
Abstract
Background:
Cigarette smoking is a risk factor for the development of Graves' ophthalmopathy (GO). In a previous study of gene expression in intraorbital fat, adipocyte-related immediate early genes (IEGs) were overexpressed in patients with GO compared to controls. We investigated whether IEGs are upregulated by smoking, and examined other pathways that may be affected by smoking.
Methods:
Gene expression in intraorbital fat was studied in smokers (n=8) and nonsmokers (n=8) with severe active GO, as well as in subcutaneous fat in thyroid-healthy smokers (n=5) and nonsmokers (n=5) using microarray and real-time polymerase chain reaction (PCR).
Results:
With microarray, eight IEGs were upregulated more than 1.5-fold in smokers compared to nonsmokers with GO. Five were chosen for confirmation and were also overexpressed with real-time PCR. Interleukin-1 beta/IL-1B/(2.3-fold) and interleukin-6/IL-6/(2.4-fold) were upregulated both with microarray and with real-time PCR in smokers with GO compared to nonsmokers. Major histocompatibility complex, class II, DR beta 1/HLA-DRB1/was upregulated with microarray (2.1-fold) and with borderline significance with real-time PCR. None of these genes were upregulated in smokers compared to nonsmokers in subcutaneous fat.
Conclusions:
IEGs, IL-1B, and IL-6 were overexpressed in smokers with severe active GO compared to nonsmokers, suggesting that smoking activates pathways associated with adipogenesis and inflammation. This study underlines the importance of IEGs in the pathogenesis of GO, and provides evidence for possible novel therapeutic interventions in GO. The mechanisms activated by smoking may be shared with other conditions such as rheumatoid arthritis.
Introduction
A
The key pathogenetic processes in GO include adipogenesis, inflammation, excess production of glycosaminoglycans by orbital fibroblasts, and, at later stages, fibrosis. Adipogenesis in vitro includes an initial proliferation phase during which growth arrested preadipocytes re-enter the cell cycle and complete two rounds of cell division known as mitotic clonal expansion (12,13). A terminal differentiation phase follows during which the specific genes that define the adipocyte phenotype are induced (14). During the initial proliferative phase, immediate early genes (IEGs) are induced by adipogenic factors such as growth factors, corticosteroids, and cAMP-increasing agents (15,16). IEGs function as triggers of subsequent transcriptional cascades that lead to the adipocyte phenotype. We have previously shown, in intraorbital tissue, overexpression of IEGs in patients with active severe ophthalmopathy (17) with a decrease in the chronic phase (16). Overexpression of IEGs was not found in chronic lymphedema, another condition involving a pathological increase in the adipose tissue volume (18).
The aim of this study was to investigate whether smoking has an effect on the expression of IEGs in severe active ophthalmopathy and to discover other pathways that are possibly affected by smoking.
Materials and Methods
Collection of patient samples and isolation of RNA
All tissue samples were collected after informed consent and with the approval of the Ethical Review Board of Lund University, Malmö/Lund, Sweden.
For microarray analysis, intraorbital adipose/connective tissue was collected from smokers (n=8) and nonsmokers (n=8) with GO. All patients were in the active phase of GO, and all but one had dysthyroid optic neuropathy (DON). Due to corticosteroid-resistant severe GO, these patients were treated surgically with orbital decompression. These eight smokers and eight nonsmokers with GO, plus seven additional smokers and four additional nonsmokers with GO not included in the microarray and collected in the same manner, were also analyzed by real-time polymerase chain reaction (PCR). For clinical characteristics, see Tables 1 and 2. Patients 1–4 in Table 1 in this study are the same patients as in our earlier study (17) (patients 1, 2, 3, and 5 in Table 1 of the earlier study).
Smokers 1–8 were analyzed with both microarray and real-time polymerase chain reaction (PCR). Smokers 9–15 were only included in the real-time PCR analyses.
GO, Graves' ophthalmopathy.
