulation during gestation on the CL, our in vitro results from whole luteal cell culture, failed to demonstrate a direct association between TH signaling and the main function of CL in the rat, namely, P4 synthesis on day 19 of gestation. The lack of an association between TH signaling and P4 synthesis may be due to the fact that the presence of blood and nonluteal cells in the culture may modify the responsiveness of luteal cells to P4-inducing factors. It is tempting to speculate that TH may exert another physiological function in CL such as the regulation of cell cycle or survival. Furthermore, some effects may be indirect through alterations in prostaglandin synthesis and metabolism in other target tissues, which, in turn, affect functional luteolysis, as we have previously demonstrated (13,14).
In conclusion, in this study we describe the expression of TR isoforms in the CL during pregnancy and the postpartum period, identifying the CL of pregnancy as a TH target organ during gestation. TRs protein expression is modulated in this tissue in coincidence with the regulation of P4 metabolism, and the abrupt changes before and after delivery suggest a role for THs during luteolysis. However, TH actions on the CL do not seem to be related to a direct regulation of P4 synthesis. Due to the consequences of thyroid dysfunction on domestic animal and human reproduction, the challenge remains to elucidate the exact functions of TH on CL and their implication on reproductive performance.
Footnotes
Acknowledgments
This work has been supported by grants PIP 2298/09 from CONICET (Consejo Nacional de Investigaciones Científicas y Técnicas, Argentina), PICT-R 32529 from the Agencia de Promoción Científica y Tecnológica, and SeCTyP 06/ M045. Universidad Nacional de Cuyo. Argentina. DFC-C, LMV-R, SRV, GAJ and MBH are Career Scientists from CONICET, PBN and ALR have fellowships from CONICET. The authors thank Dr. Parlow and the NHPP (National Hormone and Pituitary Program, Harbor–UCLA Medical Center, Torrance, California) for the materials they provided for the double-antibody RIA to measure TSH. The authors are grateful to Tiffany Weidner for the editing of the manuscript and Norma Carreño for her invaluable technical assistance.
Author Disclosure Statement
No competing financial interests exist.