
Abstract
Background:
3,5-Diiodo-L-thyronine (3,5-T2), a potential metabolite of 3,3′,5-triiodothyronine (T3), exerts marked metabolic actions without the undesirable cardiac and central side effects of T3. So far the lack of reliable quantification methods for endogenous 3,5-T2 in human serum has limited further insight into its physiological and pathophysiological roles in endocrine homeostasis and disease status.
Methods:
Monoclonal anti-3,5-T2 antibodies (3,5-T2 mAbs) were produced in mice. We developed a competitive chemiluminescence immunoassay (CLIA) with one selected mAb and optimized it for high sensitivity, linearity, recovery, and low cross-reactivity to structurally related thyroid hormones (THs) and thyronamines. The CLIA was then used to investigate the origin and action of 3,5-T2 in humans under physiological and pathophysiological conditions in comparison with THs. Patient analysis included individuals with confirmed hypo- or hyperthyroidism and a separate population of thyroidectomized patients on L-thyroxine (T4) replacement therapy.
Results:
3,5-T2 is stable in human serum after storage at 4°C or room temperature as well as several freeze–thaw cycles. The immunoassay did not show any significant cross-reactivity with naturally occurring TH metabolites in physiological and pathophysiological concentrations. The assay shows a lower detection limit of 0.2 nM 3,5-T2 and an upper detection limit of 10.0 nM. The newly established CLIA generates reliable results after spiking exogenous 3,5-T2 or by linear dilution of sera. Intra-assay variation is between 4.1% and 9.0%. Overall mean of variation between different assays is 5.6%–12.9%. 3,5-T2 serum concentrations do not differ in hyperthyroid (0.31±0.02 nM, n=24) compared to hypothyroid (0.43±0.04 nM, n=31) individuals. 3,5-T2 was detectable and elevated in serum from thyroidectomized and T4-substituted patients (0.48±0.03 nM, n=100) in comparison to a sex- and age-matched control group (0.29±0.01 nM, n=99).
Conclusion:
The established CLIA is highly specific, sensitive, precise and accurate for 3,5-T2 detection in human serum. Because 3,5-T2 is not regulated in conditions of an altered thyroid state, it is most likely that serum 3,5-T2 concentrations are not directly dependent on feedback regulation via the hypothalamic–pituitary axis. In addition 3,5-T2 is present in thyroidectomized individuals on T4 substitution, and it is elevated after T4 substitution compared with healthy controls. We conclude that these data support extrathyroidal production of 3,5-T2 from T4.
Introduction
T
Direct molecular targets and cellular mechanisms of action of 3,5-T2 have been studied in vivo mainly in various hypothyroid rodent models and in cellular and mitochondrial in vitro models. Among cellular targets postulated to contribute to the rapid metabolic action are the plasma membrane, the cytosol, and mitochondria.
3,5-T2 enhances loss of body fat by increasing energy expenditure (10,11) prevents insulin resistance (12,13), and exerts remarkably rapid anti-steatotic (14 –16) and metabolic actions (11,17) upon administration of pharmacological doses in mainly hypothyroid rodent models, two human volunteers (3), and hepatic in vitro cellular models (18,19).
Oxygen consumption is stimulated by 3,5-T2 treatment in vitro, using perfused livers (20), liver homogenate (21), or liver mitochondria isolated subsequently after 3,5-T2 pretreatment of hypothyroid rats (22). 3,5-T2 increases rat liver oxidative capacity (cytochrome C oxidase [COX] activity) and mitochondrial respiratory rate without any noticeable effect on mitochondrial mass or protein content (21,23). Addition of 3,5-T2 to liver homogenates from euthyroid rats had no effect on COX activity (24). 3,5-T2 is also not effective under conditions of direct incubation with isolated mitochondria. COX provides a specific binding site for 3,5-T2, thereby abolishing the allosteric inhibition of respiration by ATP (25,26). The underlying mechanism is a partial uncoupling of oxidative phosphorylation (27) and an intrinsic uncoupling of COX in the presence of higher membrane potential (28,29). These experiments explain in part the effects of 3,5-T2 on basal metabolic rate. Recently, increased expression and activity of hepatic mitochondrial F(0)F(1)-ATP synthase were reported after chronic treatment of hypothyroid rats with 3,5-T2 (8,30).
Besides these direct mitochondrial actions, several other rapid effects at the plasma membrane, influencing proton transport (31,32) and Ca2+ signaling (33), have been identified.
Apart from its potential therapeutic applications, the physiological relevance of the endogenous TH metabolite 3,5-T2, detected in serum and tissues, remains to be explored. The biosynthesis of 3,5-T2 in vivo seems to be independent from the thyroid, as the content of 3,5-T2 in thyroglobulin is about 0.07 % that of T4 on a molar basis (34). Faber et al. (35) showed for the first time that about 15% of T3 was converted in an extrathyroidal compartment to 3,5-T2 in euthyroid subject. T3 injection into euthyroid rats results in a fivefold increase in 3,5-T2 in the circulation after 12 h (36), supporting the idea of T3 as a precursor, although this has not yet been demonstrated in vitro. Nevertheless, neither the direct 5′-deiodination of T3 generating 3,5-T2 nor the tissues, metabolic pathways, or enzymes involved in this reaction have yet been characterized in vitro (37). One important reason is the lack of sensitive and reliable analytical (liquid chromatography–tandem mass spectrometry [LC-MS/MS]) and/or diagnostic immunoassay methods for the detection of 3,5-T2.
