
Abstract
Using gene expression profiling, we detected differential thyrotropin receptor (TSH-R) expression during human T-cell development in the thymus. This expression pattern indicated a potential role for the TSH-R within the thymus, independent of its function in the thyroid gland. Here, we demonstrate that TSH-R expression is thymus-specific within the immune system. TSH was able to bind and activate the TSH-R present on thymocytes, thereby activating calcium signaling and cyclic adenosine monophosphate signaling pathways. Mice lacking functional TSH-R expression (hyt/hyt mice) were shown to have lower frequencies of DP and SP thymocytes compared to their heterozygous littermates. Moreover, addition of TSH to co-cultures of human thymocytes enhanced T-cell development. Thus, TSH acts as a previously unrecognized growth factor for developing T cells, with potential clinical use to enhance thymic output and thereby the functional T-cell repertoire in the periphery. The direct effects of TSH on thymocytes may also explain the thus far enigmatic thymic hyperplasia in Graves' disease.
Introduction
T-
Quantitative defects in the peripheral T-cell pool are associated with several clinical conditions such as specific primary immunodeficiencies, HIV and other chronic viral infections, and after hematopoietic stem cell transplantation (HSCT) (7 –9). Moreover, during physiological aging, T-cell numbers and especially TCR diversity decline (10). In many of these conditions, reconstitution of the T-cell pool is essential to reduce the risk of opportunistic infections, reactivation of latent viruses, malignancy, and autoimmunity (11,12).
T-cell-pool reconstitution depends on active thymopoiesis as well as on peripheral expansion of T cells. Peripheral expansion increases total T-cell numbers without increasing TCR diversity. In contrast, thymopoiesis selectively complements the naive T-cell pool and thus increases TCR diversity and thereby T-cell function. Renewed thymopoiesis is therefore essential for functional T-cell recovery and repertoire development (13).
Many strategies to enhance T-cell reconstitution have been investigated without success, largely because effects were mainly restricted to peripheral expansion of T cells (14). Therefore, detailed understanding of intrathymic T-cell development and the factors that regulate this process is necessary for the development of new treatment modalities that selectively stimulate thymopoiesis, resulting in the recovery of a functional and diverse T-cell pool.
We previously performed microarray analyses on consecutive human thymocyte subsets to identify factors involved in the regulation of T-cell development (5). Upon reinvestigating these data, we noticed marked upregulation of several hormone receptors coinciding with T-cell lineage commitment. Specifically, thyrotropin receptor (TSH-R) was abundantly expressed during the earlier stages of T-cell development (Fig. 1A), where lineage commitment, proliferation, and TCR gene rearrangements take place (4,5,15). This expression pattern suggests that TSH, the main natural ligand for the TSH-R, might be involved in T-cell development.
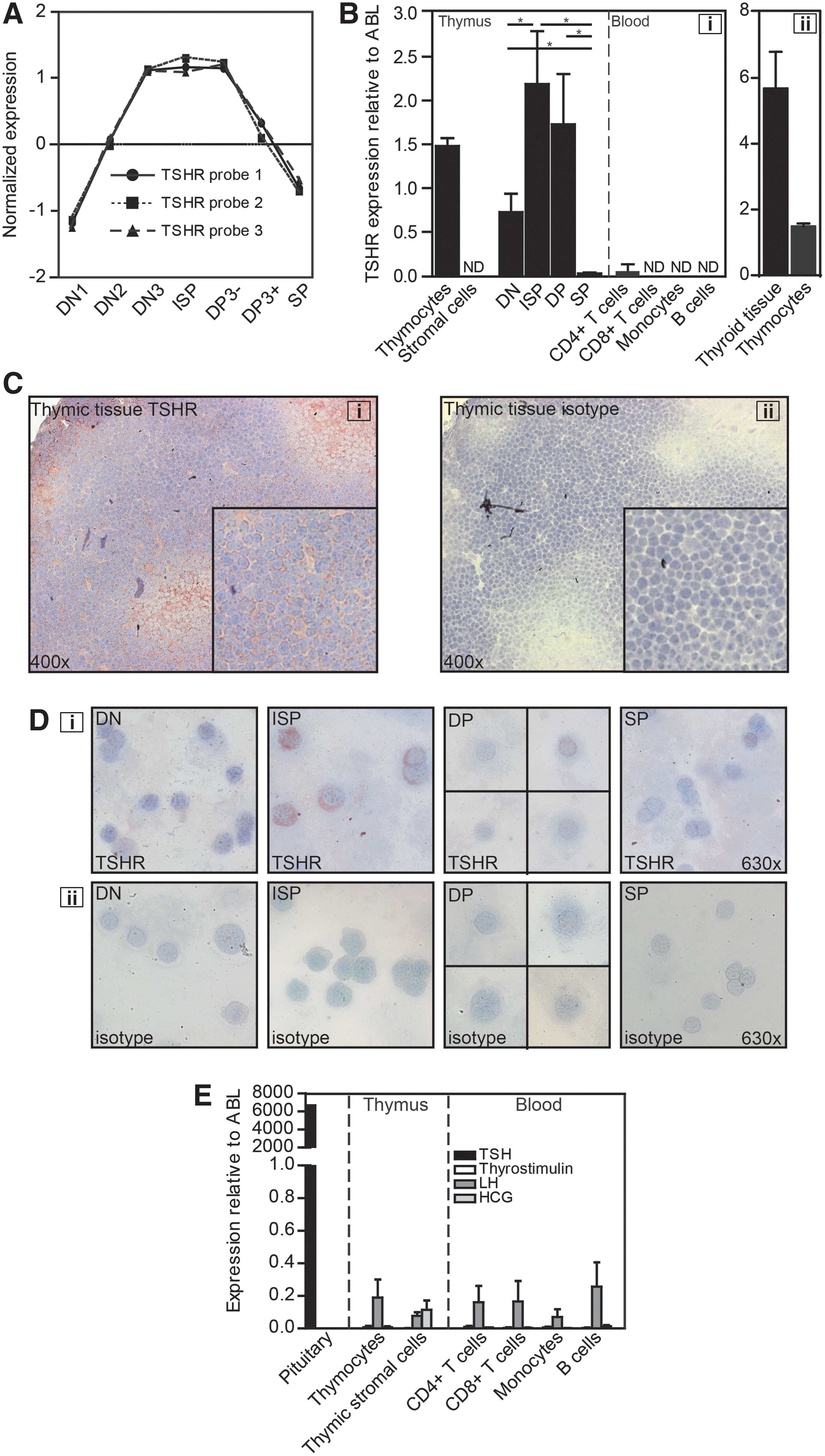
The thyrotropin receptor (TSH-R) is selectively expressed on thymocyte subsets, without expression on peripheral mononuclear blood cells.
