
Abstract
Background:
Expression of the small calcium-binding protein S100A4 is associated with poor prognosis in patients with thyroid cancer (TC). The authors have previously shown that S100A4 is a target for relaxin and insulin-like peptide 3 signaling in TC cells and that S100A4 is secreted from human TC cells. Although the pro-migratory role of intracellular S100A4 in binding to non-muscle myosin is well known, this study investigated here whether extracellular S100A4 contributes to TC migration.
Methods:
Human cell lines of follicular, papillary, and undifferentiated thyroid cancer, primary patient TC cells, and TC tissues were utilized to discover the presence of the receptor of advanced glycation end products (RAGE) in TC cells and TC tissues. Fluorescence imaging, protein pull-down assays, Western blot, siRNA protein silencing, small GTPase inhibitors, cell proliferation, and cell migration assays were used to investigate the interaction of extracellular S100A4 with RAGE in promoting a TC migratory response.
Results:
It was demonstrated that RAGE served as receptor for extracellular S100A4 mediating cell migration in TC cells. The RAGE-mediated increase in cell migration was dependent on the intracellular RAGE signaling partner diaphanous-1 (Dia-1) and involved the activation of the small GTPases Cdc42 and RhoA. Although extracellular S100A4 consistently activated ERK signaling in TC cells, it was shown that ERK signaling was not mediated by RAGE and not essential for the migratory response in TC cells.
Conclusion:
The data have identified the RAGE/Dia-1 signaling system as a mediator for the pro-migratory response of extracellular S100A4 in human TC. Thus, therapeutic targeting of the RAGE/Dia-1/small GTPases signaling may successfully reduce local invasion and metastasis in TC.
Introduction
T
The receptor of advanced glycation end products (RAGE) is a pattern recognition receptor (20,21) and mediates the actions of several molecules of the danger-associated molecular pattern molecules (DAMPs) (22,23), such as HMGB1 in enhancing inflammatory and survival mechanisms in several human pathologies (23 –25). RAGE is expressed in several human cancer entities (26 –29), but its expression in TC is not known. S100A4 binds to RAGE in the dimeric and oligomeric form in vitro (30). Interaction of extracellular S100A4 with RAGE was recently demonstrated in chondrocytes of osteoarthritis patients (31), in endothelial cells (32), and in human colorectal cancer cells (33), and resulted in ERK1/2 phosphorylation. In a xenograft model of human prostate cancer, S100A4 acts via RAGE to promote tumor growth (34).
In this study, the expression of RAGE in human TC tissues and TC cells isolated from surgical tissue specimen was identified, and the interaction of recombinant human S100A4 (rhS100A4) with RAGE in TC cells with endogenous RAGE expression was demonstrated. It was shown that the motility promoting effect of S100A4 was dependent on RAGE. S100A4-RAGE signaling was mediated through its cytoplasmic interaction partner diaphanous-1 (Dia-1) and was dependent on the small GTPases Rac-1/Cdc42 and RhoA.
Materials and Methods
Cell culture
Human TC cell lines FTC133, FTC236, C643, TPC-1, and B-CPAP were cultured in Dulbecco's modified Eagle's medium/Nutrient F-12 Ham 1:1 (DME/F12, Hyclone; Thermo Scientific, Waltham, MA) with 10% fetal bovine serum (FBS; Invitrogen, Burlington, Canada) and passaged every three to four days to maintain a proliferative culture.
RAGE knockdown in TC cells through transient transfection
The human TC cell lines FTC 236 and C643 were seeded at a density of 1×105 cells/well and treated with small interference RNA (siRNA) for RAGE (#sc-36374) or scrambled siRNA (#sc-37007; both Santa Cruz Biotechnologies, Santa Cruz, CA) at a concentration of 80 nM using SilentFect (#170-3361; Bio-Rad, Mississauga, Canada).
Production of stable Dia-1 KO clones
FTC 236 and C643 cells were seeded at a density of 5×104 cells/well and treated with 15 μg/mL polybrene for 2 h followed by transduction of lentiviral Dia-1 shRNA particles or control shRNA, respectively (both Santa Cruz Biotechnologies; sc-35190-V, sc-108080). Stable clones were selected using puromycin at 4 μg/mL (#P9620; Sigma, St Louis, MO).