The first antibody-based quantitative detection method for 3,5-T2 described in the literature was published in 1978 by Meinhold and Schurnbrand (38). They produced polyclonal anti-3,5-T2 antisera in rabbits and established a radioimmunoassay (RIA) for the measurement of 3,5-T2 in human serum and thyroid tissue after fractional enrichment of 3,5-T2 from 2 mL of serum. Several comparable RIAs for 3,5-T2 detection followed in a short period of time (34,35,39 –43). They all used 125I-labeled 3,5-T2 as tracer, except for Meinhold and Schurnbrand (38) ((3H)acetyl-3,5-T2) and Pinna et al. (43) (3-Br-5[125I]T1). The overall range of serum volume used for these assays was 100–500 μL. Pre-analytical procedures were avoided and serum was used for RIA analysis without extraction procedures. The detected 3,5-T2 serum concentrations in euthyroid subjects varied among the different RIAs between 0.01 nM (42) and 0.19 nM 3,5-T2 (41). This 16-fold difference in 3,5-T2 serum levels makes it hard to compare the different methods and to establish a reference range. Unfortunately, none of the anti-3,5-T2 antisera produced at that time are available or still in use.
Here, we present for the first time a monoclonal antibody (mAb) directly targeting 3,5-T2. Based on this mAb we set up a competitive chemiluminescence immunoassay (CLIA) for the measurement of 3,5-T2 in human sera. This immunoassay provides the analytical tool to monitor 3,5-T2 as a potent endogenous as well as pharmacological TH parameter influencing energy metabolism.
Materials and Methods
Reagents and equipment
Low cross buffer was procured from Candor Bioscience GmbH (Wangen, Germany). EZ-Link Plus activated Peroxidase Kit, ImmunoPure MAb isotyping kit, SuperSignal ELISA femto substrate, and Slide-A-Lyzer dialysis cassettes 2K MWCO were purchased from Pierce Chemical Co. (Rockford, IL). Fast performance liquid chromatography (FPLC) equipment and Sepharose-rProtein A were obtained from GE Healthcare (Freiburg, Germany). Fetal calf serum, Dulbecco's modified Eagle's medium, horse serum, and protein-free II medium were purchased from Invitrogen GmbH (Karlsruhe, Germany). Mini-PERM devices were obtained from Heraeus (Hanau, Germany). Europium-labeled streptavidin, enhancement solution, and VICTOR3 multilabel counter were procured from PerkinElmer (Waltham, MA). Reagents not further specified were purchased from Merck (Darmstadt, Germany) or Sigma (St. Louis, MO). 3,5-T2 for the immunization of mice was synthesized by Dr. R. Smits and Dr. B. Koksch (Freie Universität Berlin, Germany) and purified to ≥99% by Dr. R. Thoma (Formula GmbH Pharmaceutical and Chemical Development Company/Henning Berlin Arzneimittel GmbH, Berlin, Germany). All TH-related substances used for cross-reactivity studies were obtained by Dr. R. Thoma or kindly supplied by Dr. T. Scanlan (Oregon Health and Science University, Portland, OR). SeraCon II and SeraCon II CD human serum were kindly provided by SeraCare (Milford, MA). SeraCon II CD is stripped of T3, T4, and diiodothyronines (44). Ninety-six-well flat-bottom Maxisorp microtiter plates were purchased from Nunc (Roskilde, Denmark).
Biotinylation of antibodies
Goat anti-mouse IgG Fc-specific antibody was biotinylated as described elsewhere (45) using a 75-fold molar excess of the labeling reagent biotin-amidocaproate-N-hydroxysuccinimideester.
Conjugation of 3,5-T2 to bovine serum albumin and horseradish peroxidase
Bovine serum albumin (BSA) and 1-ethyl-3-(3-dimethylaminopropyl)carbodiimide (EDC) were dissolved in demineralized H2O. 3,5-T2 was dissolved in dimethylsulfoxide and diluted in 0.1 N NaOH. Solubilized 3,5-T2 was mixed with BSA solution. Coupling reagent EDC was added drop wise. The pH was adjusted to 9.0. The mixture was stirred at 4°C for 24 h to form 3,5-T2-BSA. 3,5-T2 was conjugated to aldehyde-preactivated horseradish peroxidase (HRP) via reductive amination cross-linking using EZ-Link Plus Activated Peroxidase Kit. Because 3,5-T2 provides only one amino group for cross-linking 3,5-T2 was used in 100-fold excess. Conjugation was performed at pH 9.4, following the supplier instructions. Incubation time was prolonged to 20 h at 4°C. 3,5-T2-BSA and 3,5-T2-HRP solutions were subsequently dialyzed against 50 mM phosphate buffer, using Slide-A-Lyzer Dialysis Cassettes. Finally, 3,5-T2-BSA and 3,5-T2-HRP solution were sterile-filtered. 3,5-T2-BSA was stored in aliquots at −20°C. 3,5-T2-HRP was stored at 4°C.
Production of mAbs specific for 3,5-T2
Three 2-month-old female BALB/c mice were used for immunization. Prior to immunization, control serum was collected by tail vein puncture. 3,5-T2-BSA was dissolved in TiterMax Gold according to the supplier's recommendations to prepare immunization solution. At day 1, all mice were injected subcutaneously at three to four abdominal sites with 50 μg of antigen solution per mouse. Immunization was repeated with half the dose per animal after 4 and 9 weeks. Anti-3,5-T2-BSA antibody titer in serum of immunized mice was tested 4 weeks after the second injection and 2 weeks following the third injection. Finally, mice were injected twice intraperitoneally with 25 μg of 3,5-T2-BSA in phosphate-buffered saline (PBS) per mouse before spleen cell harvest. Immunized mouse sera were tested both for polyclonal anti-3,5-T2 and anti-BSA antibodies. Isolated lymphocytes were fused with murine myeloma cells (PAI) to create immortalized hybridoma cell clones (46). Hybridoma cells were selected for high reactivity to 3,5-T2, no reactivity to BSA, best competition with unlabeled 3,5-T2, and low cross-reactivity to structurally related THs, decarboxylated TH (thyronamines [TAMs]), and iodothyroacetic acids (TAcs). Selected anti-3,5-T2 mAb–producing hybridoma cells were subcloned by limiting dilution and expanded.
