
Abstract
Background:
The impact of excessive iodine intake on the development of autoimmune thyroiditis (AIT) is still under debate. Transgenic, antibody-devoid TAZ10 mice spontaneously develop AIT due to autoreactive thyroperoxidase-specific T cells. In this model, development of AIT is determined by a T cell infiltration of the thyroid gland leading to an elevation of serum thyrotropin (TSH) levels and significant weight gain. In the present study we investigated the impact of moderate and high iodine supplementation on the course of disease in these mice, which are immunologically prone to AIT.
Methods:
In addition to normal nutrition, mice were supplemented for 20 weeks with 2.5 μg versus 5 μg iodine per milliliter drinking water, which corresponds to a human daily iodine supplementation of 150 μg, 315 μg, and 615 μg iodine. AIT-defining parameters (weight gain, elevation of serum TSH levels, cellular infiltration of the thyroid) and immunologic effects were analyzed.
Results:
No significant differences were displayed when comparing weight and serum TSH levels in the iodine-supplemented versus control groups. Increased thyroid infiltrates with CD8+ T cells were detected by fluorescein-activated cell sorter (FACS) and immunofluorescence staining in mice supplemented with elevated iodine amounts (315 μg and 615 μg iodine per day, respectively). Immunologic monitoring revealed selective changes in immune cell frequencies (CD8+ and regulatory T cells, natural killer [NK] cells) and cytokine production (interferon-γ, interleukin-1α, and interleukin-17), however, without affecting the overall immune balance.
Conclusion:
Our results demonstrate that elevated iodine supplementation has no physical impact on the course of disease in transgenic, antibody-devoid TAZ10 mice, which are immunologically prone to AIT.
Introduction
H
Various studies have been performed worldwide analyzing the effect of excessive iodine intake on the prevalence of autoimmune thyroiditis (AIT). The majority of publications demonstrated a significant association between excessive iodine intake and AIT (9 –13). Other studies in restricted geographical areas or population groups, however, could not confirm these results (14 –20). It is proposed that increased iodine intake operates as a potent trigger for AIT, however preferentially in genetically susceptible individuals (6). Until now, there have been no systematic data available regarding the cellular mechanisms and the physical outcome in T helper 1 dominant animal models supplemented with increased amounts of iodine. Therefore, we investigated the impact of moderate and high iodine supplementation on the disease course in TAZ10 mice, which are immunologically prone to AIT. TAZ10 mice are transgenic mice, with a CBA background, expressing a human T-cell receptor derived from a thyroid-infiltrating T cell clone of a patient with AIT and furthermore specific for a cryptic TPO epitope (21). This particular human T-cell receptor recognizes two different epitopes within the TPO535–551 peptide: the highly stimulatory TPO536–547 and TPO537–548, which are designated as a naturally occurring antagonistic epitopes, potentially inducing anergy (22,23). Nevertheless, this mouse strain exhibits an activated T cell phenotype in vivo and spontaneously develops AIT, which is equivalent to the human disease, determined by significant weight gain, elevation of serum thyrotropin (TSH) levels and cellular infiltration of the thyroid. Having been backcrossed to Rag1 −/− strains, these mice are devoid of functional antibodies. Therefore, it is to be concluded that these mice develop AIT due to spontaneous cellular self-reactivity in the absence of autoantibodies (21). So, we specifically emphasize the impact of a moderate and high iodine supplementation on cellular mechanisms and physical outcome in mice, which are immunologically prone to AIT by spontaneous cellular self-reactivity.
Methods
Mice
Transgenic TAZ10 mice have been kindly provided by Dr. S. Quaratino from the Cancer Research UK Oncology Unit at the University of Southampton (Southampton, United Kingdom). TAZ10 mice are transgenic mice expressing a human T-cell receptor-specific for a cryptic TPO epitope. This mouse strain exhibits an activated T cell phenotype and spontaneously develops AIT. Having been backcrossed to Rag1 −/− strains, these mice are devoid of functional antibodies. All experiments were conducted in accordance with the Animal Experimental Ethics Committee Guidelines and with the permission from the Regional Council North Rhine-Westphalia/Germany.
Treatment with iodine
Mice were bred in the animal facility of the Heinrich Heine University (Duesseldorf, Germany), kept under specific pathogen-free conditions, and received drinking water and complete rodent diet (ssniff Spezialdiäten GmbH, Soest, Germany) including 2.2 mg iodine per kilogram feed ad libitum to ensure a daily iodine intake of 750 ng per mouse, equal to a human daily supplementation of 150 μg iodine. At the time of initiation of the experiments all mice were 4 weeks old. Male as well as female mice were included in the study (51 male and 50 female mice).