Nonsmokers 16–23 were analyzed with both microarray and real-time PCR. Nonsmokers 24–27 were only included in the real-time PCR analyses.
For microarray analysis, subcutaneous adipose tissue was obtained from the unaffected extremities of female patients (five smokers and five nonsmokers) undergoing liposuction for chronic lymphedema of the upper or lower extremity. All patients were given a general anesthetic, and before the start of the operation, a biopsy was taken after a 1.5 cm incision without the administration of local analgesia. These five smokers and five nonsmokers, plus two additional smokers and two additional nonsmokers not included in the microarray analysis and collected in the same manner, were also analyzed by real-time PCR. To minimize the degradation of RNA, the biopsies were treated with RNA later (Ambion, Austin, TX) overnight before they were stored at −80°C. RNA was extracted with the RNeasy 96 Kit (QIAGEN, Stockholm, Sweden) according to the manufacturer's instructions. RNA concentration was measured using Nanodrop ND-1000. Since some of the biopsies used were several years old, RNA quality was measured using Agilent 2100 prior to microarray analysis and showed high quality and no signs of degradation. RNA samples of 0.12 μg from each individual were combined into four pools (smokers with GO, nonsmokers with GO, smokers subcutaneous fat, and nonsmokers subcutaneous fat) with eight individual samples in the GO pools and five in the subcutaneous fat pools.
Microarray
The SCIBLU Microarray Resource Center at Lund University performed the gene expression analyses as describes previously (18). Entrez custom chip definition files (
Pathway analysis
The analysis of canonical pathways identified pathways from the IPA library that were most significant with regard to the selected genes. Genes selected on the basis of their overexpression (>twofold) or underexpression (<0.5-fold) were analyzed separately. The association between the data set and the canonical pathway was measured by the ratio of the number of genes from the data set that map to the pathway divided by the total number of genes that map to the canonical pathway. Fisher's exact test was used to calculate a p-value determining the probability that the association between the genes in the data set and the canonical pathway is explained by chance alone. To review the relationship between the up- and downregulated genes, we also looked into the integrated total networks that include these genes, and selected the most significant networks with a cutoff p-value of 0.001.
Ranked differentially expressed genes (>2 and <0.5) were also analyzed using WebGestalt. WebGestalt is a suite of tools for functional enrichment analysis in various biological contexts, and integrates a number of pathway analysis databases such as Gene Ontology, KEGG Pathways, Pathway Commons, and Wikipathways, among others. It compares a user uploaded gene list with genes in predefined functional categories to identify those categories with enriched numbers of user-uploaded genes.
Real-time PCR
Total RNA was reverse transcribed following the instructions for the QuantiTect Rev. Transcription Kit (QIAGEN). PCRs were performed using the ABI PRISM 7900HT sequence detection system (Applied Biosystems, Stockholm, Sweden) according to the manufacturer's specifications as described previously (18) using the TaqMan Universal PCR Master Mix (Applied Biosystems) and the TaqMan Gene Expression Assays (Applied Biosystems). Based on the microarray data, the following relevant genes with smokers/nonsmokers expression ratios of >1.5 and <0.5 in GO and subcutaneous fat respectively were chosen for confirmation by real-time PCR (assay ID in brackets): early growth response 1 (EGR1)/Hs00152928_m1/; dual specificity phophatase 1 (DUSP1)/Hs00610256_m1/; B-cell translocation gene 2 (BTG2)/Hs0019887_m1/; cysteine-rich, angiogenic inducer, 61(CYR61)/Hs00998500_g1/; zinc finger protein 36 (ZFP36)/Hs00185658_m1/; prostaglandin-endoperoxide synthase 2 (PTGS2)/Hs00153133_m1/; major histocompatibility complex, class II, DR beta 1 (HLA-DRB1)/Hs04192464_mH/; interleukin-6 (IL-6)/Hs0017413_m1/; interleukin-1 beta (IL-1B)/Hs00174097_m1/; fatty acid desaturase 1 (FADS1)/Hs00203685_m1/; fatty acid desaturase 2 (FADS2)/Hs00188654_m1/; acetoacetyl-CoA synthetase (AACS)/Hs0022590_m1/; fatty acid synthase (FASN) /Hs01005622_m1/; and fatty acid elongase 6 (ELOVL6)/Hs00907564_m1/. All samples were run in duplicate with a nontemplate control in each batch. Data were analyzed with the standard curve method using cyclophilin A as an endogenous control. Cyclophilin A was chosen as an appropriate endogenous control after testing a subset of samples with the human Endogenous Control Array (4367563) (Applied Biosystems). The results of the real-time PCR were confirmed in at least two separate experiments.
