
Abstract
Background:
Papillary thyroid carcinoma (PTC) is the most common malignant tumor of the thyroid gland, accounting for 74–80% of all thyroid cancers. The 1799T>A transversion is an activating mutation of the BRAF oncogene that is common in and specific to conventional PTC. We studied the prevalence, tumorigenic role, and biochemical implications of rare BRAF variants in a large cohort of patients.
Methods:
A total of 2131 fine-needle aspiration biopsy samples were collected and subjected to BRAF mutation analysis. BRAF genetic variants were analyzed by Western blot, immunofluorescence, and in silico analysis.
Results:
BRAF mutations were found in 50% (347/700) of thyroid cancers (644 PTCs, 22 anaplastic thyroid carcinomas, 34 follicular thyroid carcinomas). They were the classic (c.1799T>A, p.V600E) mutation in 96.8% (336/347) and rare genetic variants in 3.2% (11/347). In all, five infrequent BRAF alterations were detected: (i) c.1795_1797dupACA (p.T599dup); (ii) c.1801A>G (p.K601E); (iii) c.1799_1801delTGA (p.V600_K601>E); (iv) c.1799_1814>A (p.V600_S605>D); and (v) c.1798_1810delinsA (p.V600_W604>R). The last BRAF variant has never been described in the literature. Western blot analysis and immunofluorescence both revealed a variegated reactivity pattern, again emphasizing the peculiar role of every specific BRAF genetic alteration. In silico analysis of the samples studied revealed a stabilization of the “active” geometrical conformation of the B-raf enzyme associated with the activated and productive state of the kinase domain.
Conclusions:
Rare BRAF variants were found in 1.6% of all thyroid malignancies, all clustered around the codon V600, in the binding pocket named A-loop, confirming its crucial role in the enzymatic activation of the B-Raf protein. These mutations were associated mainly with the activation of key effectors in the mitogen-activated protein kinase pathway, but a simultaneous stimulation of the PI3k/Akt cascade was demonstrated in some cases. The rare BRAF variants were not generally associated with an aggressive behavior of the PTC. To our knowledge, this is the largest series of thyroid cancers analyzed to identify and functionally characterize rare BRAF variants.
Introduction
T
Receptor kinases are involved in ligand binding; via the mitogen-activated protein kinase (MAPK) pathway, they transmit several signals necessary for cell proliferation. Consisting of several key signaling components and phosphorylation events, this pathway plays a fundamental part in tumorigenesis. Activated kinases transmit extracellular signals and regulate cell growth, differentiation, proliferation, apoptosis, and migration processes.
It has been demonstrated that genes encoding modulators of the MAPK pathway have a crucial role in the malignant transformation of PTC. The BRAF gene (v-raf murine sarcoma viral oncogene homolog B1) encodes a protein belonging to the raf/mil family of serine/threonine kinases that is pivotal in regulating cell division, differentiation, and secretion. The oncogenic mutation of the BRAF gene (p.V600E), which results in the substitution of a valine with a glutamic acid at position 600, is the most common genetic alteration seen in PTC (3). The BRAFV600E mutation gives rise to constant, catalytically competent structures with continuous phosphorylation of their downstream targets (4,5); it is found in about 45% of PTC in adults and results in abnormal B-raf protein functioning (6,7). Several studies have linked the BRAFV600E mutation to aggressive clinicopathologic features, including extrathyroidal invasion, nodal and distant metastases, and inhibition of genes involved in iodine metabolism (3,8,9). Altogether, these findings define the biological effect of the mutation with regard not only to the development, but also to the progression of PTC. Other less frequent BRAF mutations, such as K601E (10), K601del (11), and V599ins (12), have been described. Interestingly, as all the above-mentioned mutations are located close to the B-raf V600 position, the site is believed to be crucially involved in maintaining the protein's inactive conformation.
Similar to the MAPK pathway, the PI3K/Akt pathway also plays an important part in thyroid tumors, and its aberrant stimulation is found in about 10–15% of follicular thyroid carcinomas (FTC) and anaplastic thyroid carcinomas and, to a lesser extent, in PTC (13). This pathway is implicated in various cellular processes such as survival, proliferation, and angiogenesis. It is noteworthy that an important crosstalk exists between the PI3K/Akt and the MAPK pathways, reflecting a more complex picture of cell resources (14).
Large-scale molecular dynamic (MD) simulations can be combined with in vitro and in vivo studies to assess the molecular role of particular mutations found in some patients. In 2006, Moretti et al. identified the novel V599ins mutation in the B-Raf kinase domain of a classical variant PTC; using MD simulations, they described a destabilization of the inactive conformation caused by this genetic event (12). In 2009, Fratev and Jonsdottir identified a strong hydrogen bond network of interactions between amino acids thought to be extremely important for catalysis (15); in 2010, the same research group used MD coupled with GRID analysis to describe the phosphorylation specificity of the activation segment B-Raf wild type, D594V, V600E, K610E (16).
These alterations in B-Raf, which has a key role in proliferation, migration, differentiation, and apoptosis, modulating the MAPK and P13K/Akt signaling pathways, were also examined by Western blot analysis and by means of a large-scale exploration of the conformational spaces using classic MD simulations.
This work aimed to elucidate the prevalence, tumorigenic role, and biochemical implications of rare BRAF alterations in thyroid carcinoma.