Control mice (devoid of iodine in drinking water, n=46, 30 male and 16 female mice) took in 750 ng iodine per day by eating complete rodent diet. Experimental groups were additionally supplemented with 2.5 μg versus 5 μg iodine per milliliter drinking water for 19 weeks. Assuming that a mouse drinks ∼6 mL of water per day, experimental groups ingested 15.75 μg (n=27, 10 male and 17 female mice) and 30.75 μg (n=28, 11 male and 17 female mice) iodine per day, respectively. These iodine amounts correspond to a daily supplementation in humans of 150 μg, 315 μg, and 615 μg iodine. These three groups are named in the following as “normal,” “moderate,” and “high” iodine supplemented mouse groups. AIT-defining parameters, including weight gain, elevation of serum TSH levels, and cellular infiltration of the thyroid were analyzed after 19 weeks and compared to the untreated control mice receiving normal iodine supplementation.
Dissection and size measurement of the thyroid gland
Thyroid glands, esophagus, trachea, and surrounding tissue were removed from the mice. Using a Stereozoom microscope (Leica Zoom 2000, Leica Microsystems GmbH, Wetzlar, Germany) the thyroid gland was dissected from the other organs. Photographs of dissected thyroids were then taken. Based on the photographs, the thyroid size was determined by measuring the circumference (mm2) and setting in relation to the mice's body weight (g). Relative thyroid gland size is expressed as mm2/g.
Phenotypical analysis by flow cytometry
Single cell preparations were generated from spleens and thyroids as follows: thyroid glands were dissected as described above. Spleen and thyroid gland were cut several times with a scalpel, placed on a 100 μm filter (BD Biosciences), and forced through it to obtain a single cell suspension. Afterwards cells were incubated in 2-mL erythrocyte lysis buffer (Qiagen, Hilden, Germany) for 15 minutes at room temperature. Subsequent to washing in MACS buffer (Miltenyi Biotec, Bergisch Gladbach, Germany), the samples were blocked by FC blocking reagent (BD Biosciences, San Jose, CA) and labeled with fluorochrome-tagged antibodies on ice for 30 minutes. After washing the samples thoroughly, they were analyzed on a FACS Calibur (BD Biosciences) equipped with BD-CellQuest-Pro software. Monoclonal anti-mouse antibodies were purchased from BD Biosciences (alternative manufacturer as indicated), were conjugated to FITC, PE, Pe-Cy7, PerCP, or APC and targeted the following antigens (by clones): CD107a (1D4B), CD335 (29A1.4), CD45 (30F11), CD8 (53-6.7), TCR (GL-3; ImmunoTools, Friesoythe, Germany), CD4 (H129.19), CD49b (HMα2), CD25 (PC61), and NK1.1 (PK136). Appropriate rat, mouse, or hamster IgG isotype controls were used to determine the levels of background staining. Interleukin (IL)-1 (ALF-161), IL-17 (eBio17B7, eBioscience, Frankfurt, Germany), interferon (IFN)-γ (XMG1.2, eBioscience), and FOXP3 (FJK-16s, eBioscience) were stained intracellularly with the BD Cytofix/Cytoperm™ Kit according to the manufacturing protocol.
Immunofluorescence
Thyroid tissue was cryosectioned into 7 μm slices and placed on an object slide. The object slides were then dried for 10 minutes at room temperature before and subsequent to bathing in acetone for 10 minutes. Between each of the following steps, the object slides were washed thoroughly in TBS. After blocking the slices with 5% swine serum (DAKO, Glostrup, Denmark) for 1 hour at room temperature, they were incubated at 4°C with anti-CD8 (clone 53-6.7, BP Pharmingen, Heidelberg, Germany) and anti-CD4 (clone H129.19, BD Pharmingen) diluted in phosphate-buffered saline (PBS; 1:100) overnight. Secondary antibodies (goat-anti rat, rabbit-anti rat) labeled with Alexa-488 and Alexa-594 (dilution 1:1500; Invitrogen, Carlsbad, CA) were used. Slides were mounted with ProLong Gold antifade reagent (Invitrogen) with counterstaining of the cell nuclei by 4,6-diamidino-2-phenylindol. Fluorescence signals were visualized by a Nikon eclipse TE 300 fluorescence microscope with a Nikon DXM 1200 digital camera (Nikon Instruments Inc., Melville, NY).
Serum-TSH ELISA
Mouse serum was separated by centrifuging whole blood for 10 minutes at 13,000 rpm. The serum-TSH enzyme-linked immunosorbent assay (ELISA) was performed with the rodent TSH ELISA test kit (Endocrine Technologies, Newark, CA) as described in the manufacturing protocol.
Statistical analysis
For data showing a Gaussian distribution, we performed the unpaired Student's t test (data are expressed as mean±standard deviation); for non-normally distributed data, the Mann-Whitney test was used (data are expressed as median [25th, 75th percentile]) to compare data from control group versus mice with moderate iodine intake and control group versus mice with high iodine intake. Analyses were performed using the Prism computer software (GraphPad Software, Inc., La Jolla, CA). p Values<0.05 were considered as significant.