Statistical analysis
The statistical significance of the difference in clinical parameters in smokers and nonsmokers with GO (Table 3) was calculated using the Mann–Whitney rank sum test and chi-square statistics. The statistical significance of the difference in expression among groups detected in real-time PCR was calculated using the Mann–Whitney rank sum test. All statistical tests were performed using SPSS Statistics for Windows v17. (SPSS, Inc., Chicago, IL).
Data on duration of active GO, diagnosis of GD prior to operation, age at operation, and duration of the steroid treatment before surgery are presented as mean (range).
n.s., not significant.
Results
Patients with GO: gene expression in intraorbital fat in smokers compared to nonsmokers
Regarding clinical parameters of all the smokers and nonsmokers included in this study, the two groups differed slightly in a number of clinical parameters. However, these differences were not statistically significant (Table 3). There was a marked difference in gene expression between smokers and nonsmokers with GO. Microarray analysis showed that a total of 103 genes were upregulated more than twofold in smokers compared to nonsmokers. Fifty-four genes were downregulated <0.5-fold in smokers compared to nonsmokers. All up- and downregulated genes in smokers with GO are listed in Supplementary Figures S1 and S2 respectively (Supplementary Data are available online at
Among the genes upregulated in the microarray analysis, five adipocyte-related IEGs were upregulated more than twofold, and another three adipocyte-related IEGs more than 1.5-fold in smokers compared to nonsmokers with GO (Table 4). Out of the eight upregulated IEGs, five were chosen for confirmation with real-time PCR. These five IEGs were previously chosen for confirmation with real-time PCR in our earlier expression study (17). All five IEGs were significantly overexpressed with real-time PCR in both the original eight smokers and eight nonsmokers used for microarray (data not shown), and when including additional smokers and nonsmokers (Fig. 1). We chose to study BTG2 with real-time PCR, even though it was only upregulated 1.4-fold in the microarray analysis. The reason for this decision was that BTG2 belonged to the group of IEGs highly overexpressed in our previous expression study, and that our earlier data showed associations between GD and gene variants in BTG2 (24). Real-time PCR demonstrated that BTG2 was significantly overexpressed in smokers. This was in contrast to the expression in subcutaneous fat, in which none of the IEGs were upregulated in smokers compared to nonsmokers in the microarray analysis (data not shown).