Materials and Methods
Fine-needle aspiration biopsy samples and DNA extraction
At our institution it is standard procedure to analyze BRAF mutations in fine-needle aspiration biopsies (FNAB) in cases of single thyroid nodules and/or nodules showing suspect ultrasonographic features. From 2131 consecutive thyroid FNAB—in which BRAF status had been explored—we considered the first 700 cases judged to be malignant in the final pathological report. The study was approved by the local ethics committee, and all the patients gave written informed consent to their thyroid tissues being used for research purposes.
FNAB cannulas with thyrocyte material were rinsed and re-suspended in 3 mL physiological saline solution, followed by genomic DNA isolation (QIAamp DNA Micro Kit; Qiagen), using the manufacturer's protocol. DNA was extracted from frozen tissues after surgery, using the DNeasy Blood and Tissue Handbook (Qiagen). All DNA from FNAB samples and all DNA extracted from tissues were examined by direct sequencing, as described elsewhere (17). The same frozen tissues were used for protein extraction. Nucleic acids were quantified with NanoDrop 1000 (Thermo Scientific).
Mutation analysis
Exon 15 of the BRAF gene (NM_004333.4) was amplified and then sequenced on an ABI PRISM 3130 genetic analyzer (Applied Biosystems). The polymerase chain reaction (PCR) protocol and primers are described elsewhere (18). Exons 2 and 3 of H-RAS (NM_005343.2), N-RAS (NM_002524.3), K-RAS (NM_033360.2); exons 5, 7, and 8 of PTEN (NM_000314.4); and exons 9 and 20 of PIK3CA (NM_006218.2) genes were analyzed only for samples with rare BRAF variants, as described previously (13).
Mutant allele-specific PCR amplification
Mutant allele-specific PCR amplification was done essentially as described elsewhere (18). A forward primer flanking the sequence of exon 15 (5′-TAG GTG ATT TTG GTC TAG CTA CAG T-3′) was used as a positive control to amplify wild-type as well as mutant BRAF. A second primer was designed with two bases substituted at the 5′-end (GGT GAT TTT GGT CTA GCT ACA AA-3′) to amplify only the mutant BRAF gene. The sequence of the reverse primer was 5′-GGC CAA AAT TTA ATC AGT GGA-3′.
The PCRs were obtained separately in a Veriti Thermal Cycler (Applied Biosystems), including an initial denaturation for 2 minutes at 94°C and subsequent denaturation for 30 seconds at 94°C, annealing for 30 seconds at 58°C, and extension for 30 seconds at 72°C. All samples were reexamined for the BRAF mutation at least three times.
Western blot analysis
Samples were collected after surgery, frozen immediately in liquid nitrogen, and stored at −80°C.
Immunoblot analysis was performed on normal and cancerous frozen thyroid tissue segments, as described elsewhere (19). Briefly, proteins were separated by sodium dodecyl sulfate polyacrylamide gel electrophoresis, electroblotted onto nitrocellulose membranes, and saturated in 5% nonfat dry milk. Membranes were incubated overnight with primary antibodies and then incubated with secondary ones (Jackson ImmunoResearch, Europe). Immunoreactivity was detected with Pierce ECL Western Blotting substrate (Thermo Scientific). The primary antibodies were anti-Erk1/2, anti-phospho-Erk1/2 (Thr202/Tyr204), anti-MEK1/2, anti-phospho-MEK1/2 (Ser221), anti-Akt, anti-phospho-Akt (Ser473), anti-p70S6 kinase, and anti-phospho-p70S6 kinase (Thr389), all from Cell Signaling (1:1000), and anti-β-Actin, from Sigma-Aldrich (1:5000). Films were scanned and band intensity was quantified with ImageJ software 1.44p. All experiments were performed in triplicate.
Immunofluorescence analysis
Immunofluorescence analysis was performed on paraffin-embedded sections, as described elsewhere (20). Pathological and normal sections were deparaffinized in xylene, rehydrated in graded ethanol series (100–70%), and demasked by sub-boiling in citrate buffer (pH 6.0). Slides were incubated overnight with anti-phospho-Erk1/2 (1:200) and anti-phospho-Akt (1:25) from Cell Signaling. Antirabbit secondary antibody Alexa 594 (1:200; Life Technologies) was used. Sections were stained with DAPI for 5 minutes and then washed. Images obtained with the epifluorescence microscope were acquired with a digital system and processed with Corel Photo Paint×3. Hematoxylin and eosin completed the staining procedure after immunofluorescence. Sections were analyzed independently by three experts (R.P., M.R., and G.P.).
In silico analysis
Analysis of B-raf crystal structures
The RCSB Protein Data Bank (21) contains 20 entries related to the human tyrosine kinase sequence of B-Raf known as P15056 (22). All the crystal structures within the Protein Data Bank were cocrystallized with B-Raf inhibitors. None of them represented an apo-form crystal structure. All the crystal structures were superimposed with Molecular Operating Environment 2010.10 (23) and analyzed considering the geometrical orientation of the structural determinants (P-loop, αC-helix, DFG motif, A-loop).
Generation of B-raf T599dup, V600_K601>E, V600_S605>D models based on 3Q4C and on 3SKC
A B-Raf T599dup (case 1) 3D model based on the 3Q4C Protein Data Bank was chosen as the template using the SwissModel (24) workspace. 3Q4C contains the crystal structure of the B-Raf tyrosine kinase with DFG “in” and E501 “in” conformation. The A-loop was modeled from within the SwissModel procedure in the “active” geometrical conformation and further refined with a loop search with Molecular Operating Environment 2010.10. The 3D model obtained was parameterized with AMBER FF99SB force-field (25) and subsequently protonated with Protonate3D (26) within Molecular Operating Environment 2010.10. The B-raf T599dup was also submitted to 3 minimization cycles considering T599dup alone, the 4.5 Å environments around the insertion, and the global minimization. This procedure was further applied to generate the B-Raf models based on the 3SKC template representing the mutations of case 3 (V600_K601>E) and case 4 (V600_S605>D).