Immune–endocrine interactions have been recognized for several decades. Many hormones and their receptors are expressed within the immune system, including the thymus (6,16). A functional role for growth hormone, steroid hormones, and reproductive hormones in the thymus is now clearly defined (6). However, considerably less is known about the expression and function of hormones of the hypothalamus–pituitary–thyroid (HPT) axis in the thymus. TSH is an important hormone within the HPT axis where it regulates the endocrine function of the thyroid gland through TSH-R signaling. Although the TSH-R is traditionally thought to be expressed exclusively within the pituitary and the thyroid gland, several studies have now identified novel functions for TSH and TSH-R signaling in various organ systems, including the immune system. TSH-R expression has been described in human (17,18) and rat thymus (19), as well as in several other hematopoietic cell types (20,21), albeit without identification of specific subpopulations. Moreover, a clear association has been described between Graves' disease and thymic hyperplasia (18), characterized mainly by an increase in developing thymocytes (22). As thymic hyperplasia is mainly seen in Graves' disease and less in other forms of hyperthyroidism (23), TSH or TSH-R antibodies are more likely to be involved than thyroid hormone, suggesting a functional role for the TSH-R present in the thymus. Functional studies investigating the role of TSH and its receptor within the immune system, however, have produced contradictory results.
We therefore set out to investigate thymic TSH and TSH-R expression in greater detail and to investigate the possible roles for TSH-R signaling during T-cell development. We here report that the TSH-R is functionally expressed on specific thymocyte subsets. Moreover, we provide first evidence that TSH-R signaling might be a previously unrecognized growth and differentiation factor for thymocytes in both mice and man. We propose that the TSH-R might be an interesting novel treatment target to enhance thymic output in T-cell immunodeficiencies. Finally, we discuss the possibility that our results might explain the previously enigmatic thymic hyperplasia described in Graves' disease patients that have specific TSH-R autoantibodies.
Materials and Methods
Mice
Balb-c hyt −/− and Balb-c Rag2 −/− mice, originally purchased from the Jackson Laboratory (Bar Harbor, ME), were bred and maintained in the specified pathogen-free (SPF) breeding facilities of the animal facility, Erasmus Medical Center, Rotterdam, The Netherlands.
Breeding pairs of hyt/hyt mice consisted of hyt −/− or hyt +/− males and hyt +/− females. To restore thyroid hormone levels, hypothyroid hyt −/− mice received T3 (3,3′,5-Triiodo-L-thyronine sodium salt; Sigma-Aldrich Logistic GmbH, Schnelldorf, Germany; 0.2 μg/mL) supplementation in the drinking water directly from weaning.
This study was performed in accordance with the legal regulations in The Netherlands and with the approval of the local institutional Animal Ethical Committee (Permit Number: EUR1559).
Thymus samples
Thymi were obtained from children (0–6 years of age) undergoing cardiac surgery at Erasmus Medical Center, Rotterdam, The Netherlands. The children did not have any immunological abnormalities. Informed consent was given by the parents. Collection of the samples was approved by the local Medical Ethical Committee. Thymocytes and epithelial cells were isolated as described previously (5). Thymocytes were isolated by cutting thymic lobes into smaller pieces and squeezing them through a metal mesh. They were then washed and frozen viably for further analyses. For all experiments, we used thymocytes obtained after Ficoll density separation. Stromal cells were isolated from thymic tissue after removal of the majority of thymocytes. To obtain a single stromal-cell suspension, tissue was digested during three sequential incubation steps (20 min, 37°C) with dispase II, collagenase, and DNAse, followed by three sequential incubation steps (15 min, 37°C) with trypsine and DNAse. Remaining CD45+ thymocytes were depleted by AutoMACS using CD45 beads (Miltenyi Biotec GmbH, Gladbach, Germany).
Cell lines
The cell line JP09, a Chinese Hamster Ovary (CHO) cell line stably transfected with the human TSH-R, was provided by Prof. G. Vassart, Université Libre, Brussels, Belgium (24). Wild type CHO cells were provided by Dr. L.J. Hofland, Erasmus Medical Center, Rotterdam, The Netherlands (Sigma-Aldrich). The cell line OP9DL1, a mouse BM stromal cell line expressing the Notch ligand Delta-like-1, was provided by Dr. B. Vandekerckhove, University Hospital, Ghent, Belgium (25,26).
Flow cytometry and cell sorting
Antibodies used for flow-cytometric analyses and cell sorting are summarized in Supplementary Table S1 (Supplementary Data are available online at
The different developmental stages of thymocytes can be distinguished phenotypically by flow cytometry based on expression of surface molecules (5). In humans as well as mice, four main developmental stages can be distinguished based on CD4 and CD8 surface expression. T-cell precursor cells entering the thymus do not express CD4 and CD8, and therefore are referred to as double negative (DN). In mice, DN thymocytes can be further subdivided into CD25− CD44+ (DN1), CD25+ CD44+ (DN2), CD25+ CD44− (DN3), and CD25− CD44− (DN4) thymocytes. In humans, DN thymocytes can be subdivided into CD34+ CD38− CD1a− (DN1), CD34+ CD38+ CD1a− (DN2), and CD34+ CD38+ CD1a+ (DN3) thymocytes. DN thymocytes subsequently develop into immature single positive (ISP) cells that express CD4 in humans but CD8 in mice. In the next stage of development, thymocytes are called double positive (DP) cells expressing both CD4 and CD8. Moreover, during the DP stages of development, CD3 expression will occur, resulting in two subpopulations of DP thymocytes: early DP thymocytes not expressing CD3 (DP3−) and late DP thymocytes expressing CD3 (DP3+). Finally, thymocytes become single positive (SP) mature T-lymphocytes that express either CD4 (SP4+) or CD8 (SP8+) (5,27).