Results
Impact of enhanced iodine supplementation on clinical hallmarks of AIT
In terms of weight, we did not observe significant differences between the two iodine supplemented groups and the control group (normal iodine intake, 29.4±4.1 g; moderate iodine intake, 31.4±6.1 g; high iodine intake, 30.4±5.7 g; Fig. 1A).
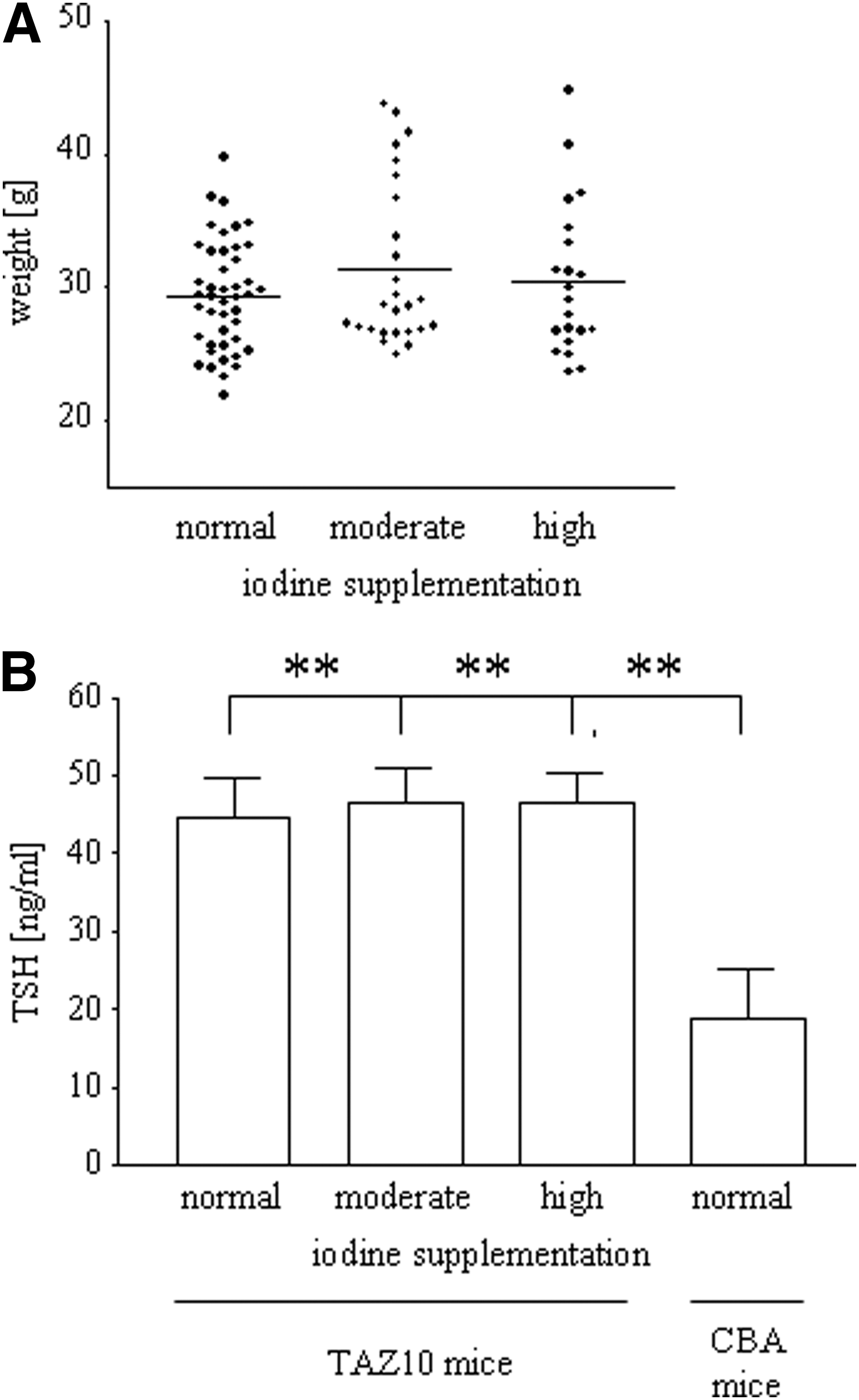
Enhanced iodine supplementation has no impact on weight of mice or serum thyrotropin (TSH) levels.