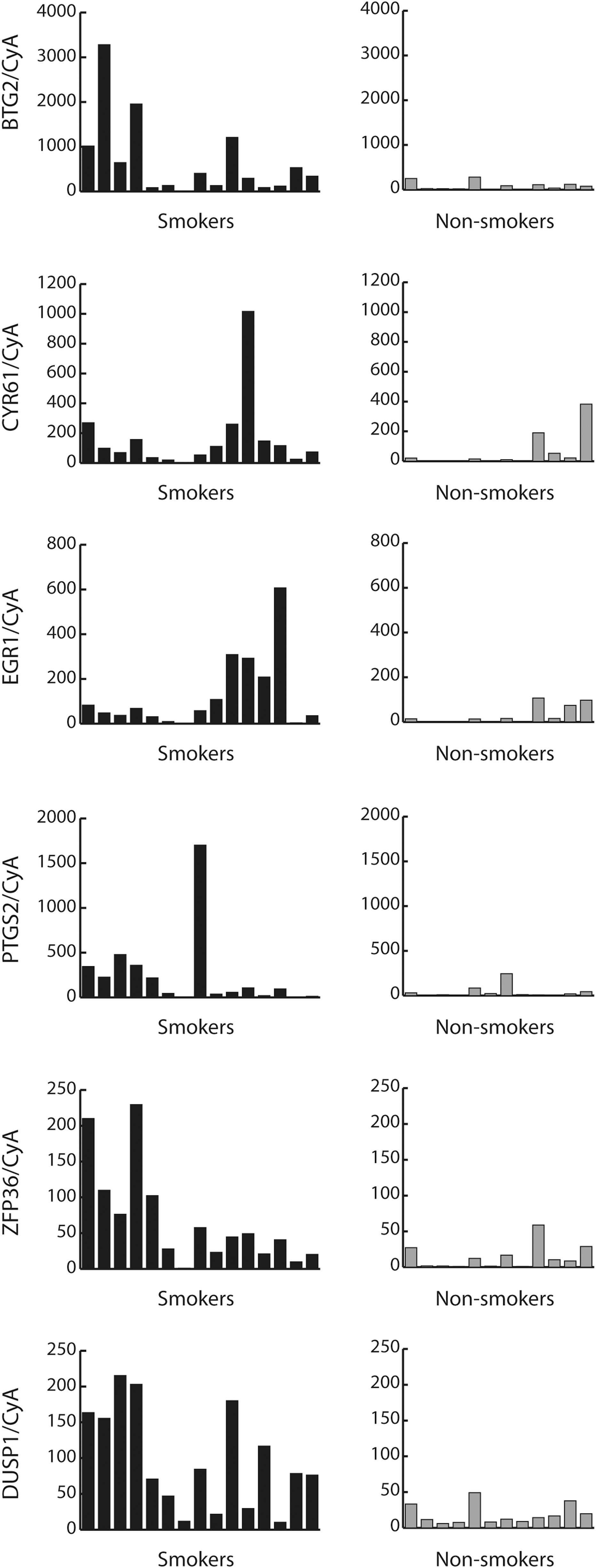
Gene expression of nine immediate early genes (IEGs) in smokers (n=15) and nonsmokers (n=12) with Graves' ophthalmopathy (GO) analyzed with real-time polymerase chain reaction (PCR). All nine genes selected for confirmation with real-time PCR were significantly overexpressed: BTG2 (p=0.002), CYR61 (p=0.009), EGR1 (p=0.03), PTGS2 (p=0.02), ZFP36 (p=0.004), and DUSP1 (p=0.0001).
BTG2, CYR61, EGR1, PTGS2, ZFP36, and DUSP1 were selected for confirmation with real-time PCR.
IEGs, immediate early genes.
IL-1B and IL-6 were found to be upregulated in both the microarray (2.3-fold for IL1-B and 2.4-fold for IL-6) and real-time PCR analyses in smokers with GO compared to nonsmokers. With real-time PCR, the overexpression was significant both when including the original eight smokers and eight nonsmokers used for microarray (data not shown) and when including the additional smokers and nonsmokers (Fig. 2). Microarray analysis showed that HLA-DRB1 was upregulated in smokers with GO compared to nonsmokers (2.1-fold). With real-time PCR, overexpression was observed in smokers with borderline significance (Fig. 3). Upregulation of the interleukin genes and HLA-DRB1 in smokers was not observed in subcutaneous fat in the microarray analysis (data not shown).

Gene expression of IL-1B (p=0.03) and IL-6 (p=0.004) in smokers (n=15) and nonsmokers (n=12) with GO analyzed with real-time PCR.