Microarray analysis
To examine the effect of TSH on gene expression profiles in thymocytes, sorted ISPs were cultured with 1 nM recombinant human TSH (rhTSH; Thyrogen; Genzyme Europe BV, Naarden, The Netherlands) dissolved in phosphate-buffered saline (PBS)/0.1% BSA or vehicle. RNA was extracted using RNeasy MinElute columns (Qiagen, Hilden, Germany). The quality of the RNA was assessed using the Agilent 2100 BioAnalyzer (Agilent Technologies, Santa Clara, CA).
Biotin-labeled cRNA was prepared using the Affymetrix two-cycle target labeling kit (Santa Clara, CA) and the MEGAscript T7 kit (Life Technologies, Grand Island, NY). Subsequently, 15 μg of biotin-labeled cRNA was fragmented and hybridized to a Human Genome U133 Plus 2.0 microarray. Thereafter, microarrays were washed and stained using the Affymetrix Fluidic Station F450 and scanned at 570 nm.
GeneScan analysis for in-frame TCRB gene rearrangements
To determine TCR diversity, we performed GeneScan analysis with DNA of sorted ISP thymocytes (defined as CD4+ CD8− CD3−), after being cultured for 3–6 h in the presence of 1 nM rhTSH or vehicle, to determine in-frame Vβ-Jβ gene rearrangements with multiplex polymerase chain reaction (PCR) as previously described (28).
RNA isolation and real-time quantitative PCR analysis
RNA was extracted using RNeasy columns (Qiagen), and RNA was reverse transcribed into cDNA as described (5). For detection of gene expression levels of the TSHR, TSHB, GPHB5 (thyostimulin, β-subunit), LHB (luteinizing hormone, LH), and CGB (human chorionic gonadotropin, hCG), primers and probes were obtained from Applied Biosystems (Assay-on-demand™; Foster City, CA). Expression levels were normalized to ABL gene expression, which is expressed at very stable levels in both healthy and leukemic lymphoid cells and is thus an optimal choice as a housekeeping gene (29). Primers and probes for the detection of gene expression levels of CXCL2, CXCL3, EGR3, MAFF, PTX3, ATP6, CREB1, CREB5, GNA11, GNAS, HLA-DRA, PDE3B, and PDE4D were designed using OLIGO primer analysis software (Molecular Biology Insights, Cascade, CO). Sequences are available upon request.
Immunohistochemistry
Immunohistochemistry was done as described previously (30). Thymic and thyroid cryosections (6 μm) and cytospins of sorted thymocyte subsets were fixed in acetone/0.05% H2O2 and stained overnight with a TSH-R specific antibody (MCA1571, 20 μg/mL; AbD Serotec, Dusseldorf, Germany) or an isotype control (BD Biosciences). Subsequently, sections were incubated with a goat-antimouse-horseradish peroxidase-labeled monoclonal antibody (Dako B.V., Glostrup, Denmark). For visualization of horseradish peroxidase activity, we used 3-amino-9-ethylcarbazole substrate (Sigma Co., St. Quentin Fallavier, France) dissolved in 50 mM sodium acetate/0.02% hydroxyperoxide. Sections were embedded in Kaiser's glycerol/gelatin (Boom B.V., Meppel, The Netherlands).
Labeling studies
Labeling studies were performed as described previously (31). To demonstrate binding of TSH to the TSH-R on thymocytes, 50 mg of thymocyte membrane was incubated for 1 h at 4°C in medium containing 10 mM Tris-HCl and 1% BSA (pH 7.4) with 125I-labeled bovine TSH (bTSH; 50 μCi/μg; BRAHMS, Berlin, Germany) with or without addition of an overload of unlabeled bTSH (250 mIU/mL; Sigma-Aldrich). JP09 cell membranes served as a positive control. Radioactivity was counted in a γ-counter. Specific binding was calculated as the difference between binding in the absence and in the presence of unlabeled bTSH.
Calcium flux
Calcium-flux experiments were essentially performed as described previously (32). Additionally, in order to allow distinction between thymocyte subsets, cells were stained for CD3, CD8, and CD4.
In vitro T-cell differentiation
To investigate the effect of TSH on human thymocyte development in vitro, immature CD34+human thymocytes were isolated from pooled total thymocytes using autoMACS beads (Miltenyi Biotec GmbH). Subsequently, these CD34+ positive immature thymocytes were cultured in fetal thymic organ cultures (FTOC) and OP9DL1 co-cultures as described previously (33,34). Twice weekly, rhIL-7 (1 ng/mL), rhSCF (5 ng/mL), and different concentrations of rhTSH (0.1–10 nM) were added.
Statistical analysis
The exact Mann–Whitney U-test was used for statistical comparisons. Statistical analyses were performed with SPSS v15.0 (SPSS, Inc., Chicago, IL). A p-value of <0.05 (two tailed) was considered statistically significant. Error bars are expressed as standard error of the mean.
Results
TSH-R expression in the thymus
Upon extensive data mining of our previously published gene expression profiling studies of human thymi (5), with the aim of identifying novel regulators of T-cell lineage commitment, the expression pattern of the TSH-R was striking (Fig. 1A). Differential TSH-R mRNA expression was validated by real-time quantitative PCR (RQ-PCR; Fig. 1B-i) showing an expression pattern similar to that obtained with microarray analyses, with the highest expression of TSH-R mRNA in ISP and DP thymocytes (Fig. 1B-i). In addition, TSH-R mRNA was found to be expressed exclusively on thymocytes (Fig. 1B-i). No TSH-R mRNA expression was observed in sorted peripheral blood mononuclear cells (Fig. 1B-i). TSH-R expression in thymocytes was approximately four times lower than in thyroid tissue (Fig. 1B-ii). In mice, the TSH-R mRNA expression pattern in developing thymocytes was comparable to the human situation (data not shown).
To investigate TSH-R protein expression, frozen human thymic tissues were analyzed by immunohistochemistry. In thymic tissues (Fig. 1C-i) as well as in thyroid control tissue (data not shown), a clear membrane staining for the TSH-R was found. No immunoreactivity was observed when tissues were stained with isotype control (Fig. 1C-ii). Cytospin stainings of sorted thymocyte subsets also demonstrated TSH-R immunoreactivity in all thymocyte subsets, in a pattern similar to the mRNA expression data, with the highest reactivity in ISP and DP thymocytes (Fig. 1D-i), while no immunoreactivity was observed in thymocytes stained with isotype control (Fig. 1D-ii). Thus, the TSH-R is selectively expressed in developing T cells within the thymus, especially during ISP and DP stages of development.