Molecular dynamics
The protonation state of the 2 protein models was assessed using the Protonate3D and Protonate within AmberTools 1.5 (25). The tLeap was subsequently used and the 4 protein models were parameterized by Amber FF99SB. The protein models were solvated in TIP3P water boxes, adding counterions (Na+) to ensure the neutrality of the molecular systems. ClickMD (26,27) was used as an MD platform to minimize (100,000-step, conjugated-gradient method) and equilibrate (0.5 ns, alpha carbon positional restraints) the production phase (100 ns moles–volume–temperature [NVT], P=1 atm, T=300 K) of the molecular systems using the 100,000 conjugated gradient method. ACEMD v2325 (28) was used as the MD engine on nVidia GTX580. VMD 1.9.1 (29), RMSD Trajectory Tools 2.01 (30), RainbowRMSD (27), and NRGPLOT (27) were used to analyze the trajectories produced.
Statistical analysis
Statistical analyses were performed using GraphPad Prism 4 software (GraphPad Software, Inc.). Groups were compared using the Kruskal–Wallis analysis followed by the Dunn's post-test. p<0.05 was considered statistically significant. Data were presented as mean±standard error of the mean.
Results
BRAF mutation analysis
Over the past 2 years, a total of 2131 consecutive cases of thyroid FNAB were collected from patients attending the Endocrinology Unit of the University of Padua and subjected to BRAF mutation analysis. Seven hundred of the 2131 FNAB samples were carcinomas: 644 were PTC, 22 were anaplastic thyroid carcinomas, and 34 were FTC. BRAF sequent variants were found in 54% (347/644) of patients with PTC and in 1 of the 34 with FTC.
Five rare genetic variants were found for a total of 11 malignant cases (1 FTC and 10 PTC).
No BRAF alterations were found in noncancerous FNAB lesions. The BRAF mutation status of the FNAB samples was the same as in the frozen tissues; all samples were re-examined and the BRAF mutations were confirmed by mutant allele-specific PCR amplification.
The classic c.1799T>A (BRAF V600E ) mutation was found in 96.8% (336/347). The 5 rare BRAF alterations were (i) an in-frame c.1795_1797dupACA (p.T599dup) insertion in 2 patients (one with a follicular and the other with a classical variant PTC; Fig. 1A); (ii) a point c.1801A>G mutation (p.K601E) in 4 patients (1 with a poorly differentiated follicular carcinoma and 3 with a follicular variant PTC; Fig. 