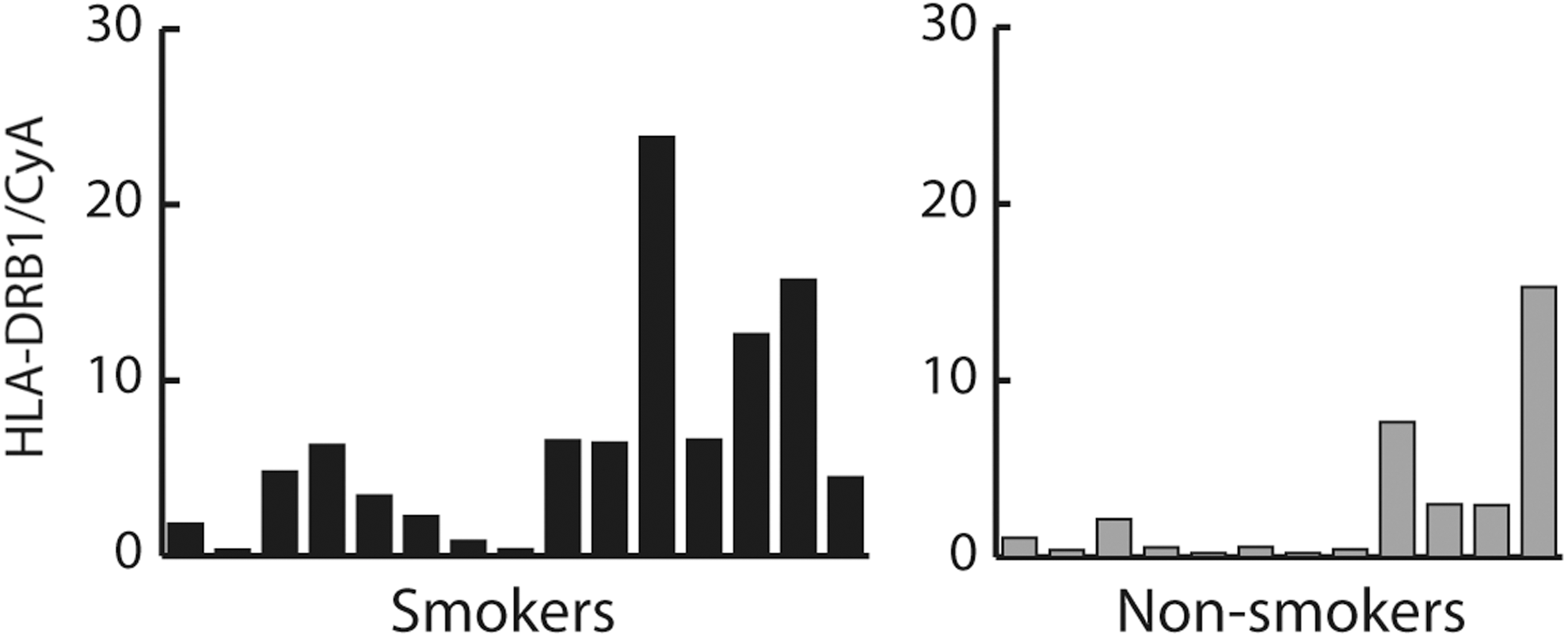
Gene expression of HLA-DRB1 (p=0.05) in smokers (n=15) and nonsmokers (n=12) with GO analyzed with real-time PCR.
Pathway analysis using WebGestalt revealed multiple significantly enriched categories for the genes upregulated more than twofold in smokers. These categories included “Adipogenesis,” “Cytokines and Inflammatory Response,” “TGF-beta receptor signaling,” and “IFN-gamma pathway,” among others.
Pathway analysis with IPA identified “Humoral Immune Response,” “Immune Cell Trafficking,” and “Inflammatory Response” as the networks, and “Immunological Disease” and “Inflammatory Disease” among the biological functions that were strongly associated with the upregulated genes in our data set. The most significant canonical pathways were involved in inflammation and the immune response (TNF, IL-1B), and many of them were involved in the pathogenesis of RA. Some significant pathways were also associated with adipogenesis (CREB1, WNT).