Besides TSH, which is the natural ligand for the TSH-R, several other hormones (e.g., LH, hCG, and thyrostymulin) can also bind and activate the TSH-R, albeit with much lower affinity than TSH (35,36). We therefore investigated mRNA expression of these hormones in thymocytes, thymic stromal cells, and peripheral blood mononuclear cells. TSH expression was undetectable in thymocytes, thymic stromal cells, and mature immune cells, while high TSH expression, as expected, was detected in pituitary tissue (Fig. 1E). Among the other TSH-R binding hormones, only LH mRNA was occasionally found in all the cell types tested, albeit at very low levels (Fig. 1E).
TSH binds and activates the TSH-R present on thymocytes
Scatchard analysis revealed that 125I-labeled bTSH bound to human thymocytes with a dissociation constant (Kd 0.55 nM) similar to that obtained with TSH-R positive control cells (JP09; Kd 0.42 nM). Although we were not able to demonstrate low affinity binding sites, probably due to the use of relatively low doses of Tris in the buffer (37), the presence of high affinity binding sites indicates that TSH binding on thymocytes is indeed via the TSH-R. Maximal binding of bTSH by human thymocytes was 34 fmol/mg protein; maximal binding of bTSH by JP09 membranes was 57 fmol/mg protein (Fig. 2A).
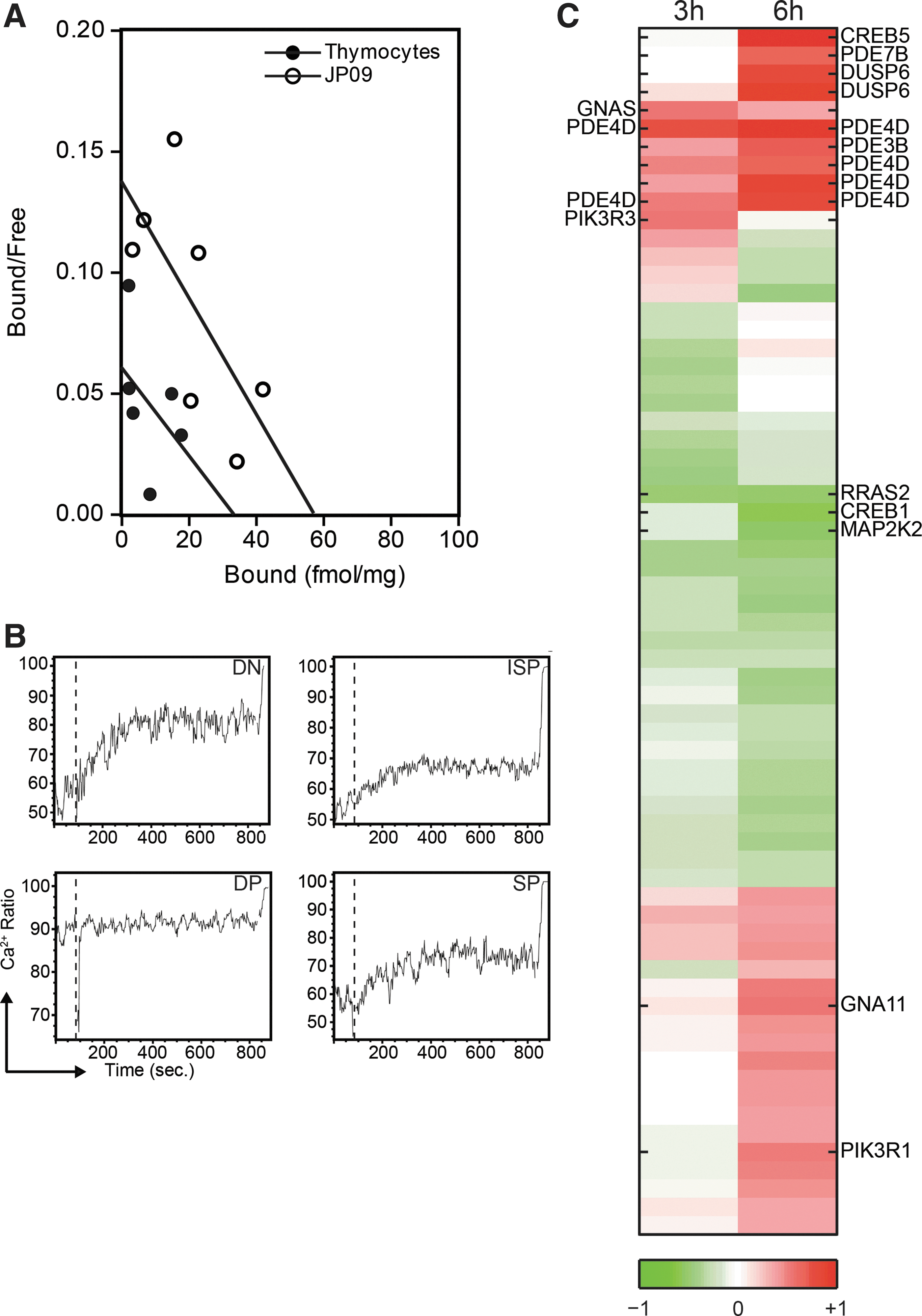
TSH binds and activates the TSH-R expressed on human thymocytes.
Thyroidal TSH-R activation results in the activation of several second messenger pathways, including the PIP2/Ca2+ and the adenylate cyclase/cAMP transduction cascade (38). We therefore investigated whether TSH activates these pathways in thymocytes. After stimulation with rhTSH, free intracellular calcium increased in ∼15% of thymocytes in all subsets except for DP thymocytes that have very high (∼90%) spontaneous calcium fluxes (Fig. 2B), virtually excluding detection of additional free calcium in this subpopulation. In addition, we stimulated flow-sorted ISP thymocytes, which have high TSH-R expression for three or six hours with rhTSH, and subsequently performed microarray analysis. Gene-expression analysis of genes involved in G-protein-coupled receptor (GPCR) signaling demonstrated differential expression of many probe sets in rhTSH relative to vehicle-stimulated ISPs, suggesting that TSH-R signaling is induced (Fig. 2C). After three hours of culture, 4 of the 399 probe sets involved in GPCR signaling were more than 1.5-fold differentially expressed. This number increased to 14 after six hours of culture. Importantly, after six hours of culture with rhTSH, expression of the transcription factor CREB5, an important target gene of cAMP signaling, was upregulated. Collectively, these data demonstrate that TSH binds and signals via the TSH-R on thymocytes.