1B); (iii) a triplet deletion c.1799_1801delTGA (p.V600_K601>E) with no frame-shift in 3 patients (the first with a hobnail, the second with a tall cell, and the third with a classical variant PTC; Fig. 1C); (iv) a double mutation producing a predictive in-frame deletion/insertion c.1799_1814>A (p.V600_S605>D) presented in a classical variant PTC (Fig. 1D); and (v) a predictive in-frame deletion/insertion c.1798_1810delinsA (p.V600_W604>R) never previously described in the literature (in a classical variant PTC; Fig. 1G).
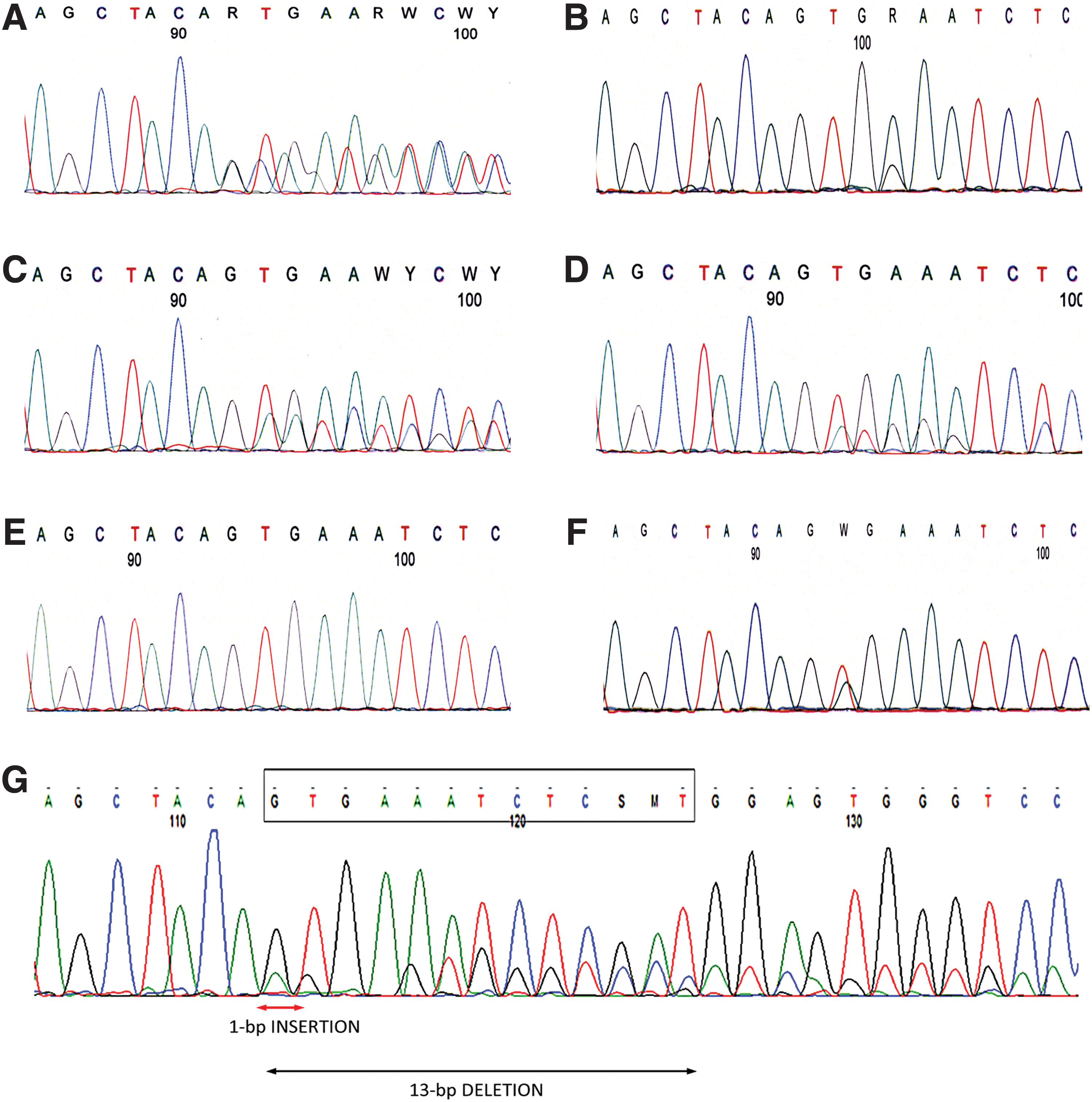
Representative sequences of electropherograms obtained in the 5 rare BRAF alterations:
A point mutation (p.E545A) in exon 9 of the PI3KCA gene was demonstrated in the patient with a poorly differentiated follicular carcinoma simultaneously carrying the previously mentioned c.1801A>G (p.K601E) BRAF mutation (data not shown) (13).
The clinical characteristics of the patients with rare BRAF variants are shown in Table 1. To date, with a median follow-up of 28 months (min. 2, max. 59), 7 patients were “disease-free,” 1 had “persistent” disease, and 3 had too short a follow-up to establish their disease status.
TNM classification, using AJCC Cancer Staging Manual, 7th edition.
Not sufficient follow-up to define disease status.
CV-PTC, classic variant papillary thyroid cancer; F, female; FTC, follicular thyroid cancer; FV-PTC, follicular variant papillary thyroid cancer; HNV-PTC, hobnail variant papillary thyroid cancer; M, male; TCV-PTC, tall cell variant papillary thyroid cancer.
No other mutations in H-Ras, K-Ras, N-Ras, and PTEN were found in the above cases.
Western blot analysis
The first rare BRAF variant, an in-frame c.1795_1797dupACA (p.T599dup) insertion, produced a nearly uniform pattern in all the antibodies analyzed except for the p70S6k and p-p70S6k ones (no reactivity; Fig. 2). The second rare BRAF variant, the point mutation c.1801A>G (p.K601E), produced an interesting pattern: no Akt, p-Akt, MEK1/2, or p-MEK1/2 expression; abnormal p70S6k reactivity; and a slight activation of Erk1/2. The third rare BRAF variant carrying the c.1799_1801delTGA (p.V600_K601>E) alteration showed an almost complete activation of all the kinases recognized by the antibodies used. The fourth rare BRAF variant, carrying the c.1799_1814>A (p.V600_S605>D) alteration, revealed a weak activation of all the components studied with the exception of p-Erk1/2.