Hybridoma supernatant and mouse serum screening procedure
A 96-well high-binding microplate was coated with either 10 ng 3,5-T2-BSA/0.1 mL 50 mM Na2HPO4 buffer (pH 7.4) or 10 ng BSA/0.1 mL Na2HPO4 buffer and incubated at 4°C overnight. The plate was washed three times with PBST (0.05% Tween 20 in PBS). Mouse serum was diluted 1:1000 in assay buffer [50 mM Tris-(hydroxymethyl)-aminomethane, 154 mM NaCl, 20 μM diethylenetriaminepenta-acetic acid, 0.01% Tween 40, and 0.05% NaN3 (pH 7.75)]. Immobilized 3,5-T2-BSA or BSA was incubated with diluted serum from immunized mice or hybridoma supernatant (0.1 mL/well) for 1.5 h at room temperature under agitation. The plate was washed again three times with PBST. In the next step biotinylated goat anti-mouse IgG Fc-specific antibody (10 ng/0.1 mL) was added for 1.5 h at room temperature under agitation. The plate was washed three times with PBST. Streptavidin-europium (10 ng/0.1 mL) was added for 0.5 h under agitation. The plate was washed six times with PBST. In the last step 0.2 mL of enhancement solution was added, and the plates were incubated for 10 min under agitation. The fluorometric signal was detected using a VICTOR3 multilabel counter.
Immunoglobulin isotyping and mAb purification
The mAb isotype was tested with a commercially available Immunopure mAb isotyping kit, following the manufacturer's protocol. Monoclonal antibodies were purified by FPLC on an rProtein A-Sepharose column, with respect to antibody isotype.
Ultrasensitive CLIA procedure
White microtiter plates were coated overnight at 4°C with 75 ng of goat anti-mouse IgG Fc-specific diluted in Na2HPO4 buffer (pH 7.4). The plates were washed with PBST (0.05% Tween 20) and incubated for 1 h at room temperature with 1% BSA-PBST (0.02% Tween 20). The plate was washed again with PBST. 3,5-T2 standards (0–10 nM) diluted in SeraCon II CD serum, and serum samples (25 μL of each) were loaded to the plate in duplicate. Next, 4 ng of mAb 4F8 diluted in 1:1 0.5% BSA-PBST/low cross buffer was added per well. The plates were incubated for 5 min under agitation and placed at 4°C for overnight incubation. After pre-incubation, 0.2 ng of 3,5-T2-HRP was added as tracer. The plates were incubated at room temperature under agitation for 5 min, following 1 h of incubation at 4°C. After a final wash step, 50 μL chemiluminescence substrate was added per well, and the plates were incubated for 1 min under agitation. The chemiluminescence signal was detected by multilabel counter. Commercially available sera were used as internal quality control (SeraCon II [SeraCare], Lyphochek [Bio-Rad, München, Germany], and off-the-clot serum [PAN-Biotech, Audenbach, Germany]).
3,5-T2 assay validation
The limit of detection calculation is based on 3,5-T2 measurement in analyte-free assay matrix. Three times the standard deviation (SD) was added to the mean (n=24 single measurements) value of the assay blank. Intra-assay precision was assessed by assaying four individual human sera (n=12 single measurements on one plate each) with 3,5-T2 concentrations from 0.5 to 1.5 nM. We evaluated interassay precision from a series of 15 assays with three different human sera covering a broad 3,5-T2 range. For determination of assay accuracy, five different sera with low, medium, and high amounts of the analyte were spiked with additional amounts of 3,5-T2. In addition, we performed serial dilution of five sera with known high concentrations of 3,5-T2 in the serum matrix. Dilution linearity was calculated by multiplying the assayed 3,5-T2 concentration with the dilution factor, following a comparison with the undiluted concentration. Cross-reactivity of the 3,5-T2 chemiluminescence assay was determined by assaying known concentrations of structurally related THs and TAMs (10–250 nM) diluted in TH-deficient serum matrix and quantified in terms of 3,5-T2–equivalent assay response.
Clinical subjects
Clinical samples representing thyroidal illnesses were selected from patients with an initial diagnosis of either hypothyroidism (8 men and 23 women, 20–84 years; body mass index [BMI], 17.5–45.2; free [f]T4, 5.7±0.6 pM; fT3, 2.9±0.2 pM; thyrotropin [TSH], 65.1±7.8 mU/L) or hyperthyroidism (5 men and 19 women, 21–75 years; BMI, 16.8–35.6; fT4, 43.3±4.5 pM; fT3, 19.1±1.9 pM; TSH, 0.01±0.0 mU/L). The hypothyroid group included 23 patients with Hashimoto's thyroiditis and nine patients with postablative 131I-induced hypothyroidism. The hyperthyroid group consisted of 18 patients with Graves' disease and six patients with toxic nodular goiter. Another clinical cohort of patients consisted of 100 thyroidectomized thyroid cancer patients on TSH-suppressive T4 therapy (78 women and 22 men, age 18–84 years). This group of patients consisted of 73 individuals with papillary carcinoma, 21 with follicular carcinoma, two with Hürthle cell carcinoma, and four with medullary thyroid carcinoma. As a healthy control group, 99 age-matched (±2 years) and sex-matched individuals from the Study of Health in Pomerania (SHIP-TREND) were chosen (47). All patients provided their informed consent and all studies were approved by the local ethics committee.