Primary human thyroid cell culture
Human thyroid primary cells were isolated from surgical tissues obtained from patients undergoing hemi-thyroidectomy at St. Boniface General Hospital with ethics approval from the University of Manitoba Human Ethics Board. TC cells were isolated using 500 μg/mL collagenase and 10 μg/mL DNAse (both Sigma) for tissue digestion, filtered through a 40 μm cell strainer (BD Bioscience, San Diego, CA) and subjected to red blood cell lysis. Primary cells from TC tissues were cultured in DME/F12 1:1 supplemented with 10% FBS and 1× penicillin/streptomycin (Gibco, Grand Island, NY).
Polymerase chain reaction
The cDNAs were generated using random primers. Transcripts were polymerase chain reaction (PCR) amplified from selected TC cell lines and primary TC cells. The PCR cycle consisted of an initial denaturation step at 95°C for 3 min, a denaturation step at 95°C for 1 min, annealing at specific temperature for 1 min, and a final elongation step at 72°C for 2 min. Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was used as a control to assess the quantity of cDNA used per PCR reaction. The specific oligonucleotide primer sequences for the detection of S100A4, RAGE, Toll-like receptor 4 (TLR4), and GAPDH and their respective melting temperatures are listed in Table 1.
Tm, melting temperature; PCR, polymerase chain reaction; RAGE, receptor of advanced glycation end products; TLR4, Toll-like receptor 4; GAPDH, glyceraldehyde-3-phosphate dehydrogenase.
Recombinant proteins, peptides, and inhibitors
Recombinant human S100A4 (rh S100A4; #GWB-P1718B) of Escherichia coli origin with N-terminal His-tag was purchased from GenWay Biotech (San Diego, CA). Advanced glycation end product–bovine serum albumin (AGE-BSA) was purchased from BioVision, Inc. (Milpitas, CA; #2221-10). The rhS100A4 and the AGE-BSA had been tested and were confirmed LPS-free. Inhibitors for Rac-1/Cdc42 (ML141; #217708) and Rho kinase (H-1152P, #555552) were both purchased from Calbiochem (Cambridge, MA). The MEK inhibitor U0126, the EGFR inhibitor AG1478, and EGF were purchased from Sigma.
RAGE pull-down assay
Dynabeads® (Antibody Coupling Kit 143.11D; Invitrogen) were coupled with rhS100A4 (4137S4; R&D Systems, Minneapolis, MN) or BSA, respectively, according to the manufacturers' protocol, and blocked with 2% BSA in phosphate-buffered saline (PBS). B-CPAP cells (3×107) were lysed in RIPA buffer supplemented with protease inhibitor cocktail (Roche, Mississauga, Canada). The lysate was split into three equal volumes. One volume of lysate was supplemented with 5 μg RAGE antibody (H-300; Santa Cruz Biotechnologies) before the beads were added. The lysates were incubated with 10 μg of coupled beads overnight at 4°C under constant rotation. The beads were washed three times with PBS and subjected to Western blotting and immunostaining of membranes with RAGE-specific antibody (A11; Santa Cruz Biotechnologies).
Western blotting
The preparation of protein lysates and polyacrylamide gels for Western blotting was performed as described previously (35). For the detection of RAGE, Dia-1 and S100A4, an anti-RAGE rabbit polyclonal antibody (1:1000) and an anti-Dia-1 mouse monoclonal antibody (1:1000; both Santa Cruz Biotechnologies), and an anti-S100A4 rabbit monoclonal antibody (1:1000; Cell Signaling Technology, Danvers, MA) were probed overnight at 4°C, respectively. Horseradish peroxidase (HRP) coupled goat anti-rabbit (Cell Signaling Technology) and goat anti-mouse (Sigma) secondary antibodies were used (both at 1:5000). For the detection of ERK1/2 signaling intermediates, antibodies for total ERK1/2 and phospho-ERK1/2 were purchased (Cell Signaling Technology), and blots were developed by autoradiography using ECL (Pierce, Nepean, Canada). Densitometry analysis of the X-ray films was done using the spot denso tool in Fluorchem software.