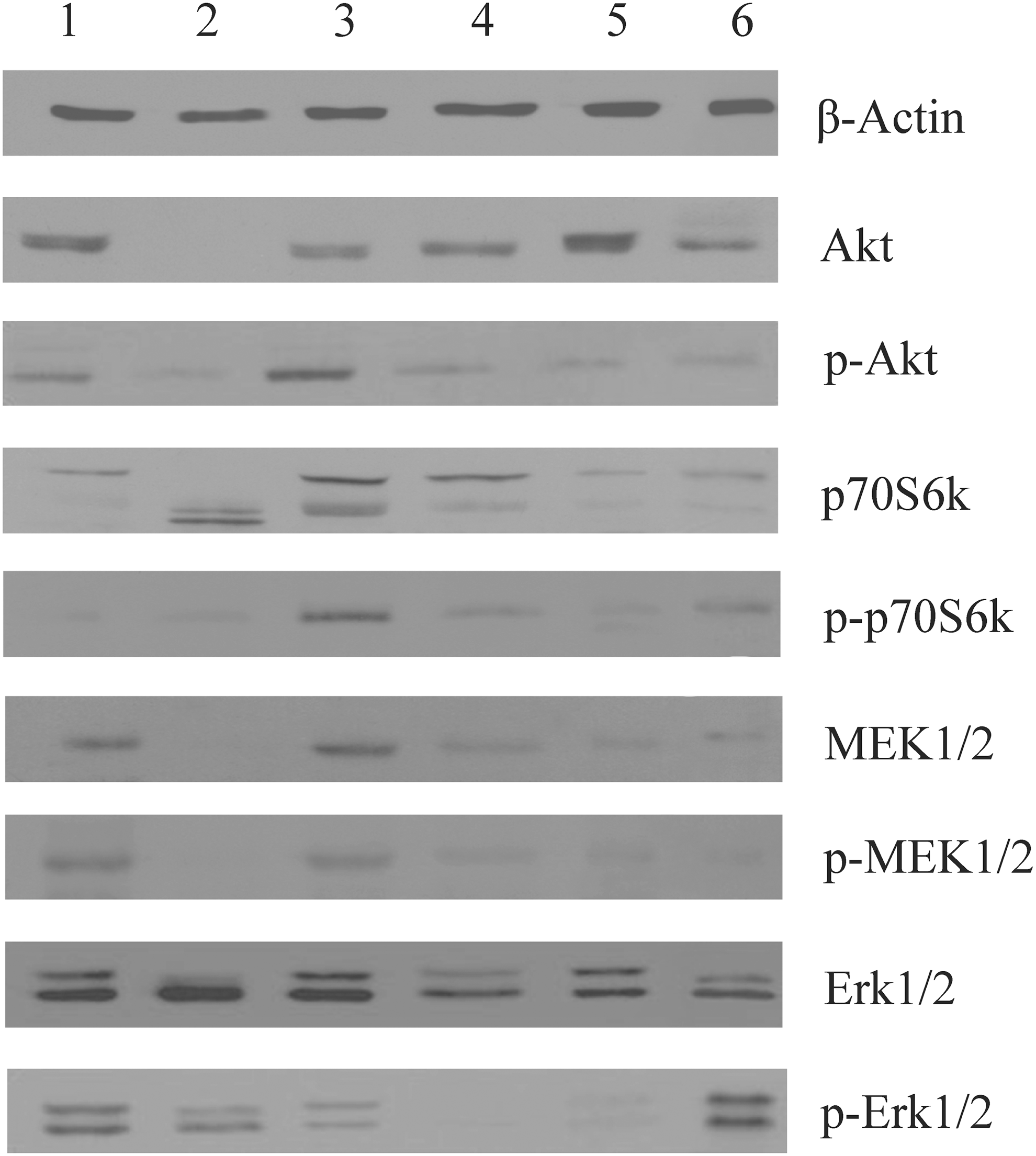
Representative Western blots of six thyroid tissues. Case 1 concerns a patient carrying the insertion c.1795_1797dupACA (p.T599dup); Case 2 is a patient with the point mutation c.1801A>G (p.K601E); Case 3 is a patient with the deletion c.1799_1801delTGA (p.V600_K601>E); Case 4 is a patient carrying the in-frame deletion/insertion c.1799_1814>A (p.V600_S605>D); Case 5 is normal thyroid tissue obtained from a cadaver; and Case 6 is a patient with the classic BRAF V600E mutation c.1799T>A. These profiles are representative of three independent experiments.
Four PTC tissues without any BRAF mutations were analyzed as well: they reacted to all the antibodies studied in the same way as normal thyroid tissue (data not shown).
Band quantification statistical analyses are shown in Supplementary Table S1 (Supplementary Data are available online at
Immunofluorescence analysis
Immunofluorescence was performed on the same tissues as above. Five thyroid neoplastic tissues were analyzed (Supplementary Fig. S1), and one normal sample was used as a control (not shown). The p-Erk1/2 and p-Akt reactivity was comparable with the findings of Western blot for all the samples.
In silico analysis
B-Raf mutation models (T599dup, V600_K601>E, V600_S605>D) based on 3SKC (“inactive” geometrical conformation) and 3Q4C (“active” geometrical conformation) templates were generated and 6 models were obtained. Both the “active” and the “inactive” conformations were explored, also examining how the mutations on the two extreme conformations affected the molecular level.
Visual inspection of the B-Raf T599ins model based on 3Q4C
T599ins was located in the modeled A-loop of the B-Raf protein and its side chain was surrounded by R575 (catalytic loop) and K601. The side chain of R575 made a H-bond with the backbone of L597 (A-loop). One of the two γ-hydrogens of R575 pointed toward the phenyl ring of Y634 (C-lobe) making a cation–π interaction. The A-loop conformation was further stabilized by the K601 side chain, which made a H-bond with the backbone of Y634. The S602 side chain was solvent exposed and its conformation was suitable for phosphorylation. The T599 side chain was closed to the αC-helix, which did not allow a suitable conformation for its phosphorylation (Supplementary Fig. S2).