Statistical analysis
The standard curve to calculate 3,5-T2 serum concentrations was fitted to a sigmoidal dose–response curve with variable slope (four-parameter logistic equation) using GraphPad Prism version 5.01 (GraphPad Software, Inc., San Diego, CA). The 3,5-T2 serum concentrations of the cohort with hyper- and hypothyroid patients were adjusted for age and BMI followed by a univariate variance analysis using SPSS Statistics version 19.0.0.1 (IBM, Armonk, NY). For all other statistical tests, GraphPad Prism version 5.01 was used. Relative recovery values are expressed as mean±SD. Statistical tests are indicated in the corresponding figures. Differences were considered significant if p<0.05.
Results
Production of mAbs specific for 3,5-T2
To generate mAb-targeting 3,5-T2, we used 3,5-T2 cross-linked to BSA for Balb/c mice immunization (Fig. 1A). Four weeks following the second injection with the antigen, polyclonal antibody titers were tested in mouse serum (Fig. 1B, left). The screening assay indicated that two of three mice responded with high antibody production at this time point. The polyclonal antibody titers observed were predominantly due to high reactivity to the BSA carrier protein. The immune response in mouse 1 to 3,5-T2-BSA was slightly elevated compared to the conjugation partner BSA, indicating 3,5-T2 specific antibody production. All three mice were immunized with the antigen for a third time (Fig. 1B, right). Thereafter, only mouse 1 responded properly by doubling the reactivity to 3,5-T2-BSA, including a decent amount of antibodies specific for 3,5-T2. Hybridomas were produced by fusing immortalized murine myeloma cells PAI with lymphocytes from mouse 1, according to the hybridoma technology (46). Almost 50 wells with hybridomas produced antibodies that were able to bind whole antigen 3,5-T2-BSA. However, 41 of them did not react specifically to 3,5-T2 itself, but recognized BSA. Only supernatants from seven wells of hybridomas showed no reactivity to BSA and best competition with 3,5-T2 in the screening assay. The best performing mAbs, 2D2, 4F8, and 9D3, were chosen for cloning, expansion, and further characterization. MAb 4F8 was chosen for establishment of a CLIA due to its high specificity to the analyte and low cross-reactivity with THs and TAMs with similar structure.

Reaction scheme of conjugating 3,5-T2 to BSA to form the antigen for immunization
CLIA optimization