Immunofluorescence
TC cells were fixed with 3.7% formaldehyde, permeabilized with 0.01% Triton-X100 for 15 min, and blocked with 5% normal goat serum (NGS) for 1 h at room temperature (RT). Cells were incubated overnight at 4°C with the primary antibodies for RAGE (1:100), Dia-1 (1:100) (both Santa Cruz), and Pax-8 (1: 100; Biocare Medical, Inc., Concord, CA) and corresponding isotype control IgG immunoglobulins diluted in 5% NGS (Sigma). Slides were washed three times for 5 min and incubated with AlexaFluor conjugated goat anti rabbit (1:500; #A11012) and goat anti mouse (1: 1000; #A11005; both Invitrogen) secondary antibodies for 1 h at RT and washed three times for 5 min. 4′,6-diamidino-2-phenylindole (DAPI) was used for nuclear counterstain, and cells were covered in anti-fade mounting medium (Life Technologies, Burlington, Canada).
Immunohistochemistry
Human thyroid tissues of normal thyroid and TC were collected from surgical patient specimens at the Department of Surgery, University of Halle-Wittenberg, and formalin-fixed paraffin-embedded tissue sections (5 μm) were used for this study. This study was approved by the ethical committee of the Martin-Luther-University, Faculty of Medicine. The human TC tissue micro array (TMA) was obtained from US Biomax (Rockville, MD; TH208). It comprised of a total of 208 cores of 1 mm diameter and 60 cases with triplicate cores per case, including nine normal thyroid tissues. Dewaxing, antigen retrieval, and immunohistochemistry were performed as described previously (15). Sections were probed with the anti-RAGE rabbit primary antibody (1:100) or an isotype control antibody (Sigma) at equivalent protein concentrations overnight at 4°C. Following incubation with a biotin-conjugated goat anti-rabbit secondary antibody (1:500) and avidin-biotin complexes (Vectstain ABC kit, #PK-6105), specific staining was visualized using 3,3′-diaminobenzidine substrate (DAB). Sections were imaged using a Zeiss M1 bright field microscope and a 20× objective.
Bromodeoxyuridine cell proliferation assay
Human TC cells were seeded at a density of 5×103 cells/well, serum starved for 24 h, and treated with rhS100A4 (100 nM, 250 nM, 500 nM). After 24 h of treatment, a calorimetric enzyme-linked immunosorbent assay (ELISA) was done using the Bromodeoxyuridine (BrdU) cell proliferation kit (Roche). The absorbance was measured at 450 nm using an ELISA reader (Perkin Elmer, Boston, MA).
Cell viability (WST-1) assay
Human TC cells were seeded at a density of 5×103 cells/well, serum starved, and treated with rhS100A4 (100 nM, 250 nM, 500 nM). After 24 h of treatment, WST-1 reagent was added to the cells and incubated at 37°C for 4 h. The formation of water-soluble formazan was measured at 450 nm.
Migration assay
Cells were cultured in 1% fetal calf serum medium and seeded onto a transwell filter membrane with 8 μm pore size (Costar, Corning, NY) at a density of 1×104 cells/well. TC cells were treated with rhS100A4 (100 nM, 250 nM, 500 nM) and AGE-BSA (10 μg/mL) for 24 h. For Rac-1/Cdc42 and RhoA inhibition experiments, cells were treated with the respective inhibitors at 10 μM for 24 h and then seeded on the filters along with the inhibitors. For MEK inhibition experiments, cells were treated with U0126 at 10 μM for 1 h prior to treatment with rhS100A4. After 24 h of migration, cells were washed, fixed with methanol, and stained with 0.1% toluidine blue dye (Sigma). Cells that had migrated to the underside of the filter were imaged and counted as described (15).
siRNA RAGE silencing
RAGE was silenced in transient transfections using SilentFect transfection reagent (BioRad) with RAGE siRNA and compared to non-silencing (scrambled) siRNA (both SantaCruz). At 48 h following siRNA treatment, cells were used for migration assays.