Compared to wild-type CBA mice, serum TSH levels were significantly elevated in all TAZ10 mice irrespective of the iodine supplementation (p<0.01). In contrast, however, there were no significant differences between the groups of mice treated with different amounts of iodine (normal iodine intake, 44.6±20.1 ng/mL; moderate iodine intake, 46.6±17.7 ng/mL; high iodine intake, 46.6±16.8 ng/mL; CBA mice, 18.8±23.9 ng/mL; Fig. 1B).
A significantly increased infiltration of the thyroid by CD8+ T cells could be demonstrated by FACS and immunofluorescence staining in the mice supplemented with moderate iodine (0.4±0.3% CD45+/CD8+ cells, p=0.0244) and high iodine (0.5±0.3% CD45+/CD8+ cells, p=0.0034) compared to the control mice (normal iodine intake, 0.1±0.2% CD45+/CD8+ cells; Fig. 2A, B). The thyroid gland size was only significantly augmented in those mice treated with high iodine levels (normal iodine intake, 0.21±0.04 mm2/g; moderate iodine intake, 0.26±0.09 mm2/g; high iodine intake, 0.35±0.09 mm2/g, p<0.0001; Fig. 2C), which correlates with the increased number of infiltrating T cells into the thyroid gland.
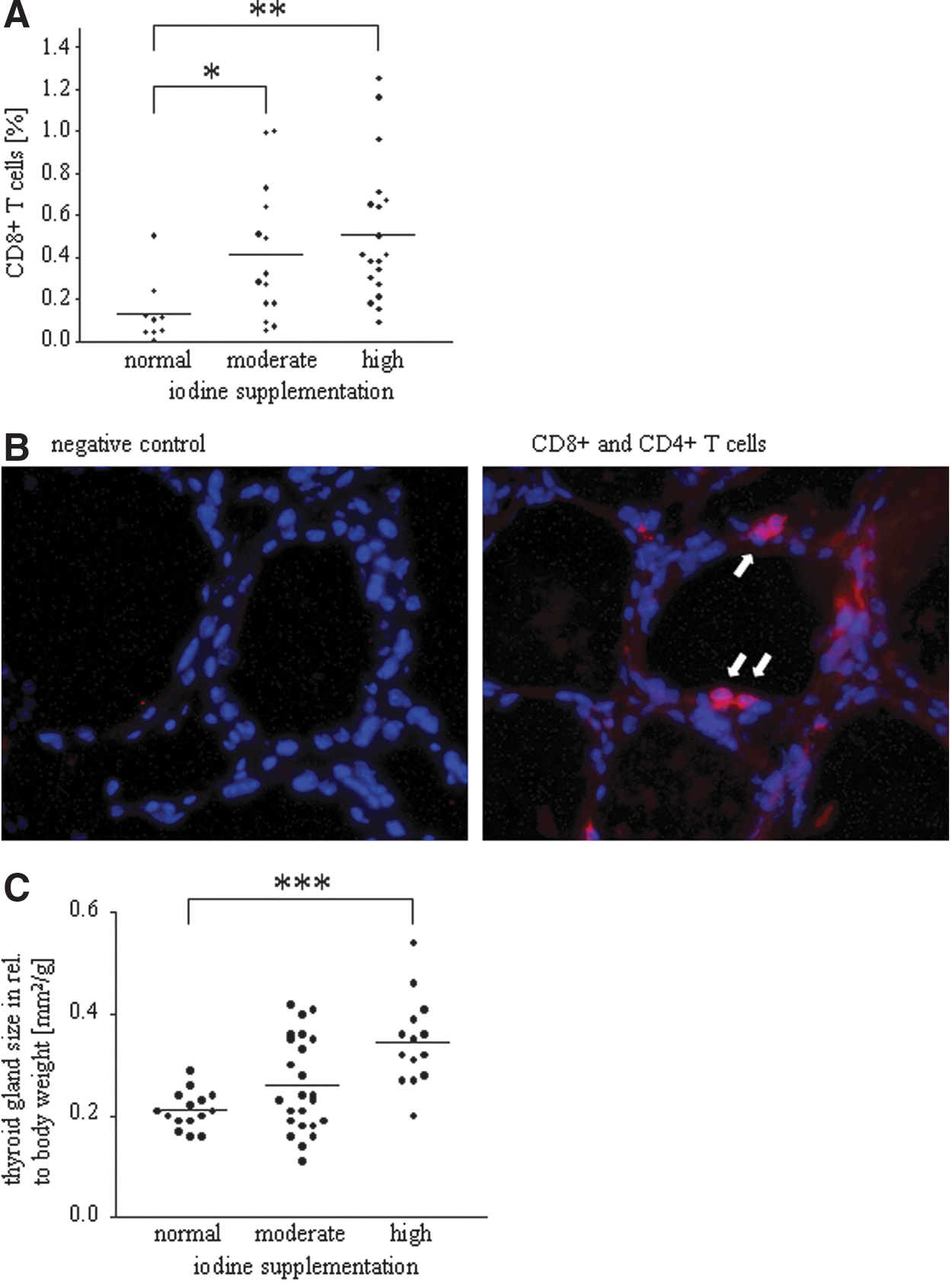
Enhanced iodine supplementation increases lymphocytic thyroid infiltration as well as thyroid size.