Molecular dynamics of B-Raf models
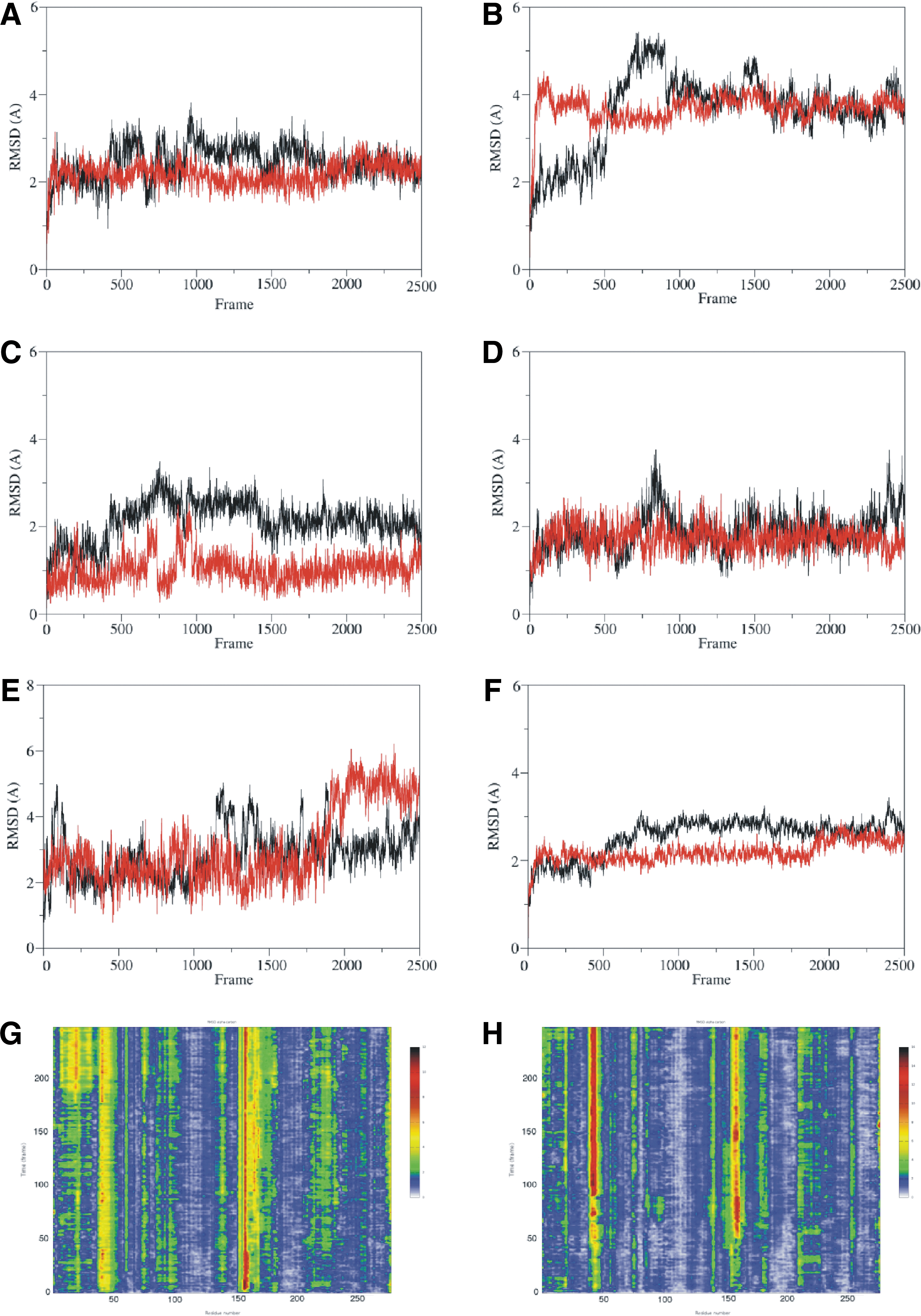
We performed a 100 ns NVT MD simulation of both B-Raf wild-type and T599dup models based on the 3Q4C template. The root mean square deviation (RMSD) behavior during the dynamics of each structural determinant (αC-helix [Fig. 3A], A-loop [Fig. 3B], DFG-motif [Fig. 3C], hinge region [Fig. 3D], and P-loop [Fig. 3E] of the kinase domain and of the whole catalytic domain [Fig. 3F]) was analyzed. The previously mentioned RMSD results were further summarized in the RainbowRMSD representations in Figure 3G and H, where the x-axis represents the α-carbons of the amino acids of the molecular system, and the y-axis the time scale of the MD simulation (from 0 to 100 ns). The RMSD values during the MD run are associated with a color scale from blue (low RMSD values) to red (high RMSD values). The results of B-Raf wild type are outlined in black, and those of B-Raf T599dup are shown in red. Analyzing the RMSD changes over time indicated that T599dup stabilized the active geometrical conformation to a greater extent than the wild type. The active geometrical conformation is associated with the productive catalytic state. These results suggest that an in vitro study of B-Raf T599dup might depict an increase in kinase activity and an activation of the Erk1/2 pathway in the cell.
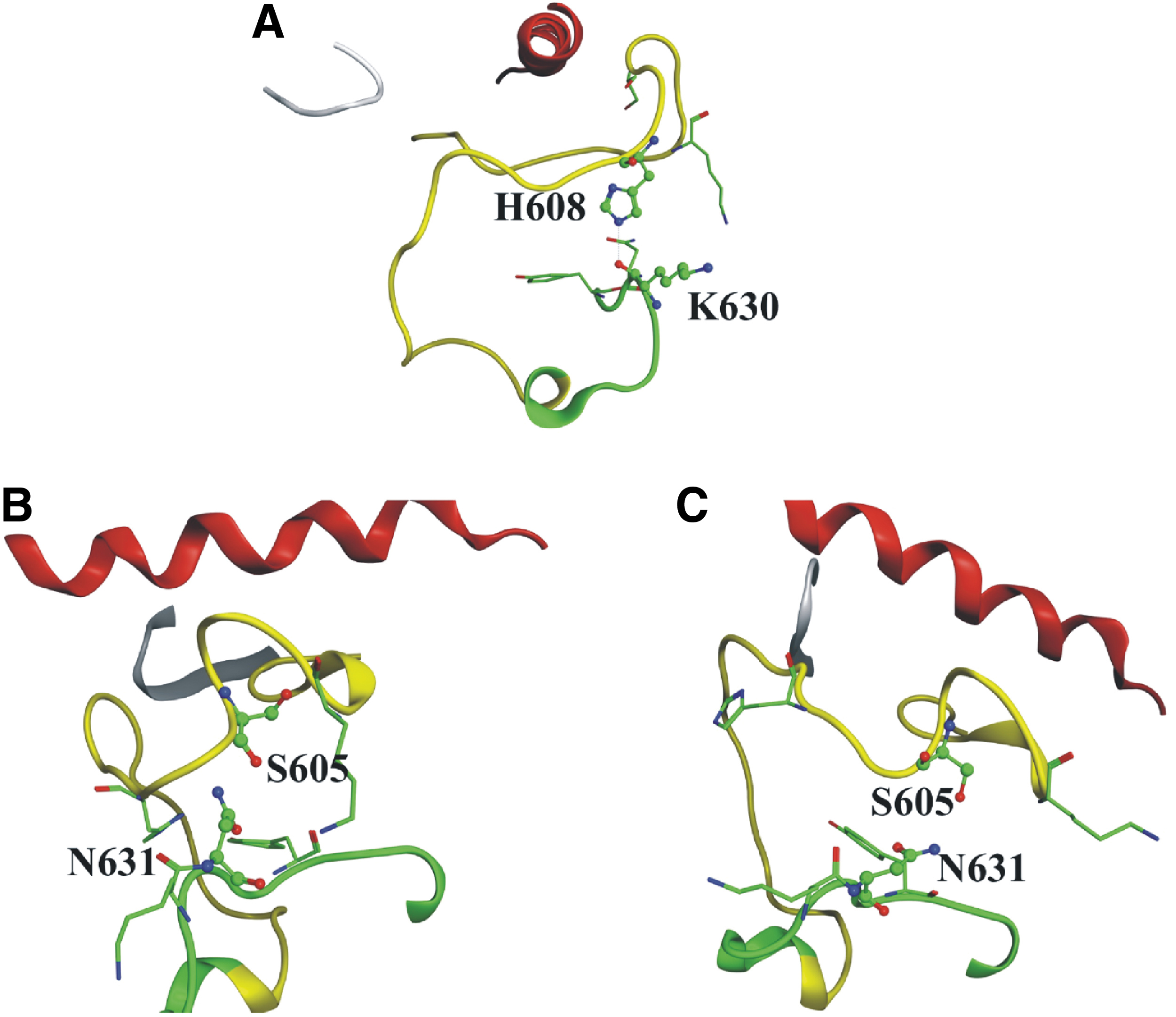
The conformation of the A-loop was stabilized by polar interactions with the C-lobe. The previously described K601–Y634 interaction was substituted by H608–K630 (Fig. 4A) and S605–N631 (Fig. 4B, C) interactions during a 100 ns NVT large-scale MD simulation.
The MD trajectories of B-Raf wild type and B-Raf V600_K601>E, both 3Q4C-based, were visually inspected and compared. We compared the RMSD over time analysis of the A-loop and the DFG-motif of the two active geometrical conformation models. The RMSD values of the A-loop of B-Raf V600_K601>E were similar to those of the B-Raf T599dup, which differed substantially from those of the wild-type kinase domain (Fig. 5A). These results were consistent with the Western blot data and suggest that V600_K601>E causes a stabilization of the “open” A-loop conformation. In particular, B-Raf V600_K601>E during the 100 ns NVT MD simulation showed the formation of a H-bond between the A-loop and the C-lobe involving the backbone of S571 and side chain of S607 (Fig. 5B, C). The stability of this interaction is confirmed by the distance analysis depicted in Figure 5D.

We visually inspected and compared the MD trajectories of B-Raf wild type and B-Raf V600_S605>D, both 3Q4C based.