TSH-R activation in ISP thymocytes results in changes in gene expression profile
To identify molecular pathways by which TSH-R signaling might be involved in T-cell development, the above-described microarrays of flow-sorted ISP thymocytes were analyzed for differential expression of genes important for T-cell development. After three hours of culture, 32 probe sets were more than twofold differentially expressed in rhTSH relative to vehicle-stimulated ISPs. This number increased up to 180 probe sets after six hours of culture. A substantial number of these probes represented genes that are known to play a role in different processes involved in T-cell development and TSH-R signaling (Fig. 3A), such as HLA, TGF-βR, MAF, and EGR. Many of the differentially expressed genes function to drive T-cell development after the pre-TCR-β-selection point (e.g., Egr, c-maf, various HLA Class I molecules). The microarray analyses were validated by performing RQ-PCR analyses for selected genes that were found to be differentially expressed, demonstrating a comparable pattern (Fig. 3B). Moreover, we performed GeneScan analysis of TCRB rearrangements in DNA from flow-sorted ISP thymocytes stimulated for six hours with vehicle or rhTSH. In previous studies, it was demonstrated that upon TCR rearrangements and subsequent selection processes of in-frame rearrangements, a triplet-nucleotide peak pattern becomes visible in GeneScan analysis (5). As this pattern is more clearly visible in the rhTSH-stimulated ISP thymocytes, these data suggest that TCRB rearrangements and subsequent selection occurred earlier in rhTSH-stimulated ISP thymocytes (Fig. 3C). Overall, these data, together with the marked increase of TSH-R expression on ISP and early DP thymocytes, indicate that one of the functions of TSH-R signaling is to enhance post β-selection T-cell differentiation and proliferation.
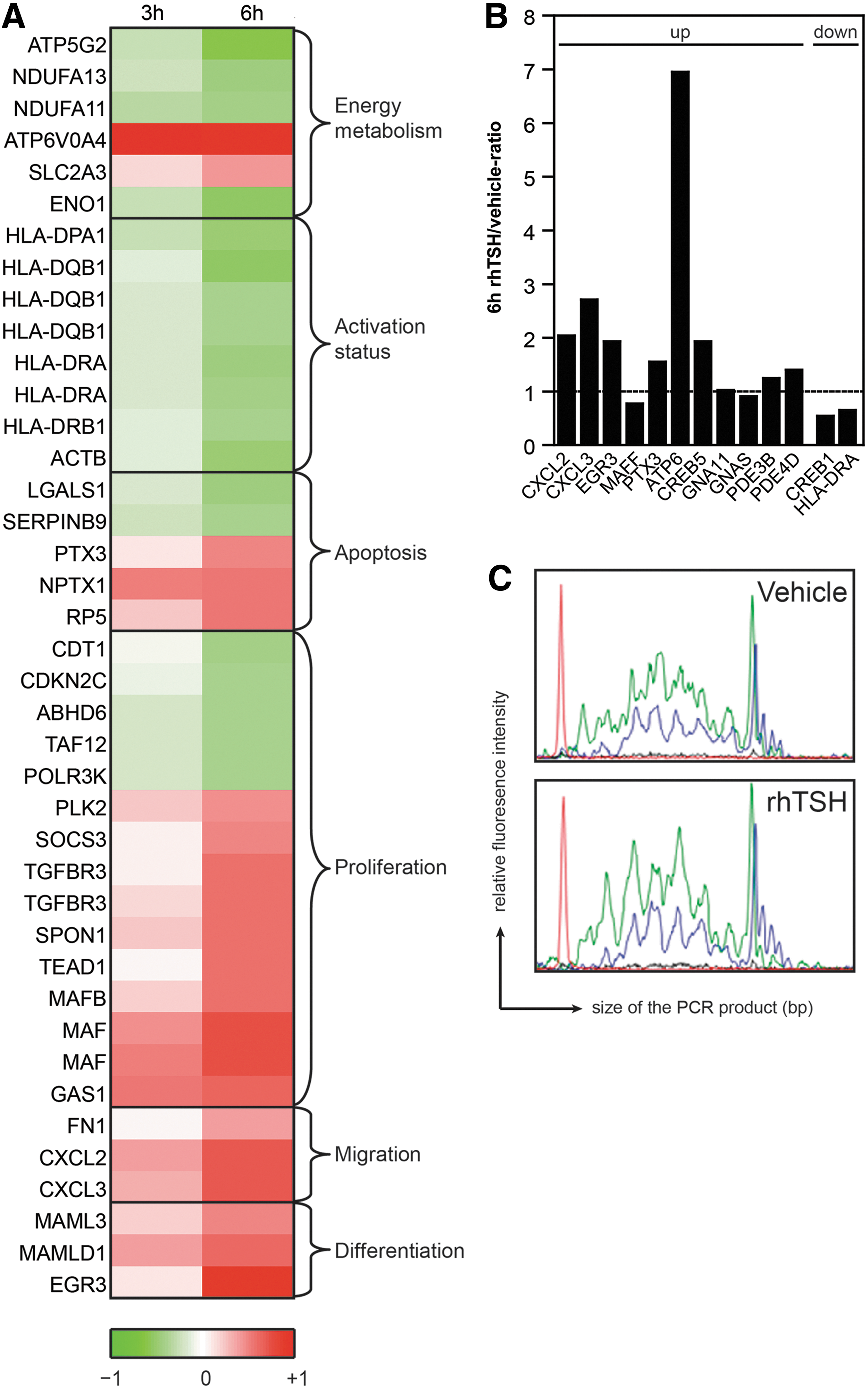
Induction of TSH-R signaling in ISP thymocytes induces changes in expression of genes involved in T-cell development.