Impact of enhanced iodine supplementation on cellular immunity
The impact of enhanced iodine supplementation on cellular immunity was measured by FACS analysis of splenic single cell suspensions. The percentage of CD45+/CD8+ T cells was significantly enhanced in the group of mice treated with elevated amounts of iodine (normal iodine intake, 8.9±6.3%; moderate iodine intake, 18.0±12.9%, p=0.0254; high iodine intake, 11.9±7.7%; Fig. 3A). In contrast, however, no changes were seen in the number of activated cytotoxic T cells with CD107a positivity (Fig. 3B; activated cytotoxic T cells: CD45+/CD8+/CD107a+). A significant elevation of CD45+/CD25/FoxP3+ regulatory T cells was observed in the group of mice treated with a moderate amount of iodine (normal iodine intake, 0.2±0.5%; moderate iodine intake, 1.6±1.7%, p=0.0133; high iodine intake, 0.5±0.4%; Fig. 3C). Additionally, mice treated with elevated iodine levels showed a significantly reduced amount of CD45+/CD335+ natural killer (NK) cells when compared to the control mice (normal iodine intake, 25.2±25.4%; moderate iodine intake, 10.5±8.8%, p=0.0253; high iodine intake, 8.4±7.5%, p=0.0059; Fig. 3D).
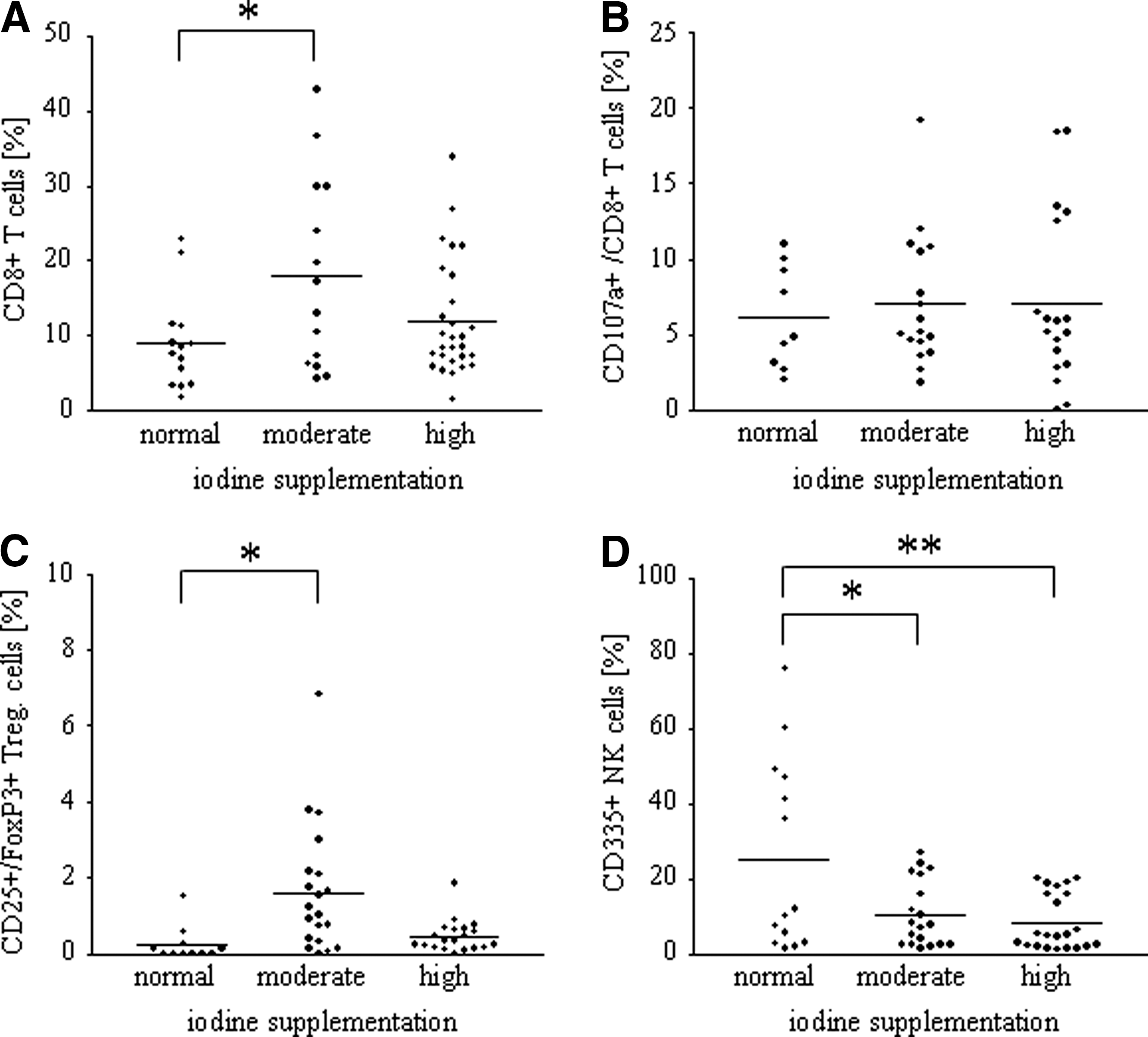
Iodine dependent leukocytic alterations in splenic mononuclear cells. The impact of enhanced iodine supplementation on leucocytic alterations was measured by fluorescein-activated cell sorter (FACS) analysis of splenic single cell suspensions.