During the overnight assay procedure a limited amount of mAb 4F8 was preincubated with 3,5-T2 containing serum samples to ensure complete analyte binding to the mAb (Fig. 2). HRP-labeled 3,5-T2 was added and incubated for 1 h. The 3,5-T2 level in human serum was determined according to a standard curve by measuring chemiluminescence. All assay components and incubation conditions were optimized to increase assay sensitivity and specificity. PBST mixture with low cross buffer (PBST-LCB) was used to reduce matrix effects. A commercially available 3,5-T2–deficient serum as matrix, also devoid of T4 and T3 (44), provided best results for creating a standard curve. We tested whether this matrix was comparable to 7% BSA as an artificial matrix for dilution of 3,5-T2 standards (Fig. 3A). The deficient serum showed a minor shift to the left compared to 7% BSA diluted in assay buffer. Using either BSA or serum did not influence generation of a proper standard curve for quantification of 3,5-T2. Overall, the linearity and recovery of the CLIA was improved by using the real serum matrix instead of the artificial one (data not shown). Therefore we chose the commercial serum pool as matrix for our standard curve because of its performance during serum dilution and spiking compared to the artificial BSA matrix.
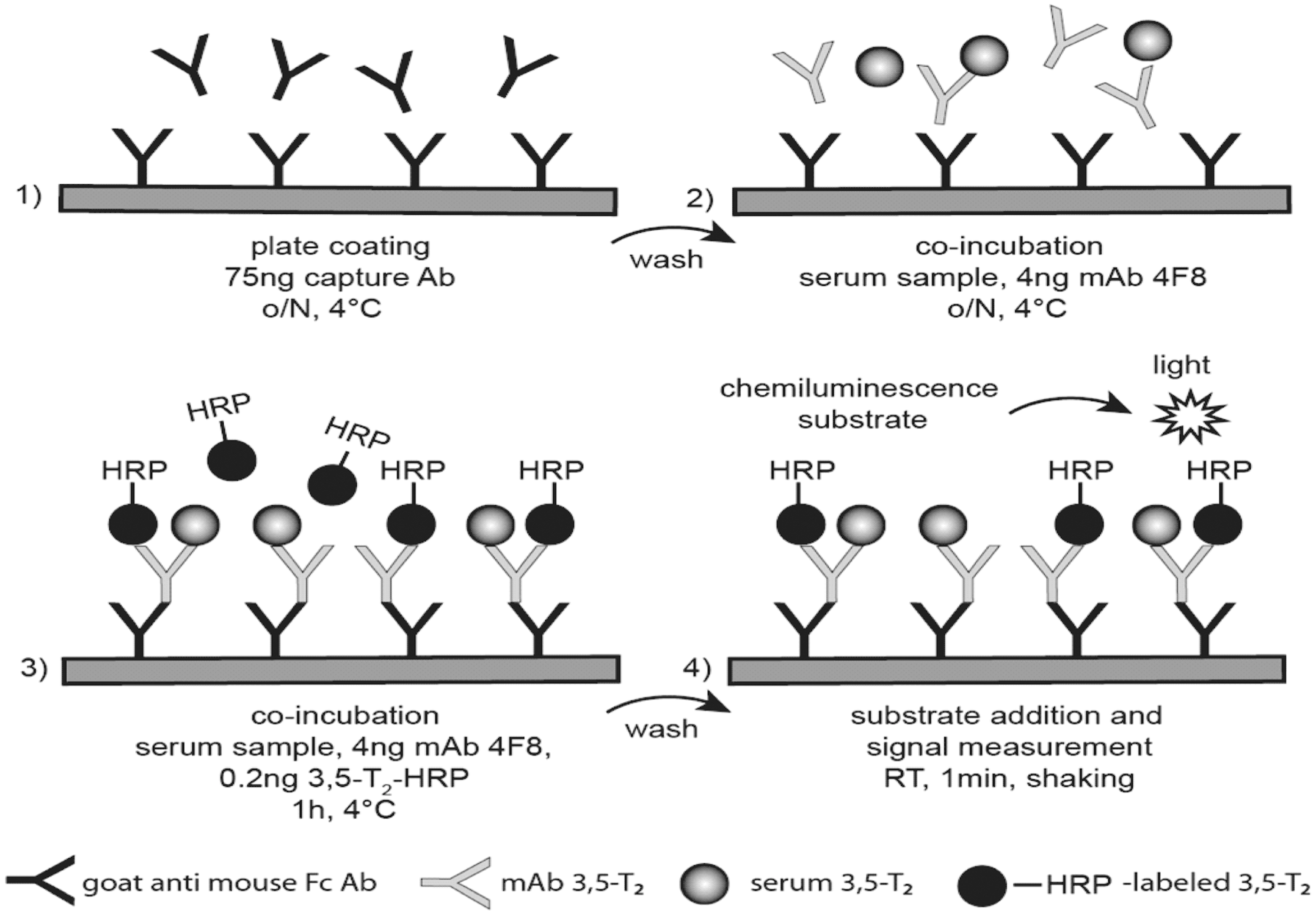
Principle of the developed 3,5-T2–specific chemiluminescence immunoassay (CLIA).
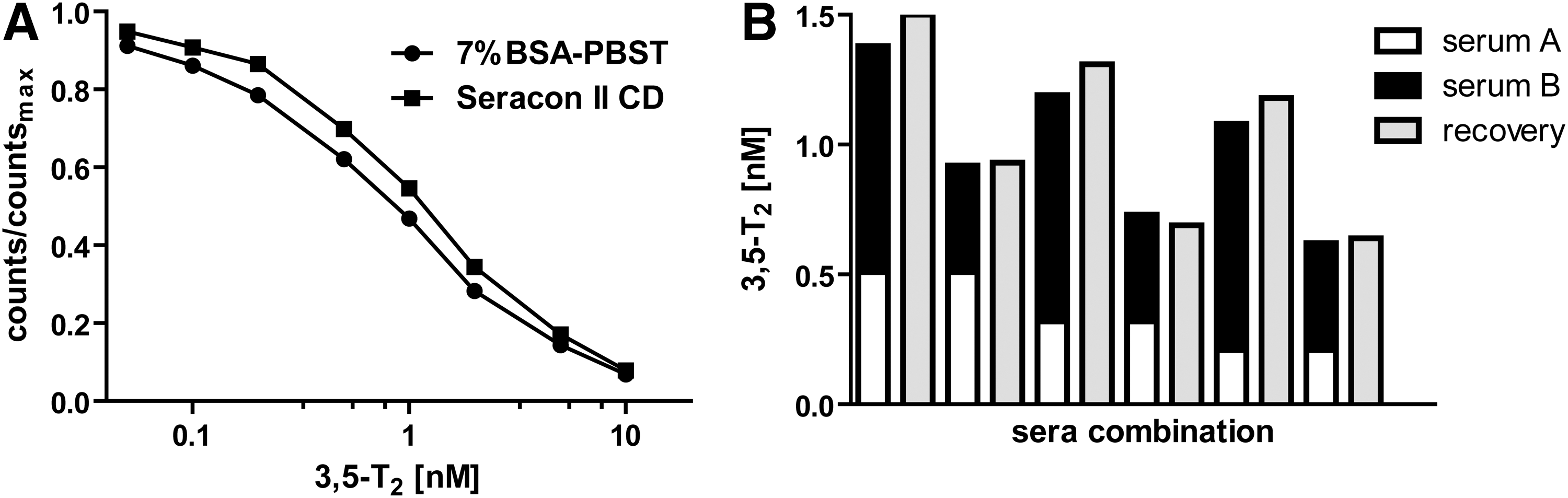
Serial dilution of non-labeled 3,5-T2 in thyroid hormone (TH)-deficient serum (SeraCon CD II) or 7% BSA-PBST
Specificity of CLIA
The mAb 4F8 (IgG1)-based CLIA was characterized for possible assay interference with other THs and TH metabolites (Table 1). Cross-reactivity in the assay was lower than 0.2% for most of the THs (T0, 3-T1, 3′,5′-T2, 3,3′-T2, reverse-T3 [rT3], T4), TAMs (T4AM, T3AM, rT3AM, 3-T1AM, T0AM), and TAcs (T3Ac/Triac, 3-T1Ac) in this assay protocol. Only T3 showed a cross-reactivity of 2.3% at a concentration 50-fold higher than its physiological concentration. Cross-reactivity of 3′-T1 and T4Ac was below 0.5%. We therefore assumed that the detected relative cross-reactivity would not interfere with the detection of 3,5-T2 in serum samples. As expected, mAb 4F8 had some cross-reaction with 3,5-T2AM (3.5 %) and 3,5-T2Ac (10.9%). However, neither of these substances have been detected with any analytical method in serum or in tissue so far (see Table 1, for review see Hoefig et al. [48]). All these data support high specificity of the generated mAb 4F8 to 3,5-T2.
rT3, reverse-T3; NA, not analyzed/available; NQ, not quantified; ND, not detected.