The RMSD analysis of the kinase domain over time exhibited lower RMSD values of V600_S605>D than the wild-type model (Supplementary Fig. S3A). This was also true for the A-loop and the hinge region RMSD of V600_S605>D graphs, and suggests that the mutants were more stable than the wild type (Supplementary Fig. S3B, C). The V600_S605>D A-loop RMSD over time showed a pattern similar to the one obtained for the T599dup and V600_K601>E models. Heat maps and Rainbow RMSD supported these results.
Discussion
In this study, we investigated the prevalence, tumorigenic role, and biochemical implications of rare genetic BRAF variants in a large cohort of patients.
Ranging between 36% and 69% of PTC in other studies, the frequency of the BRAF V600E mutation (52%) found in our cohort is consistent with previous reports (2,3).
Mutation analysis revealed 5 rare BRAF variants, globally demonstrated in 10 PTC and in 1 FTC case with an overall prevalence of 1.6% (10 out of 644) of PTC and 1.6% (11/700) of all thyroid cancers in our series.
It is well known that the p.V600E and p.K601E alterations are the primary contributors to the constitutive activation of BRAF mutations in PTC (10,11). As a result of these genetic modifications, B-Raf triggers a downstream signaling cascade through the phosphorylation of diverse serine-threonine kinases, leading to the final activation of Erk1/2 kinases before they go on to influence gene transcription and expression. The p.K601E BRAF mutation is more frequent in the follicular variant of PTC, in which it has been identified in about 7% of cases and, as we and others previously demonstrated, it should be included (although it occurs very infrequently) in the spectrum of genetic alterations seen in follicular adenoma and carcinoma too (13,31). As already described in a case of FTC in a recent work by our group (13), Western blot analysis showed that Erk1/2 was phosphorylated in the presence of the BRAF K601E mutation, thus supporting the hypothesis that the MAPK signaling pathway was activated. Interestingly, this mutation was associated with a particular reactivity pattern for p70S6 kinase characterized by bands at a different molecular weight when compared with the control. It may be that the RPS6KB1 gene, which encodes for p70S6k, has some genetic alterations that give rise to a reduced protein, without modifying the recognition site for the monoclonal antibody. An alternative explanation could be that a specific p70S6k isoform exists with a different molecular weight. It is not clear whether this alteration relates to a modulation in the PI3k/Akt pathway: further studies are needed to elucidate these findings.