Impact of enhanced iodine supplementation on cytokine production
The impact of enhanced iodine supplementation on cytokine production was measured by using FACS analysis of splenic single cell suspensions. The groups of mice supplemented with elevated amounts of iodine exhibited a lower percentage of IFN-γ producing CD45-positive leukocytes and also a reduced intensity of IFN-γ production compared to the control mice with normal iodine intake (normal iodine intake, 87.0±4.7% and 30.2±15.9 mean fluorescent intensity [MFI]; moderate iodine intake, 71.9±19.3%, p=0.0065 and 25.4±14.9 MFI; high iodine intake, 80.9±14.4% and 18.3±9.9 MFI, p=0.0079; Fig. 4A). Regarding IL-1α, additional iodine supplementation led to a higher percentage of IL-1α producing CD45 positive leukocytes, as well as a higher intensity of IL-1α production (median [25th, 75th percentile] for IL-1α positive cells and mean±standard deviation for MFI: normal iodine intake, 1.2% [0.48%, 3.39%] and 25.0±18.3 MFI; moderate iodine intake, 4.2% [1.15%, 27%], p=0.0211 and 56.7±42.5 MFI, p=0.0115; high iodine intake, 3.7% [2.6%, 7.75%], p=0.0085 and 44.8±29.5 MFI, p=0.0343; Fig. 4B). The percentage of IL-17 producing CD45 positive leukocytes and the intensity of IL-17 production remained unaffected by iodine supplementation (mean±standard deviation for IL-17 positive cells and median [25th, 75th percentile] for MFI: normal iodine intake, 30.7±23.6% and 20.6 [9.88, 49.5] MFI; moderate iodine intake, 27.8±25.3% and 28 [1.3, 41.8] MFI; high iodine intake, 29.0±30.3% and 36 [15, 46] MFI; Fig. 4C).
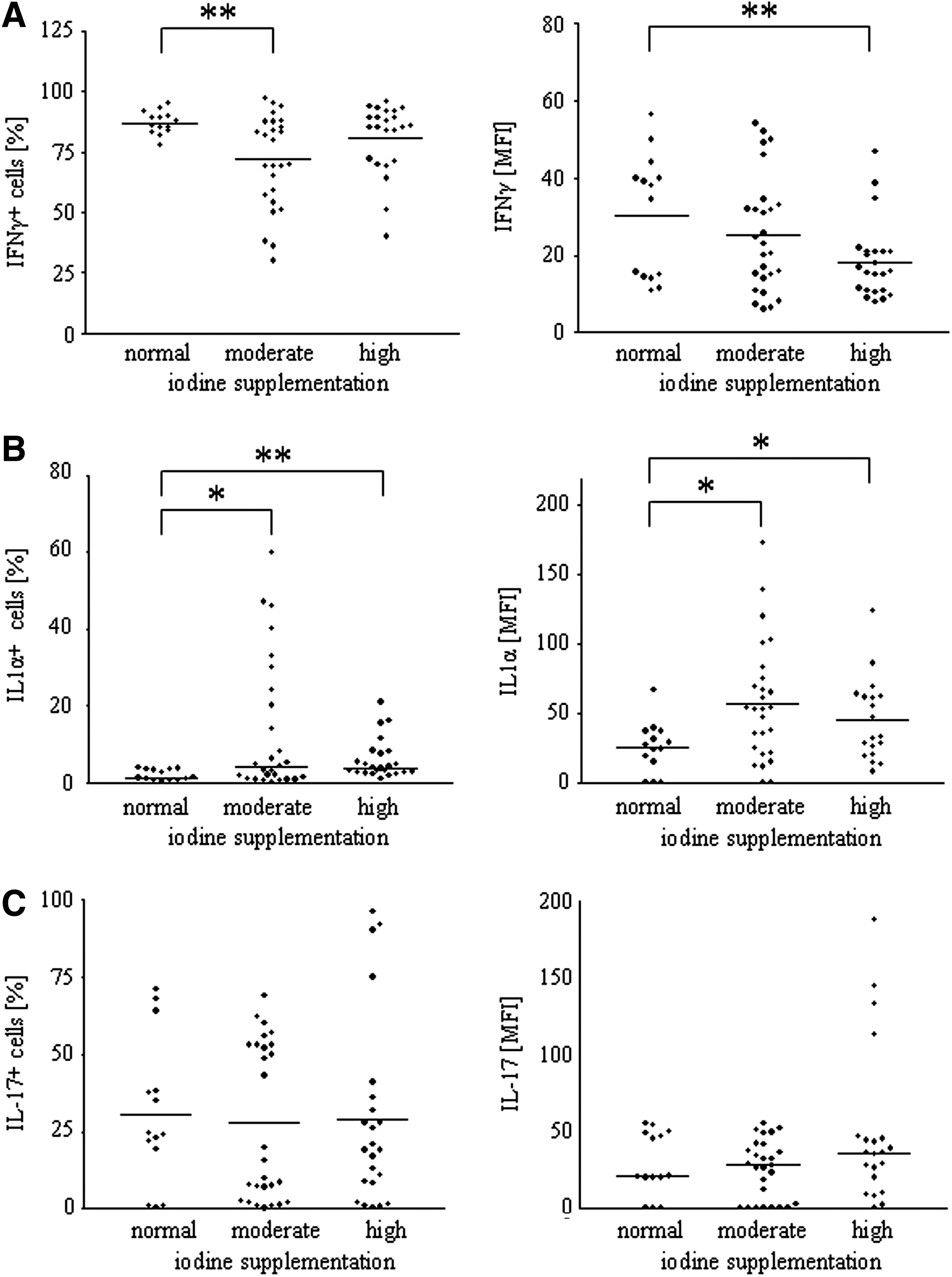
Iodine dependent alterations on cytokine production. The impact of enhanced iodine supplementation on cytokine production was measured by fluorescein-activated cell sorter (FACS) analysis of splenic single cell suspensions.
Discussion
In this study, we demonstrate that moderate and high iodine supplementation alters the immune balance in TAZ10 mice, which are immunologically prone to develop AIT. We saw some changes in the cellularity and cytokine milieu, with the most significant alterations being detected in the innate immune system represented by NK cells. Here, a large decrease of peripheral NK cells was seen. In addition, a moderate increase in lymphocytic infiltration into the thyroid gland was observed in the iodine-supplemented groups. All these alterations did not, however, lead to any significant changes in the clinical outcome in these mice, which had similar TSH serum levels and no significant changes in body weight.