Accuracy and precision of the immunoassay
Immunoassay linearity was assessed by determining serial dilutions of human sera with T2-deficient serum. Relative recovery was calculated by direct comparison to nondiluted sera. Upon 1:2 dilution, the recovery was 92.3±16.0% (n=6 sera). Further 1:4 dilution produced similar results (95.4±11.6%; n=6 sera), indicating good comparability between observed and expected 3,5-T2 concentrations. CLIA accuracy was characterized by spiking human sera with rising concentrations of exogenous 3,5-T2, covering the linear range of the standard curve. Results showed a very good agreement of observed and expected 3,5-T2 concentrations after addition of 0.2 nM (93.7±7.6%) and 0.5 nM (96.8±11.0%). Relative recovery after addition of 1 nM (78.0±9.6%) or 2 nM (85.8±8.3%) was still good. Overall mean recovery of 3,5-T2 in the range of 0.2–2 nM using this mAb-based assay is 88.5±8.5%. Data are derived from five individual human sera in the low, medium, and high range of endogenous 3,5-T2 to ensure coverage of a broad range of individual variations. To further validate CLIA accuracy, we compared equal-volume mixtures of two different sera with at least three additional sera (Fig. 3B). Theoretical values of single sera were compared with the combinations of the equal-volume mixtures. Results showed a good agreement between theoretical and observed 3,5-T2 concentrations, resulting in a relative recovery range of 104.7±6.1%.
Intra-assay precision of the assay was analyzed by repetitive measurement (n=10–12) of 3,5-T2 in individual human sera (n=4), covering a range of 0.5–1.5 nM. Mean measured 3,5-T2 concentrations were 0.6, 0.8, 1.1, and 1.4 nM (see Table 2). The coefficient of variation was 9.0%, 8.9%, 5.0%, and 4.1%, respectively. Interassay precision was calculated based on repeated measurement of three sera on 15 assay plates on different days. Mean measured 3,5-T2 concentrations were 0.5, 1.0, and 1.5 nM (see Table 2). The coefficient of variation was 12.9%, 6.5%, and 5.6%, respectively. The lower detection limit of 0.2 nM in the assay was determined by the mean (0.07 nM) plus three SD (0.04 nM) according to quantification of deficient serum (n=24). The working range of the assay is 0.2 to 10 nM.
Analyte stability
Analyte stability was tested in human serum. To evaluate analyte stability, sera were incubated for 24 h at room temperature or 4°C in comparison to storage at −20°C. Relative recovery was calculated in comparison to −20°C as the control condition. Storage at room temperature (104.4±13.2%) or at 4°C (97.9±9.9%) did not affect analyte stability (n=3 individual human sera, covering low, medium, and high concentrations of endogenous 3,5-T2). Variation in relative recovery was comparable to the calculated intra-assay variation. In addition, different human sera were frozen (−20°C) and thawed four times. Relative recovery was again calculated in comparison to continuous storage at −20°C. Several freeze–thaw cycles did not disturb 3,5-T2 stability (103.0±13.6%; n=8 sera from healthy human individuals). We conclude that 3,5-T2 in serum is stable under different storage temperatures and not affected by four freeze–thaw cycles.
Analysis of human samples representing thyroidal illnesses
3,5-T2 was determined in 31 hypothyroid and 24 hyperthyroid subjects. Detected 3,5-T2 serum concentrations were not significantly different between the groups (0.43±0.04 nM and 0.31±0.02 nM), considering adjustment for age and BMI, respectively (Fig. 4A, see age and BMI distribution in Fig. 4D, 4E). As expected, fT4 and fT3 levels were significantly reduced in hypothyroid subjects and increased in hyperthyroid individuals (Fig. 4B, 4C). Detected 3,5-T2 concentrations in serum showed no correlation to fT3 and fT4 in hypothyroidism (fT3: p=0.07, r=−0.34; fT4: p=0.16, r=−0.26) and hyperthyroidism (fT3: p=0.12, r=−0.35; fT4: p=0.38, r=−0.21) (data not shown).

3,5-T2 serum concentrations (nM) in patients with an initial diagnosis of hypo- or hyperthyroidism. There was no significant difference in the median values between the hypo- and the hyperthyroid group of patients. 3,5-T2 serum concentrations (nM) were adjusted for age and body mass index (BMI). Statistical analysis was done with SPSS using a univariate variance analysis
In addition, sera from thyroidectomized thyroid cancer patients (n=100, 78 women, 22 men) under TSH suppressive T4 therapy were analyzed in comparison to an age- and sex-matched euthyroid group (Fig. 5). Cancer patients (0.48±0.03 nM) showed significantly elevated 3,5-T2 serum concentrations compared to the control group (0.29±0.01 nM) (Fig. 5A, p<0.001). Within the thyroid cancer patient group, no sex-based difference in 3,5-T2 serum values (Fig. 5B) and no obvious correlation between 3,5-T2 and fT4 (Spearman r=−0.08, p=0.89; Fig. 5C) or 3-T1AM were observed (Spearman r=−0.01, p=0.63; Fig. 5D).