The p.T599dup alteration already described in the literature (32) positively modified the Western blot reactivity of Erk1/2 and p-Erk1/2, supporting the hypothesis that the MAPK pathway is activated, and consistent with the findings of Eisenhardt and collaborators, who reported activation of this pathway in HEK 293 cells expressing B-Raf (33). Akt and p-Akt in the PI3k/Akt pathway were also slightly activated, suggesting that a cross-interaction between the MAPK and PI3k/Akt pathways might be taking place, as previously suggested (14). Large-scale MD simulations (100 ns NVT conditions) and RMSD analysis of the B-Raf T599dup model based on the “active” geometrical conformation suggest that the T599dup mutation plays a part in stabilizing the “active” geometrical conformation of the kinase domain. This conformational state is associated with a productive one, enabling the kinase to phosphorylate the B-Raf substrates detected by Western blot analysis.
The triplet deletion (p.V600_K601>E), involving 2 targeted codons in the previously reported 1799T codon 600, and 1801A codon 601 positions, was first described in a solid variant of PTC (34) and later associated with a lymph node metastasis of PTC (11). Western blot reactivity was very similar to that of the p.T599dup alteration. Our data show that key components of the signaling pathway downstream from B-Raf (MEK1/2, p-MEK1/2, Erk1/2 and p-Erk1/2) are activated. The 100 ns NVT MD simulation showed the formation of a H-bond between the C-lobe and the A-loop involving the S571 and S607, respectively. This interaction, which helps to keep the A-loop in the “open” conformation, linked to the productive state of the kinase, was not seen in the wild-type model. This deletion was functionally characterized by Hou and collaborators (35), who stably transfected NIH3T3 cells with the p.V600_K601>E construct, demonstrating an anchorage-independent colony-forming ability of the transfected cells, similar to the one observed with the BRAF V600E construct, and an increase in the phosphorylation of MEK and ERK in cells expressing BRAFV600E and BRAF V600_K601>E. These data suggested that this mutation is capable of constitutively activating the B-Raf kinase and the MAP kinase pathway, similar to the classical BRAFV600E mutation (35). This BRAF mutation was reported in a recent work as a very rare genetic event (1/500 PTC examined histologically) (36). Fairly consistent with this conclusion, we confirm that the p.V600_K601>E BRAF variant is quite a rare genetic event in the development of PTC, occurring in 0.5% of cases (3 out of 644 PTC); in our series, one case interestingly showed prominent hobnail features—a recently described rare variant of PTC with a particularly aggressive behavior (37).
The deletion/insertion (p.V600_S605>D) BRAF variant spanned an additional five amino acids and ended with the insertion of an aspartic acid. Acidic residues [Asp (D) or Glu (E)] substituted at this position may serve as phosphomimetics for two nearby phosphorylation sites (T598 and S601) that are crucial to kinase activation (6,38). Judging from Western blot analysis, this genetic event showed a mild band pattern for all the antibodies screened, while there was no p-Erk1/2 reactivity. Only one work in the literature has described this BRAF mutation in a case of uveal melanoma (39). The alteration did not seem to produce any MAPK pathway activation through the phosphorylation of Erk1/2. We can assume that other means could be used by the cell machinery to trigger proliferation signals in this particular case of PTC. The formation of the I572–T598 backbone interaction that stabilizes the “active” geometrical conformation was found during the 100 ns NVT MD simulation of B-Raf V600_S605>D (Supplementary Fig. S3D).
During the submission of the present article, we found a novel deletion/insertion c.1798_1810delinsA (p.V600_W604>R) of the BRAF gene in a patient with a classical variant PTC. This mutation was a heterozygous 13 bp deletion with 1 bp insertion, resulting in a complex amino acid change, with the substitution of valine 600 by arginine and the deletion of lysine 601, serine 602, arginine 603, and tryptophan 604. This genetic event has never been reported before in human cancer, and further studies are needed to elucidate its functional consequences on BRAF activation status.
To conclude, we examined the prevalence of rare BRAF alterations in thyroid carcinomas and the biochemical characterization of these genetic modifications, which revealed a complex picture that appears to be consistent with the hypothesis of a downstream activation of B-Raf signaling.
All BRAF mutations were found clustered around the codon V600 in the binding pocket named A-loop (Fig. 6), suggesting that this region—which is crucial to activation of the enzymatic activity of the B-Raf protein—is highly susceptible to alterations and changes from an electrically neutral to a charged status. For some rare BRAF variants, Western blot also revealed a simultaneous activation of both the MAPK and the PI3k/Akt pathways, suggesting that the two signaling cascades might cooperate to promote cell transformation under particular genetic conditions.

Conformation of B-raf wild type, highlighting the location of rare mutations in the binding pocket named A-loop. Color images available online at
Clinically, these genetic events are not associated with an aggressive biological behavior in PTC, although they are sometimes associated with aggressive PTC histotypes such as the tall-cell or hobnail variants.
Footnotes
Acknowledgments
We thank Linda Inverso and Frances Coburn for text editing. The molecular modeling work coordinated by S.M. was carried out with financial support from the University of Padua, Padua, Italy; the Italian Ministry for University and Research, Rome, Italy; and the Associazione per l'Emofilia e le Coagulopatie delle Tre Venezie, Padua, Italy. We also thank the Associazione Italiana per la Ricerca Oncologica di Base (AIROB), Padua, Italy.
Author Disclosure Statement
The authors have no competing financial interests to declare.