Cdc42 and RhoA pathway inhibition assays
Cells were serum starved and treated for 24 h with a Cdc42/Rac-1 inhibitor at 10 μM (ML141) and a Rho kinase inhibitor at 10 μM (H-1152P; both Calbiochem). Cells were lysed, and active GTPase proteins were pulled down using PAK-PBD (to bind GTP-Cdc42) and Rhotekin-RBD beads (to bind GTP-RhoA) using the RhoA/Rac1/Cdc42 Activation Assay Combo Biochem Kit (BK030; Cytoskeleton, Denver, CO). The immunoprecipitated active small GTPases were identified by Western blot using specific antibodies for Cdc42 and RhoA provided with the kit.
Statistical analysis
Statistical analyses were done using GraphPad Prism software. The differences between the groups were calculated using one way analysis of variance, and the confidence interval in each analyses was set at 95% so the comparisons having a p-value of <0.05 were considered significant.
Results
RAGE is expressed in human TC cells and tissues
The study demonstrated the expression of RAGE transcripts (Fig. 1A) and RAGE protein (Fig. 1B) utilizing human TC cell lines representing the three histological types of human TC: FTC (FTC133, FTC 236), PTC (TPC-1 and B-CPAP), and UTC (C643). Expression of S100A4 was detected in FTC133 but was negligible in the other TC cell lines (Fig. 1B and C). S100A4-negative TC cells but positive for endogenous RAGE were