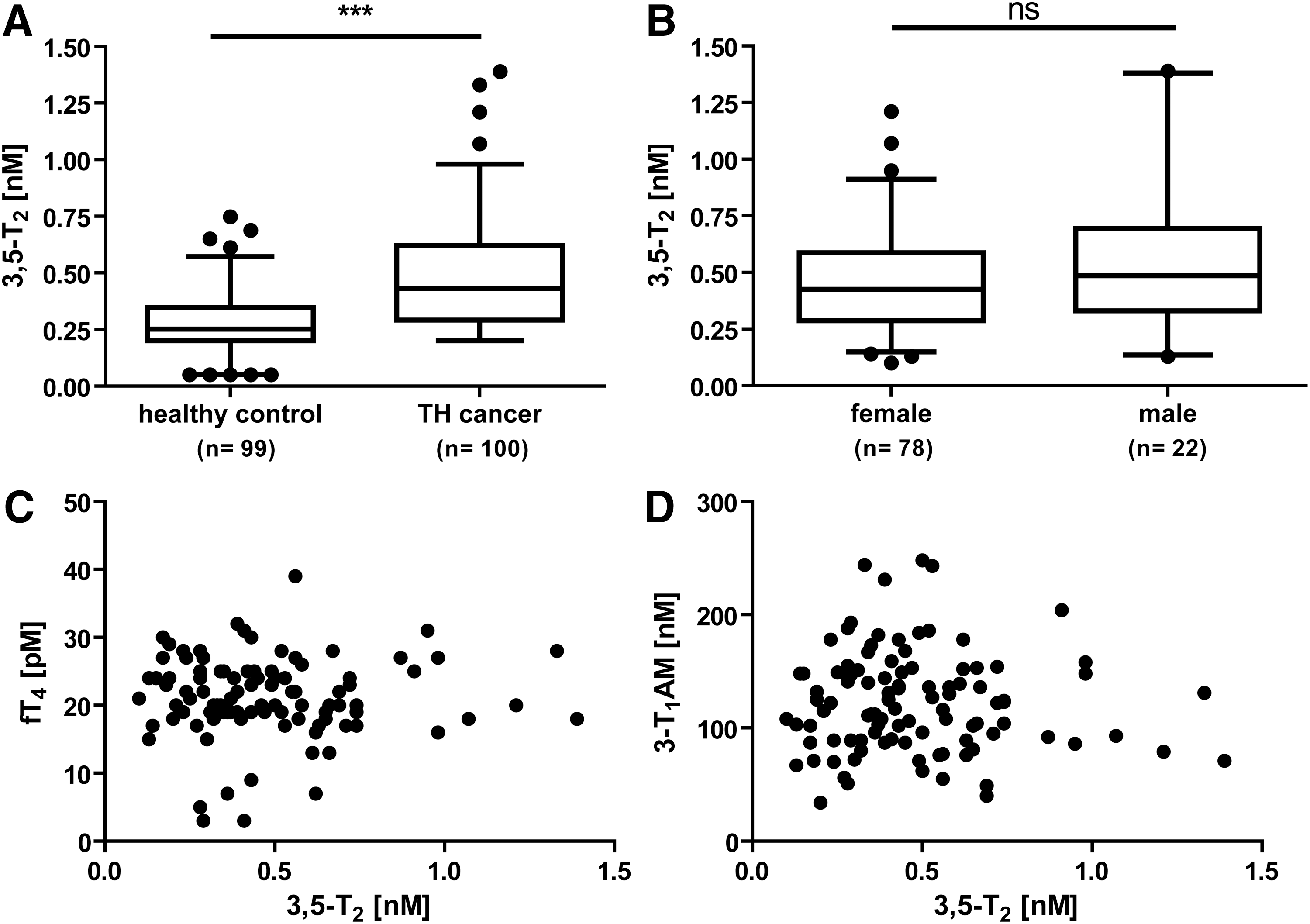
3,5-T2 serum concentration (nM) in thyroid cancer patients (n=100) compared to age-matched (±2 years) and sex-matched healthy control individuals (n=99). 3,5-T2 was significantly increased in thyroid cancer patients in comparison to control group
Discussion
In this study we present a mAb-based CLIA for the quantification of 3,5-T2 in human serum. We generated mAbs highly specific for 3,5-T2. The 4F8 mAb-based immunoassay showed only relevant cross-reactivity to 3,5-T2AM and 3,5-T2Ac. However, both compounds were tested as synthetic substances and have not yet been detected in human or animal sera. In addition, the immunoassay showed a low cross-reactivity with T3. Since we tested a 50-fold (100 nM) higher T3 concentration than the physiological human serum levels (2.1 nM) it is unlikely that, even in hyperthyroid patients, T3 cross-reactivity will influence the assay performance to quantify 3,5-T2. Indeed, serum 3,5-T2 levels were not elevated in hyperthyroid compared to normal or hypothyroid patients. This new assay precisely measures 3,5-T2 in the range of 0.2–10 nM with an intra-assay variability of 4.1%–9.0% and an interassay precision of 5.6%–12.9%. Using the CLIA presented here, we found no differences in 3,5-T2 serum concentrations of patients with thyroid dysfunction. Based on fT3 and fT4, patients were initially diagnosed for hypo- and hyperthyroidism. We found no correlation between free TH levels and measured 3,5-T2 in both groups (data not shown). 3,5-T2 values represent a puzzling parameter in evaluating thyroidal illnesses. In 1979 Maciel et al. (34) did not observe any differences between euthyroid, hypothyroid, and hyperthyroid individuals using a 3,5-T2 RIA. Contrasting data were published by several other groups, stating that 3,5-T2 serum levels are significantly increased in hyperthyroidism and are decreased or not different from euthyroid patients in hypothyroidism (39,42,43). Comparing the two groups analyzed in this study, the number of female (23 hypothyroid, 19 hyperthyroid) and male patients (8 hypothyroid, 5 hyperthyroid) is nearly balanced between the different thyroid states.