Thyroid-healthy individuals: gene expression in subcutaneous fat in smokers compared to nonsmokers
Microarray analysis revealed that in comparison to intraorbital adipose tissue, a much smaller number of genes were differentially expressed between smokers and nonsmokers in subcutaneous adipose tissue. Eleven genes were upregulated in smokers compared to nonsmokers more than twofold. Out of the eleven genes upregulated more than twofold, none was upregulated more that twofold in GO. However, two genes—FBJ murine osteosarcoma viral oncogene homolog B (FOSB) and FBJ murine osteosarcoma viral oncogene homolog (FOS)—were upregulated 1.6-fold and 1.5 fold respectively in GO. Only two genes were downregulated in smokers <0.5-fold. All up- and downregulated genes in subcutaneous fat in smokers are listed in Supplementary Figures S3 and S4. Among the genes upregulated in smokers, a group of genes with functions in fatty acid metabolism was identified, including FADS2, FADS1, AACS, ELOVL6, and FASN. Significant overexpression of these genes, except for ELOVL6, was confirmed by real-time PCR (Fig. 4). HLA-DRB1, which was upregulated in smokers with GO compared to nonsmokers with GO, was downregulated in subcutaneous fat in smokers compared to nonsmokers when analyzed by microarrays (0.3-fold) and real-time PCR (Fig. 4).

Gene expression of five genes with functions in fatty acid metabolism and of HLA-DRB1 in subcutaneous fat of healthy smokers (n=7) and nonsmokers (n=7) with real-time PCR. All fatty acid genes except for ELOVL6 selected for confirmation with real-time PCR were significantly overexpressed: FADS1 (p=0.01), FADS2 (p=0.001), AACS (p=0.03), FASN (p=0.05), and ELOVL6 (p=0.07). HLA-DRB1 was significantly downregulated (p=0.004).
Pathway analysis with WebGestalt and IPA identified “Lipid metabolism” as the biological function and network strongly associated with the upregulated genes in our data set, with fatty acid metabolism and cholesterol metabolism being the top significant canonical pathways. Comparing the results of IPA in GO and subcutaneous fat, pathways associated with “Synthesis of lipid” and “Differentiation of connective tissue” were upregulated in both conditions.
Discussion
This study compared gene expression in intraorbital fat in smokers and nonsmokers with severe active GO. The principal finding was the overexpression of IEGs, IL-1B, and IL-6 in smokers. None of these changes were observed in subcutaneous fat in smokers without thyroid disease compared to nonsmokers.
We have previously shown overexpression of IEGs in intraorbital tissue in patients with severe active ophthalmopathy (17) compared to healthy controls. In a genetic study, single nucleotide polymorphisms (SNPs) in several IEGs were associated with GD and/or GO (24). IEGs are a group of genes known to be expressed in differentiating preadipocytes (25,26). The expression of IEGs during adipogenesis initiates the mitotic clonal expansion phase, which is followed by a second growth arrest and then the terminal differentiation phase, leading to the mature adipocyte phenotype (14). In the present study, overexpression of nine adipocyte-related IEGs, including CYR61 and EGR1, in smokers with active ophthalmopathy compared to nonsmokers supports the hypothesis that smoking activates pathways associated with adipogenesis. In addition, the pathways analysis showed a significant overrepresentation of genes involved in adipogenesis.
This hypothesis is further supported by findings from other studies. In the study by Cawood et al. (27), cigarette smoke extract (CSE) increased adipogenesis in a dose-related manner, as did IL-1. The effects of CSE and IL-1 on adipogenesis were synergistic. We have previously shown that IL-1 was upregulated in active ophthalmopathy compared to thyroid-healthy controls (17), and IL-1 is known to be a CYR61-responsive gene (28). In the present study, both CYR61 and IL-1B were further overexpressed in smokers with active GO. Apart from IL-1B, IL-6 was also found to be upregulated in this study. IL-1 and IL-6 are cytokines with pathogenic functions in the inflammatory reaction in orbital tissue in GO. Binding of CD40 on orbital fibroblasts and CD40-ligand on CD4+ T cells results in production of high levels of IL-1 and IL-6, which augments B-cell maturation and antibody production (29). Therefore, smoking may influence inflammatory pathways, which was also revealed by the pathways analysis of the microarray data.