used as the experimental model to investigate the role of extracellular S100A4 in TC. Primary TC cells isolated from surgical patient tissues (Supplementary Fig. S1A; Supplementary Data are available online at
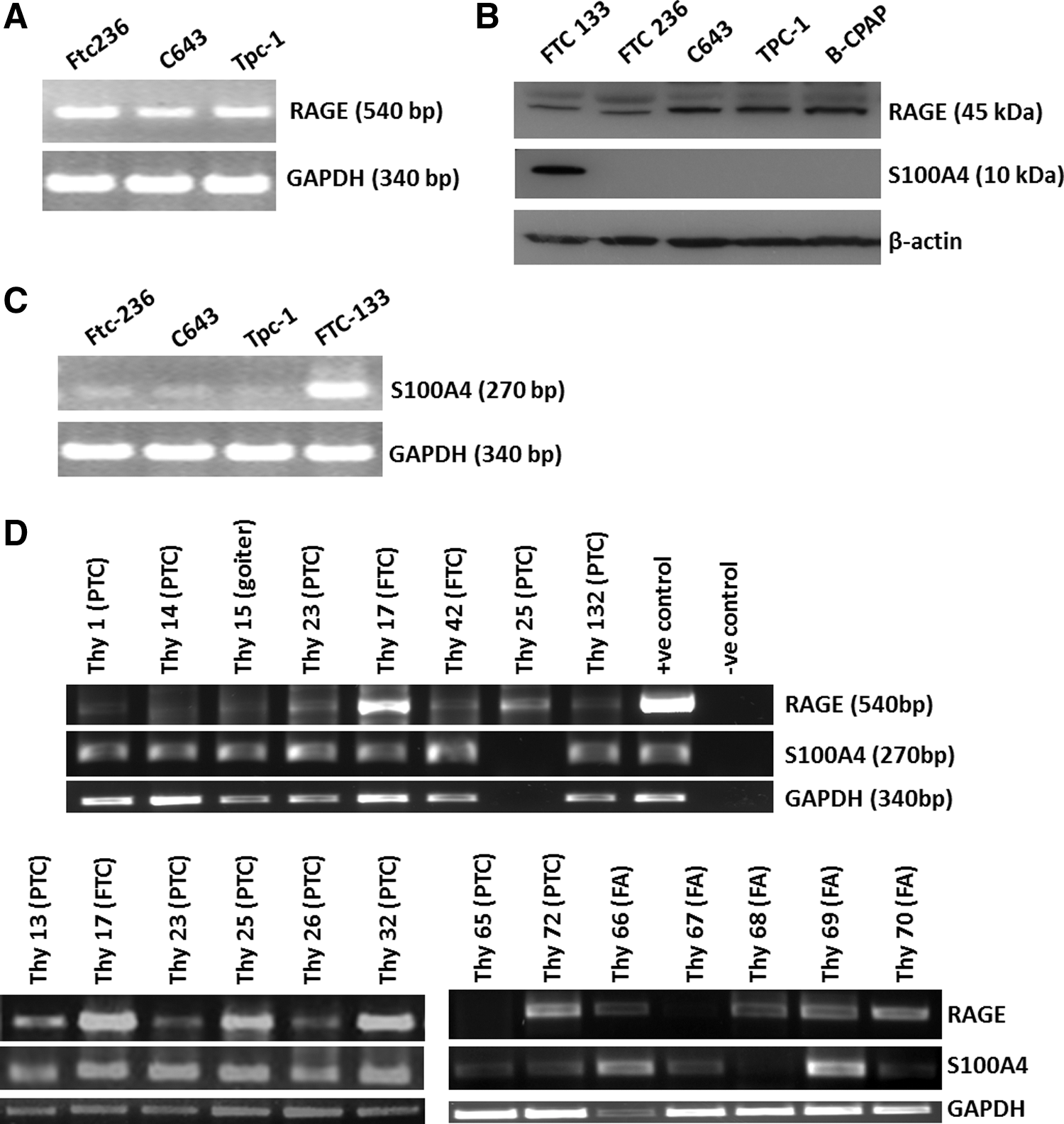
Receptor of advanced glycation end products (RAGE) is expressed in established and primary thyroid cancer (TC) cells. RAGE transcripts (
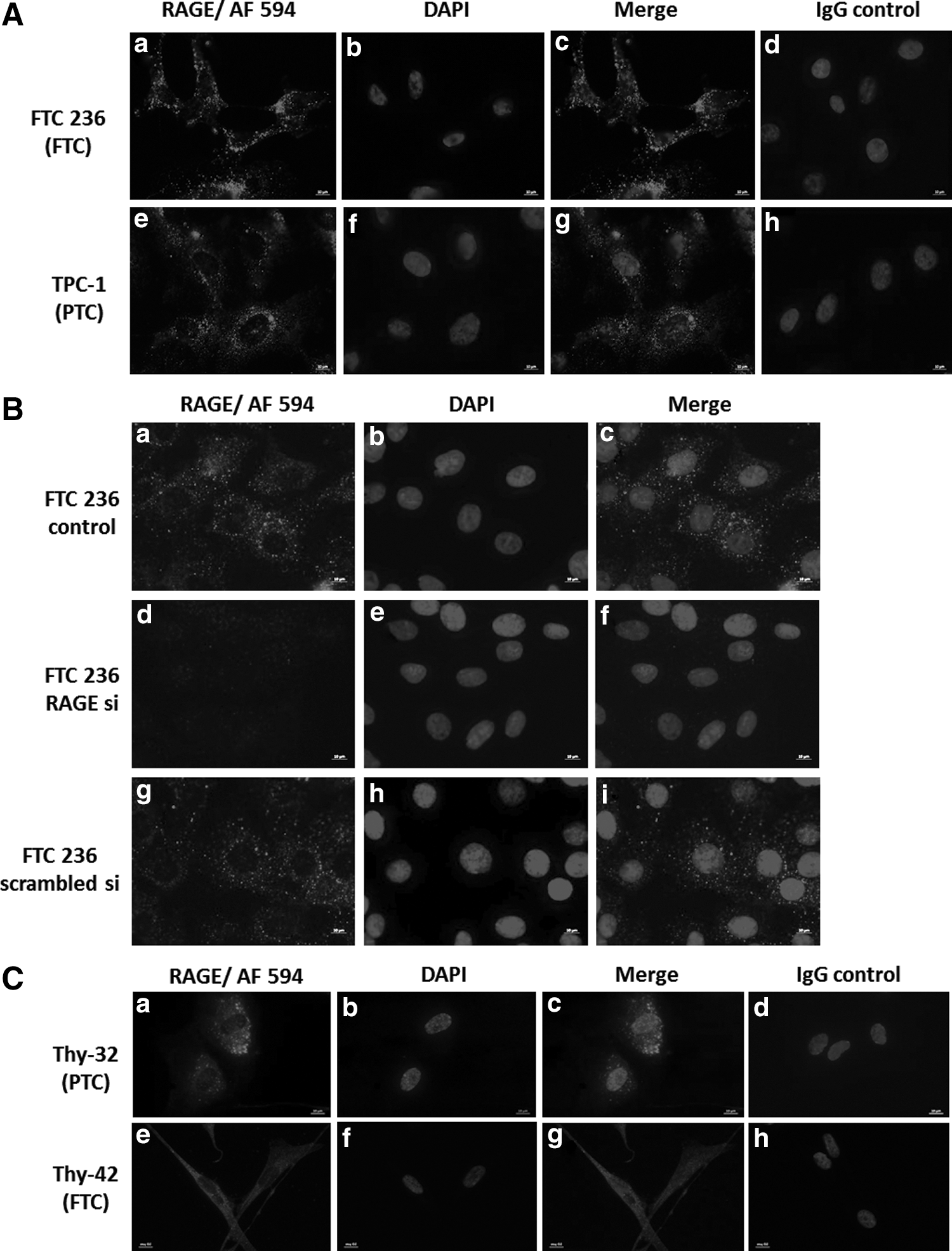
RAGE protein detection by immunofluorescence in TC cells. RAGE protein was detected by immunofluorescence in all human thyroid carcinoma cell lines as shown here for FTC-236 (
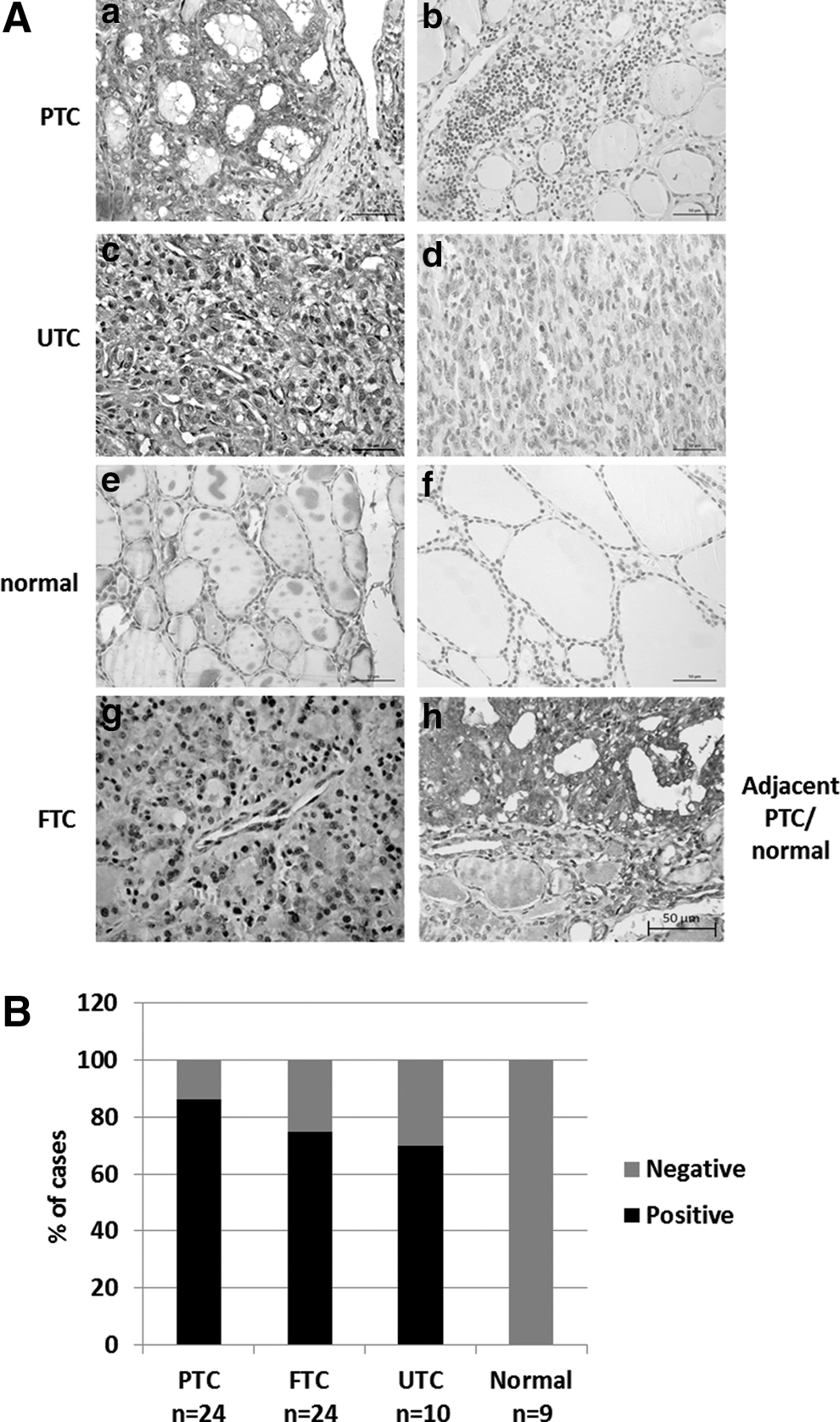
RAGE is expressed in neoplastic but not in normal thyroid tissues. Immunohistochemistry (IHC) was performed on sections from paraffin-embedded human thyroid tissues. Sections of human FTC, PTC, and UTC probed with the RAGE antibody showed positive staining in neoplastic thyroid cells, as shown here for PTC (
Extracellular S100A4 binds to RAGE to promote cancer cell migration
S100A4 has been described as a RAGE ligand in other human cell types (36). To determine whether exogenous S100A4 binds to RAGE in TC cells, S100A4-coupled beads were utilized to pull down endogenous RAGE protein in human B-CPAP PTC cells. RhS100A4, but not BSA, was able to pull down RAGE protein, suggesting that S100A4 binds