Due to the well-accepted assumption of T helper 1 cells being dominant for the cellular immune response in HT, we used transgenic TAZ10 mice for our study as these mice represent a T helper 1 cell mediated mouse model for thyroiditis. These mice are characterized by massive thyroid lymphocytic infiltration, low or undetectable serum thyroxine, increased serum TSH, and increased body weight (21). In comparison to other well-established animal models in which thyroiditis develops either spontaneously (obese strain chickens, BB rats, non-obese diabetic [NOD].H-2h4 mice) or in which thyroiditis is induced by using homologous TPO or Tg (24 –28), only TAZ10 mice show a pure cellular immune response without participation of the humoral immune system. This is the case because TAZ10 mice are backcrossed to Rag1 −/− and are devoid of functional antibodies (21). To investigate the cellular mechanisms and clinical outcomes after long-term exposure to increased iodine intake without the participation of autoantibodies, TAZ10 mice were used in our study. In accordance with former studies in obese strain chickens, BB rats, NOD, and NOD.H-2h4 mice, in which an increased lymphocytic thyroid infiltration led to AIT following elevated dietary iodine intake (24,29 –32), our present study revealed similar results: moderate and high iodine intake significantly enhanced thyroid infiltration with CD8+ T cells, which presumably leads to the increase of the thyroid gland size and additionally correlates with an increase of peripheral T cells. This, did not, however, alter the clinical course of disease as we did neither detect any further effect on the weight nor on the TSH serum levels, which are elevated in all TAZ10 mouse groups irrespective of iodine supplementation in comparison to the CBA wild-type mice. At this point, however, we cannot exclude that a longer experimental time frame would lead to clinically apparent hypothyroidism (elevated TSH values) as already described by others (33,34).