Lack of functional TSH-R expression alters normal thymic composition and development
To investigate whether TSH-R signaling is involved in T-cell development in vivo, we used hyt/hyt mice as an in vivo model. Hyt/hyt mice have a point mutation in the TSH-R gene resulting in severely diminished TSH binding capacity and TSH-R functionality (39). Consequently, these mice are hypothyroid. In order to study the effects of absence of TSH-R signaling on the development of the immune system without additional effects of thyroid hormone abnormalities, the mice were supplemented with T3 from weaning. Flow-cytometric analysis of thymocyte subsets demonstrated that hyt −/− mice had a relative block in T-cell development at the ISP or DP stages, resulting in a decreased frequency of DP and SP thymocytes compared to heterozygous littermates (Fig. 4). This did not result in decreased T-cell frequencies or numbers in blood and spleen (data not shown), most likely due to peripheral homeostatic expansion.
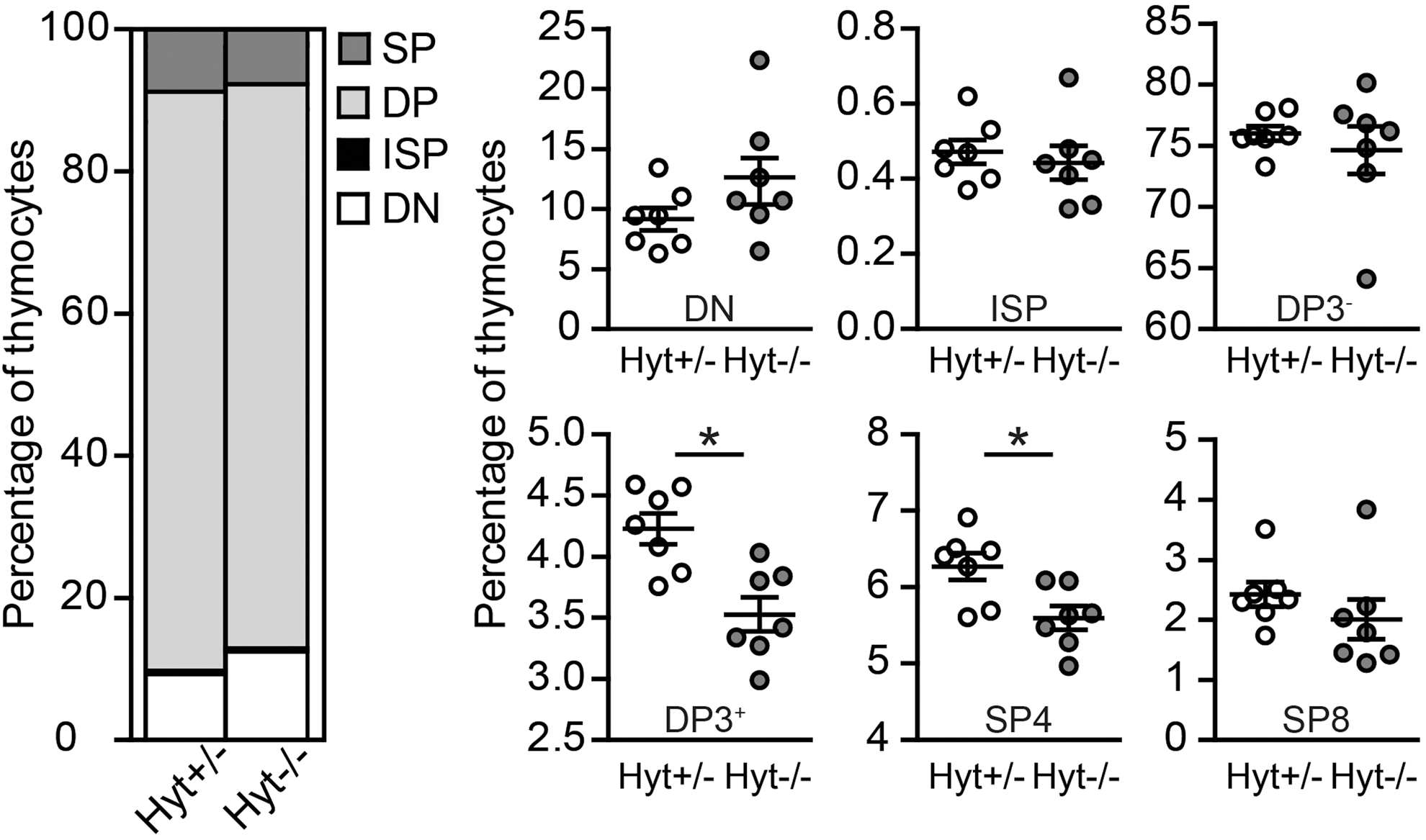
Lack of TSH-R signaling alters normal thymic composition in mice. Flow cytometric analysis of the thymus of eight-week-old hyt +/− and hyt −/− mice. Left panel: thymic composition, right panel: percentages of thymocyte subsets, *DP3+, p=0.007; SP4, p=0.02 (two-tailed). Data are from a representative experiment (n=7 for hyt +/− group, n=7 for hyt −/− group). DP3−, double positive thymocytes not expressing CD3 (CD4+ CD8+ CD3−); DP3+, double positive thymocytes expressing CD3 (CD4+ CD8+ CD3+); SP4, single positive thymocytes expressing CD4 (CD3+ CD4+ CD8−); SP8, single positive thymocytes expressing CD8 (CD3+ CD4− CD8+).
rhTSH accelerates human T-cell development in vitro
Because we were ultimately interested in the effects of rhTSH on human T-cell development, we tested rhTSH in two culture systems supportive for human T-cell development: the OP9DL1 culture system and the FTOC. Both culture systems support T-cell development from DN up to at least the DP stages of development, and even functional SP thymocytes can be found in some of these cultures (40,41).
Flow-cytometric analysis showed that harvested cells were fully T-cell committed, with clear differentiation into ISP and DP thymocytes (Fig. 5A). When TSH was added, an increase in the more mature ISP and DP thymocytes relative to DN thymocytes was seen in four out of five co-cultures using the OP9DL1 system (Fig. 5B) and six out of eight co-cultures using the FTOC system (Fig. 5C), indicating that rhTSH induced acceleration of human T-cell development in vitro. These experiments together also demonstrate a significant increase in the percentage of DP and SP thymocyte subpopulations in rhTSH stimulated co-cultures relative to vehicle stimulated co-cultures (Fig. 5D).