to RAGE (Fig. 4). RhS100A4 did not reduce cell viability (Fig. 5A) and failed to increase cell proliferation (Fig. 5B) in TC cells. However, rhS100A4 at 500 nM significantly increased cell migration (Fig. 5C) in TC cells, as demonstrated by two-chamber migration assays. The known physiological ligand of RAGE, advanced glycation end products (AGE), was used as a positive control and elicited a pro-migratory response in TC cells (Fig. 5D).

Endogenous RAGE interacts with rhS100A4 in TC cells. Recombinant human (rh)S100A4 was covalently coupled to Dynabeads for immunoprecipitation. BSA-coupled beads were used as negative control. S100A4-coupled beads immunoprecipitated RAGE protein when incubated with total cell lysate of the human PTC cell line B-CPAP. When a RAGE-blocking antibody was added to the cell lysate, S100A4 failed to immunoprecipitate RAGE protein.

Exogenous S100A4 is not mitogenic but increases cell migration. Extracellular rhS100A4 did not cause cytotoxic reactions in the concentrations used, as shown by WST-1 assays (
S100A4-induced TC cell migration is dependent on RAGE and Dia-1
RAGE expression in TC cells was silenced using RAGE siRNA and scrambled siRNA treatment was used as a control to maintain endogenous RAGE expression (Fig. 6A). Upon specific RAGE knockdown, TC cells failed to respond to rhS100A4 or AGE with