In contrast to NOD.H-2h4 mice (35), moderate iodine supplementation was associated with an elevated number of CD25/FoxP3 positive regulatory T cells in TAZ10 mice. One could critically argue that the reduced systemic cellular activity may be linked to an enhanced migration into the thyroid and that the CD25/FoxP3 positive cells resemble activated T cells. However, this seems very unlikely as physical hallmarks of AIT were not detectable. No significant results were detected in the group of mice treated with high iodine. One reason for this surprising result might be given by the required duration of iodine supplementation based on the known preclinical phase in AIT: despite the presence of lymphocytic infiltration of the thyroid and despite the development of autoantibodies against TPO and Tg, patients can, at last initially, remain clinically healthy (36). Therefore, thyroid autoimmune changes can have a clinical effect on thyroid function only in the long term. Longer exposure to iodine might have provided significant results. Nevertheless, in analogy to retrospective human studies, mice were supplemented with increased nutritional iodine intake for 19 weeks, resembling one-fourth of their lifespan (11).
Regarding the frequency of NK cells, we saw an iodine-dependent decrease of CD335+ NK cells in splenic mononuclear cells. These data are in contrast to other reports: Chen et al. reported an iodine-dependent induction of NK cell activity in vitro that has been determined by measuring the cytotoxic capacity of murine (BALB/c) spleen cells toward YAC-1 cells (37). Kawashima et al. reported on an increase of IL-18 mRNA expression by thyroid cells itself, following stimulation of BALB/c mice with high amounts of iodine in vitro (38). This is important because IL-18 is a potent inducer of NK cell activation (39,40). These discrepancies might be explained by the higher amount of immunoregulatory NK cells induced by IL-18, as recently reported by us (41) and by others (42), leading to immune suppression in autoimmune and malignant diseases.
Because of the aforementioned iodine-induced thyroid infiltration with CD8+ T cells, we additionally analyzed the frequency of peripheral leukocytes as well as inflammatory cytokines such as IFN-γ, IL-17, and IL-1α, which are linked to iodine-induced AIT in NOD.H-2h4 mice (43 –45). Both IFN-γ and the proinflammatory cytokine IL-17 produced by Th17 cells, were required for development of lymphocytic spontaneous AIT in NOD.H-2h4 mice (43,45). In our study, the number of IFN-γ producing T cells, as well as the intensity of cytokine production was reduced in the iodine-treated mice. This observation is consistent with the previously described decrease of NK cell frequency in human as NK cells are the main producers of endogenous IFN-γ (46). In our study, we neither saw any effect on IL-17 producing CD45 positive leukocytes, nor on the intensity of IL-17 production. Contrary to the observations using NOD.H-2h4 mice, our data suggest a protective or at least a negligible effect of elevated iodine levels on the innate immunity of TAZ10 mice. Parts of the innate arm are downregulated (frequency of NK cells, IFN-γ positive cells as well as the intensity of IFN-γ production), while other parts are upregulated (IL-1α). IL-1α, a proinflammatory cytokine, was present in both groups of mice supplemented with higher amounts of iodine. The absent clinical impact of IL-1α, which has been shown to impair the epithelial thyroid barrier (47), on the course of disease in these mice may be due to the absence of antibodies, the presence of regulatory T cells and presumably antagonistic cytokines.
Finally, one has to ask about the effect of iodine on thyroiditis and the responsible mechanisms. Four possible mechanisms are discussed most commonly: (i) iodine by itself is associated with the development of thyrocyte necrosis, thereby leading to thyrocyte death and additional thyroiditis due to enhanced immunologic infiltration (48); (ii) iodine and IFN-γ seem to cooperate to promote thyroidal expression of intercellular adhesion molecule 1 (ICAM-1), at least in the NOD.H-2h4 mouse model leading to additional infiltration of lymphocytes (49); (iii) iodine stimulates thyroid follicular cells to produce chemokines, particulary CCL2, CXCL8, and CXCL14, which attract immunocompetent cells into the thyroid (50); and (iv) iodine excess leads to iodinated peptides in Tg that are described to enhance the immunogenicity of that antigen [reviewed in Carayanniotis (51)]. One has to keep in mind that the used TAZ10 mouse model was TPO-specific. A stronger immunologic effect might have be seen in a Tg-based model (51 –55).
In conclusion, clinical hallmarks of AIT such as increase of serum TSH and increase of body weight were not augmented in TAZ10 mice supplemented with moderate and higher amounts of iodine, even though thyroid infiltrates were shown to be increased. Although iodine supplementation selectively altered immune cell frequency and cytokine production, the overall immunologic balance remained unaffected. Therefore, we propose that iodine supplementation alters the T helper 1 cell driven AIT in immunologically prone individuals with low clinical effect: despite the increase in thyroid gland infiltration, this is not sufficient to alter TSH levels. However, this hypothesis, including a detailed immunological monitoring also of thyroid infiltrating T cells, has to be investigated in AIT patients as well.
Footnotes
Acknowledgments
We thank Roswitha Charko and Ulla Dötter for excellent technical assistance.
Author Disclosure Statement
No competing financial interests exist.