In a recent study, dermal fibroblasts exposed to cigarette smoke showed upregulation of the IEGs EGR-1 and CYR61 (30). Both of these genes were overexpressed in smokers compared to nonsmokers in the present study. CYR61 is a multifunctional gene with roles in adipogenesis, inflammation, cell proliferation, extracellular matrix production, and fibrosis (31,32). Thus, there are several mechanisms by which CYR61 could contribute to the pathogenesis of GO. Previous studies have implicated CYR61 in the pathogenesis of inflammatory diseases, including RA. Microarray analysis of B cells from monozygotic twins revealed a significantly higher expression of CYR61 in the twin with RA compared to the healthy twin (33). CYR61 has also been shown to be abundantly expressed in synovial tissue of monozygotic twins with RA (33) and plays a critical role in the proliferation of rheumatoid arthritis synovial fibroblasts (34). In a recent study, Kok et al. demonstrated that proinflammatory cytokines, including IL-1B, IL-6, and TNF-alpha, stimulated CYR61 expression in osteoblastic cells, and that this action was inhibited by simvastatin. Moreover, in a mouse model of inflammatory arthritis, CYR61 expression in osteoblasts correlated with disease activity, and intraarticularly injected simvastatin alleviated arthritis. The authors concluded that simvastatin might have potential as a therapeutic agent for inflammatory arthritis (35). If these mechanisms are shared between RA and GO, it is tempting to speculate that simvastatin could represent a promising therapeutic agent in GO. However, in vitro studies on CYR61 expression in orbital fibroblasts and the effects of simvastatin are required first.
HLA-DRB1 alleles are known to be associated with GD (36,37). In RA, HLA-DRB1 and smoking interact by increasing the risk of anti-citrullinated protein antibody-positive RA (38 –40). In our study, overexpression of HLA-DRB1 was observed by microarray analysis. However, when analyzed by real-time PCR, the overexpression reached borderline significance, most likely due to the inclusion of a limited number of individuals. Overexpression of HLA-DRB1 in smokers with GO compared to nonsmokers could suggest that similar mechanisms of interaction with smoking, which exist in RA, could be present in GO/GD.
We are aware of the limitations of this study. Our findings are based on a limited number of individuals included in the study, and the groups of smokers and nonsmokers differed slightly with regards to clinical parameters such as sex, age, and duration of GD and GO. Therefore, some caution must be exercised when interpreting the results, as factors other than smoking may have contributed to the observed differences in expression. However, interestingly, although the smokers used for the microarray analysis had, on average, been treated longer with corticosteroids, pathways associated with inflammation were upregulated in smokers compared to nonsmokers with GO. Another limitation is the use of subcutaneous adipose tissue as a control, as it is well known that orbital fibroblasts/preadipocytes differ from fibroblasts from other sites of the body. Hence, some of the differences observed between the expression patterns in the orbital and subcutaneous tissues might be attributable to the difference in type of adipose tissue. Unfortunately, the perfect control tissue—intraorbital fat from healthy smokers and nonsmokers—was not available for this study.
In conclusion, IEGs, IL-1B, and IL-6 were overexpressed in smokers with severe active GO compared to nonsmokers, suggesting that smoking in GO activates pathways associated with adipogenesis and the inflammatory response. This study underlines the importance of IEGs in the pathogenesis of GO and provides evidence for possible future therapeutic interventions in GO. The mechanisms activated by smoking may be shared with other conditions such as rheumatoid arthritis.
Footnotes
Acknowledgments
The study was supported by grants from Svenska Läkarsällskapet, Svenska Endokrinologföreningen, research funds of Malmö University Hospital, the Faculty of Medicine at Lund University, and the Skåne Research Foundation.
Author Disclosure Statement
The authors have nothing to disclose.