increased migration (Fig. 6B), whereas scrambled siRNA treated RAGE-positive TC control cells showed a significant pro-migratory response to both rhS100A4 and AGE (Fig. 6B). S100A4 has recently been shown to signal via TLR4 in mouse mammary carcinoma cells (37). This study sought to detect the presence of TLR4 in TC cells as a potential mediator for S100A4-induced migration. RAGE silencing did not affect the expression levels of TLR4. Thus, binding of S100A4 to TLR4 did not contribute to the S100A4-induced migratory response in TC (Fig. 6A). Dia-1 was identified as cytoplasmic interaction partner for RAGE, mediating increased cell migration toward modified human serum albumin in rat glioma cells (38). Dia-1 protein expression was detected by Western blot (Fig. 7A) and the presence of Dia-1 in the cytoplasm of TC cells was demonstrated by immunofluorescence (Fig. 7B). Stable Dia-1 knockdown clones of FTC236 and C643 cells were established using lentiviral shRNA silencing (Fig. 7C), and it was discovered that Dia-1 silencing completely abolished the pro-migratory response of rhS100A4 (Fig. 7D). Dia-1 silencing did not change the expression of RAGE protein in Dia-1 knockout clones (Fig. 7C), confirming that S100A4-induced and RAGE-mediated cell migration was dependent on the cytoplasmic RAGE interaction partner Dia-1 in TC cells.