We find 3,5-T2 serum concentrations to be significantly elevated in patients with thyroid cancer who are receiving T4 supplementation therapy. Levels were compared to an age-matched (±2 years) and sex-matched euthyroid control group. T4 supplementation in these patients lacking functional thyroid tissue suggests that 3,5-T2 serum concentration can only be explained by extrathyroidal generation via deiodinases. Maciel et al. (34) described extraction of thyroid gland tissue followed by a 3,5-T2 RIA procedure, which resulted in low 3,5-T2 levels compared to T4 levels. Only 0.07% of thyroglobulin-bound THs was 3,5-T2. They concluded that only a low amount of 3,5-T2 is directly produced by the thyroid gland and suggested that most 3,5-T2 is generated via extrathyroidal metabolism. In addition Faber et al. (35) showed that about 15% of infused labeled T3 is extrathyroidally converted to 3,5-T2. Recently, a mAb-based luminescence immunoassay to detect 3-T1AM in human serum was published, reporting an increase of 3-T1AM in T4-substituted thyroid cancer patients and also suggesting extrathyroidal formation of 3-T1AM from supplemented T4 (48). Sera of the same TH cancer patients were now measured and compared to another control group. Since 3,5-T2 was proposed to be the precursor of 3-T1AM (either via 3,5-T2AM or 3-T1), we tried to find a direct link between 3-T1AM and 3,5-T2 in the sera of the same patients with thyroid cancer; however, neither parameter showed correlation.
Polyclonal Ab-based RIAs helped decades ago in forming the initial conception about the functional role of 3,5-T2 in human serum. Nevertheless, the data produced gave controversial results concerning the role of 3,5-T2 in thyroidal and nonthyroidal illnesses and serum levels of this TH metabolite. Unfortunately, none of these polyclonal antibodies remain available for direct comparison. In euthyroid subjects, serum 3,5-T2 levels vary in the literature from 0.008±0.003 nM to 0.14±0.07 nM, which is about 15 times higher (34,39,40,42,43,49). Using the mAb-based CLIA presented here, we measured a mean serum concentration of 0.29±0.014 nM 3,5-T2 including both female and male subjects (mean±SEM, n=99). This first set of data on individuals without thyroid disorders does not yet represent a reference range for 3,5-T2 serum concentrations. Although direct comparison might be elusive, the measured 3,5-T2 concentrations are of the same magnitude as the highest reference values from previous RIA procedures.
We do not know to what extent 3,5-T2 is bound to serum proteins and how this binding might influence the CLIA procedure. As RIA and CLIA give comparable results of the same order of magnitude, we suggest at this point that both methods determine total 3,5-T2 in human serum. As no pre-analytic extraction step is included in the mAb-based CLIA, we expect that the high-affinity 3,5-T2 mAb effectively competes with serum proteins binding 3,5-T2 and that both protein-bound and free 3,5-T2 are quantified in the assay. This hypothesis is supported by several trials to detect 3,5-T2 in human serum with LC-MS/MS–based or electronspray ionization (ESI)-MS/MS–based analytical methods. Zhang et al. (44) published an ESI-MS/MS-based method, using solid phase extraction (SPE) cartridges as offline pre-analytical preparation. Their established protocol worked for TH-free and processed serum spiked with exogenous 3,5-T2. However, no data were included in this report to demonstrate that their method was sensitive enough to detect endogenous 3,5-T2 concentrations in serum. Piehl et al. (37) published an LC-MS/MS–based method to separate 3,5-T2 from 3,3′-T2 and 3′,5′-T2 in a cell culture–based trial. Only one publication based on LC-MS/MS technique, using another SPE cartridge system as the pre-analytical step, at least stated that endogenous levels of 3,5-T2 in human serum are below their validated detection limit (53). Both MS/MS-based analytical methods may lose the protein-bound portion of 3,5-T2 on the SPE column during the pre-analytical step. In addition, both methods need 0.5 mL of serum for a single measurement. Both of these problems, serum binding and LC-MS/MS detection limits, are avoided by using high-affinity mAb CLIA because no pre-analytical step or enrichment protocol is needed for proper measurement. The CLIA works with 50 μL of serum for 3,5-T2 determination in duplicate, one tenth the volume needed in the MS/MS-based technologies. Partial binding of 3,5-T2 to proteins would also explain the varied 3,5-T2 concentrations reported by different assays and the increased analyte stability upon storage at 4°C or room temperature for an overnight period, as well as up to four freeze–thaw cycles. Serum binding of 3,5-T2 might protect the TH metabolite from further modification, including deiodination.
In this work, we generated mAbs and established an immunoassay specifically detecting 3,5-T2 for quantitative studies clarifying the role of 3,5-T2 in serum and tissues of human and animal origin. In addition to the human serum sample cohort presented here, more 3,5-T2 quantifications have to be performed to identify the pathophysiological conditions for which 3,5-T2 might be a useful clinical biomarker.
Footnotes
Acknowledgments
We gratefully thank Sabine Kaiser (Department of Medicine for Endocrinology, Diabetes and Nutritional Medicine, Charité, Berlin) for the technical assistance. We are deeply thankful to Eddy Rijntjes for support with statistical data analysis, as well as Eva K. Wirth and Carolin S. Höfig for stimulating discussion (Institute for Experimental Endocrinology, Charité, Berlin). This work was supported by grants from the Deutsche Forschungsgemeinschaft (DFG Graduiertenkolleg 1208, TP3 to J.K., and TP8 to C.J.S.) and the Charité-Universitätsmedizin Berlin to J.K., C.J.S., and Z.W. This work in part contributes to the PhD thesis of I.L.
Author Disclosure Statement
All authors have nothing to disclose.