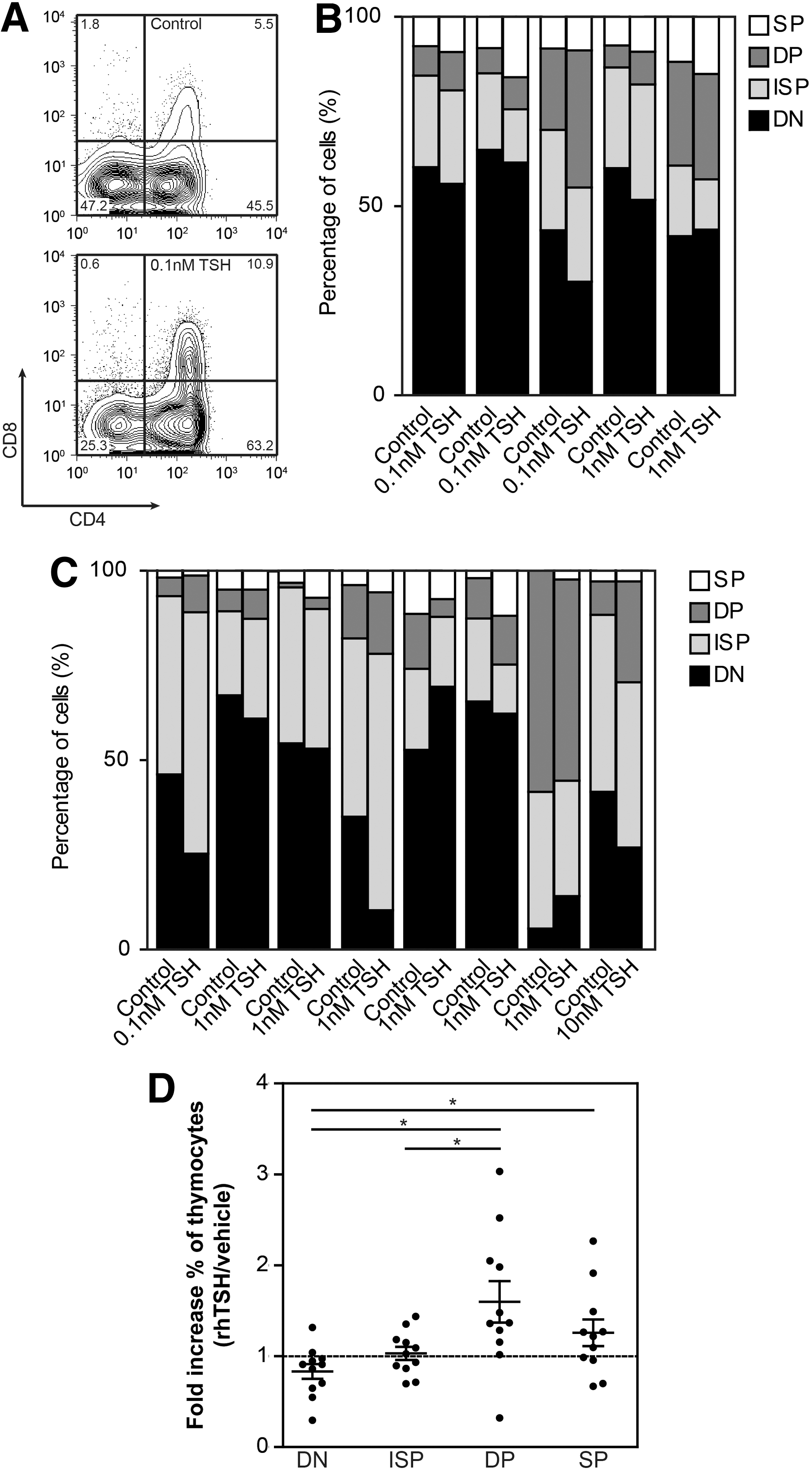
rhTSH enhances human T-cell development in vitro. Sorted human CD34+ thymocytes were cultured in T-cell development supporting systems in the presence of 0.1–10 nM rhTSH or vehicle for two to three weeks. After harvesting, relative expression of the different thymocyte subsets was measured using flow cytometry.
Discussion
The TSH-R is well recognized for its essential role in thyroid homeostasis, and was long thought to be uniquely expressed within the HPT axis. Recently, however, the TSH-R was also shown to be expressed in several nonthyroidal tissues (42 –44). Here, we provide a detailed study on TSH-R expression pattern within the immune system, and demonstrate for the first time a functional role for TSH-R signaling in T-cell development.
In the present study, TSH-R mRNA and protein expression within the immune system was found to be restricted to developing T cells, with highest expression during the ISP and DP stages of development. These findings extend earlier observations of TSH-R expression in total rodent (19) and human (17,18) thymic tissue. We did not find TSH-R expression in peripheral blood mononuclear cells, but published data on the presence of TSH-R expression in these cells are contradictory. While some studies support our data (17), others did detect TSH-R expression on peripheral blood mononuclear cells, especially monocytes (21). Also, a small population of naive T lymphocytes within the lymph node (20) and specific intestinal lymphoid cells (intraepithelial lymphocytes, IELs) (45) have been found to express TSH-R. In general, given the high sensitivity of RQ-PCR compared to other techniques used, we conclude that TSH-R expression within the immune system is restricted mainly to developing T cells within the thymus, although rare peripheral T-cell subpopulations might express the TSH-R as well. Compared to thyroid tissue, TSH-R expression in thymocytes was only about four times lower. These expression levels support the notion that TSH-R signaling in thymocytes might be functional.
Despite the fact that several studies demonstrated the presence of TSHR expression in thymic epithelial cells (19,46,47), where they are assumed to play a role in central tolerance, we did not detect TSHR expression in thymic epithelial cells in our study. This discrepancy may be caused by technical differences such as isolation methods.
We demonstrate that TSH specifically binds thymocytes and that TSH activates the PIP2/Ca2+ and adenylate cyclase/cAMP signaling pathways, which are downstream of the TSH-R. Despite the lower expression of TSH-R on DN and SP thymocytes, TSH was able to induce a calcium flux in these cells as well. TSH stimulation did not, however, increase the calcium flux in DP thymocytes, which could be related to apoptosis-induced calcium flux, a well-known feature of DP thymocytes (48). Calcium flux was already present in ∼90% of DP thymocytes before stimulation, which will hamper the detection of a rhTSH-induced calcium flux.
Functional expression of the TSH-R on developing T cells suggests the presence of a previously unrecognized neuroendocrine–immune pathway that involves pituitary-derived TSH and the thymus. Similar hormonal interactions with the thymus have been described for other hormones, such as growth hormone and cortisol (6). We found no evidence of intrathymic TSH production. Therefore, we favor the explanation that pituitary-derived TSH is involved in this neuroendocrine–immune pathway. A role for locally produced LH, the only other hormone with TSH-R binding capacities that we infrequently detected in thymic tissue, is less obvious, as it was expressed at very low levels and it has a substantially lower affinity for the TSH-R compared to TSH.
A likely mechanism whereby TSH-R signaling affects T-cell development is through combined effects on differentiation and apoptosis protection. In ISP thymocytes, rhTSH changed the expression patterns of genes involved in several processes related to T-cell development. Overall, expression levels of genes related to apoptosis induction and proliferation were reduced, while expression levels of genes that stimulate differentiation were increased.
Moreover, the abundant expression of the TSH-R during the DN3, ISP, and DP stages of development in which the processes of β-selection and TCRA rearrangements take place (5), as well as the presence of more in-frame TCRB rearrangements in TSH-stimulated ISP thymocytes, demonstrate that TSH-R signaling is involved in pre-TCR-signaling and β-selection and/or subsequent induction of TCRA rearrangements. Several changes in gene expression pattern found in the microarray analyses of TSH-stimulated ISP thymocytes support this notion. In particular, EGR3 expression was clearly upregulated in rhTSH-stimulated ISP thymocytes. Recently, it was demonstrated that pre-TCR signaling induced EGR1-3 expression in mouse thymocytes. Moreover, overexpression of EGR3 enhanced progression through the β-selection checkpoint in pre-TCR signaling deficient thymocytes (49).
Finally, similar to its effect on thyroid cells, rhTSH increased cytosolic calcium levels and CREB expression in ISP thymocytes. Interestingly, cytosolic calcium is an important signaling molecule in pre-TCR signaling (50). Moreover, pre-TCR signaling also enhances expression and phosphorylation of CREB, and several genes involved in β-selection (e.g., CD3δ, TCRα, and TCRβ) have binding sites for CREB in their enhancer or promoter regions (51,52). Additionally, inhibition of CREB activity has been shown to ameliorate the development of DP thymocytes in FTOC cultures (53).
Based on our in vitro results, we investigated the T-cell compartment in hyt/hyt mice in vivo. Hyt −/− mice had lower frequency of DP and SP thymocytes compared to their hyt +/− littermates. Consistent with our observations, others demonstrated that also the numbers of TSH-R expressing intestinal T cells were reduced in thyroid-hormone-substituted hyt −/− mice (45). In normal mice, TSH treatment did not affect these intestinal lymphocytes; effects of TSH on developing thymocytes were not investigated in this study (54).
Ultimately, we demonstrate in two human ex vivo systems that TSH-R signaling enhanced normal T-cell development, even in the presence of IL-7 and SCF, two cytokines crucially important for normal T-cell development (55).
Factors that stimulate T-cell development are of special interest for the treatment of conditions associated with T-cell deficiencies. Such factors have been sought after, but many, including IL-2 (56) and IL-7 (57), show strong effects on peripheral T-cell expansion but not on thymic output due to the expression of their receptors on peripheral mature T cells. Activation of the TSH-R, most likely by rhTSH or a TSH-analogue, might be an interesting new therapeutic target in T-cell deficiencies, as a selective effect on developing T lymphocytes is expected based on the specific expression pattern of the TSH-R in the T-cell compartment. Moreover, in contrast to the other factors, anticipated side effects of rhTSH are limited to the thyroid gland, mainly being hyperthyroidism during rhTSH treatment. These side effects can be blocked relatively easily by treatment with antithyroid drugs, which have a long history of clinical use.
Our data demonstrate that TSH-R signaling has selective stimulatory effects on human T-cell development in the thymus, which could be relevant in physiological and pathophysiological conditions. Interestingly, in the first hours after birth, a cold-induced increase in TSH levels up to 100 mU/L (TSH surge) has been described (58). As T-cell numbers are still largely increasing just after birth (59), it is tempting to hypothesize that this TSH surge could support the highly active process of T-cell development in the thymus just after birth. Moreover, an association between thymic hyperplasia, mainly due to an increase in lymphoid cells, and Graves' disease has been described (18,22), but the cause for this has remained elusive. Based on our results, we postulate that Graves' disease-associated stimulating TSH-R autoantibodies activate thymocytes, resulting in enhanced T-cell development, increased thymocyte numbers, and subsequent thymic hyperplasia. In support of this, in a pilot study, we observed that purified immunoglobulins from sera of some but not all patients with Graves' disease were capable of enhancing T-cell development in the OP9DL1 co-culture system to a comparable level as rhTSH (unpublished data). However, further research into this topic is necessary.
Collectively, our data support the notion of a novel neuroendocrine–immune interaction in which TSH-R signaling in the thymus, most likely mediated by TSH, enhances thymic T-cell development, possibly by affecting pre-TCR signaling and β-selection. Due to the selective expression pattern within the immune system, the TSH-R might be an interesting therapeutic target for primary and acquired T-cell immunodeficiencies to enhance thymic output.
Footnotes
Acknowledgments
The authors thank Edwin de Haas and Benjamin Bartol for their assistance with cell sorting, Tiago Luis for his assistance with transplantations, Peter Koetsveld for assistance with 125I-TSH labeling studies, Hans van Toor and Yolanda de Rijke for measuring T3 levels in mouse plasma samples, Sandra de Bruin–Versteeg for her assistance with the figures, and all other members of the Staal laboratory and Dik laboratory for their technical assistance. We thank Prof. G. Vassart, Dr. B. Vandekerckhove, and Prof L.J. Hofland for providing the JP09, OP9DL1, and CHO-WT cell lines. This work was supported by internal grants from the departments of Internal Medicine and Immunology. F.J.T.S. is supported in part by Kika, ZonMW, and AICR.
Author Disclosure Statement
No competing financial interests exist