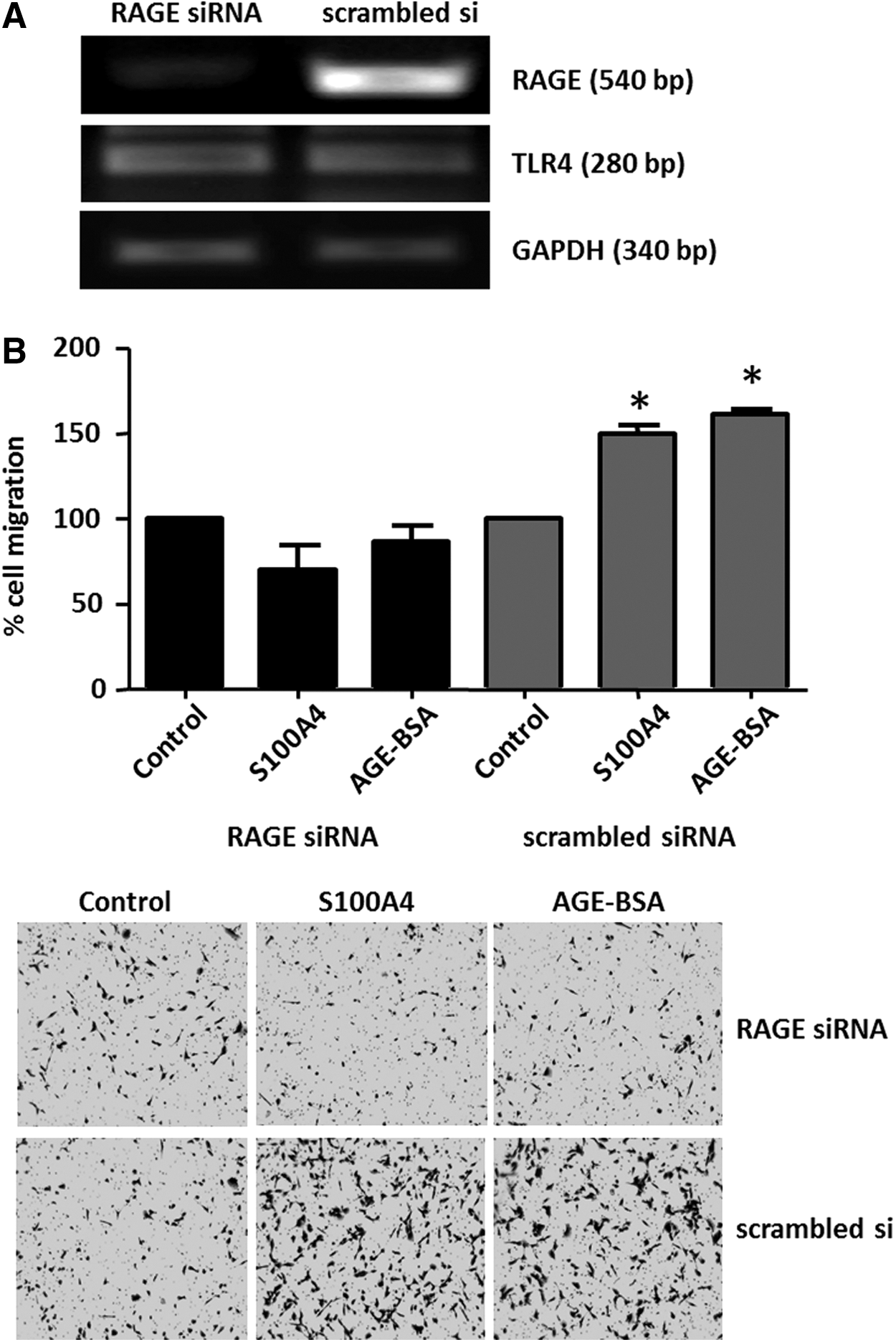
S100A4-induced cell migration in TC cells is RAGE-dependent. Reverse transcription polymerase chain reaction (RT-PCR) detection of RAGE and TLR4 transcripts is shown in FTC-236 cells following 48 h of RAGE knockdown using transient transfections with RAGE siRNA. Non-silencing “scrambled” siRNA served as a negative control (
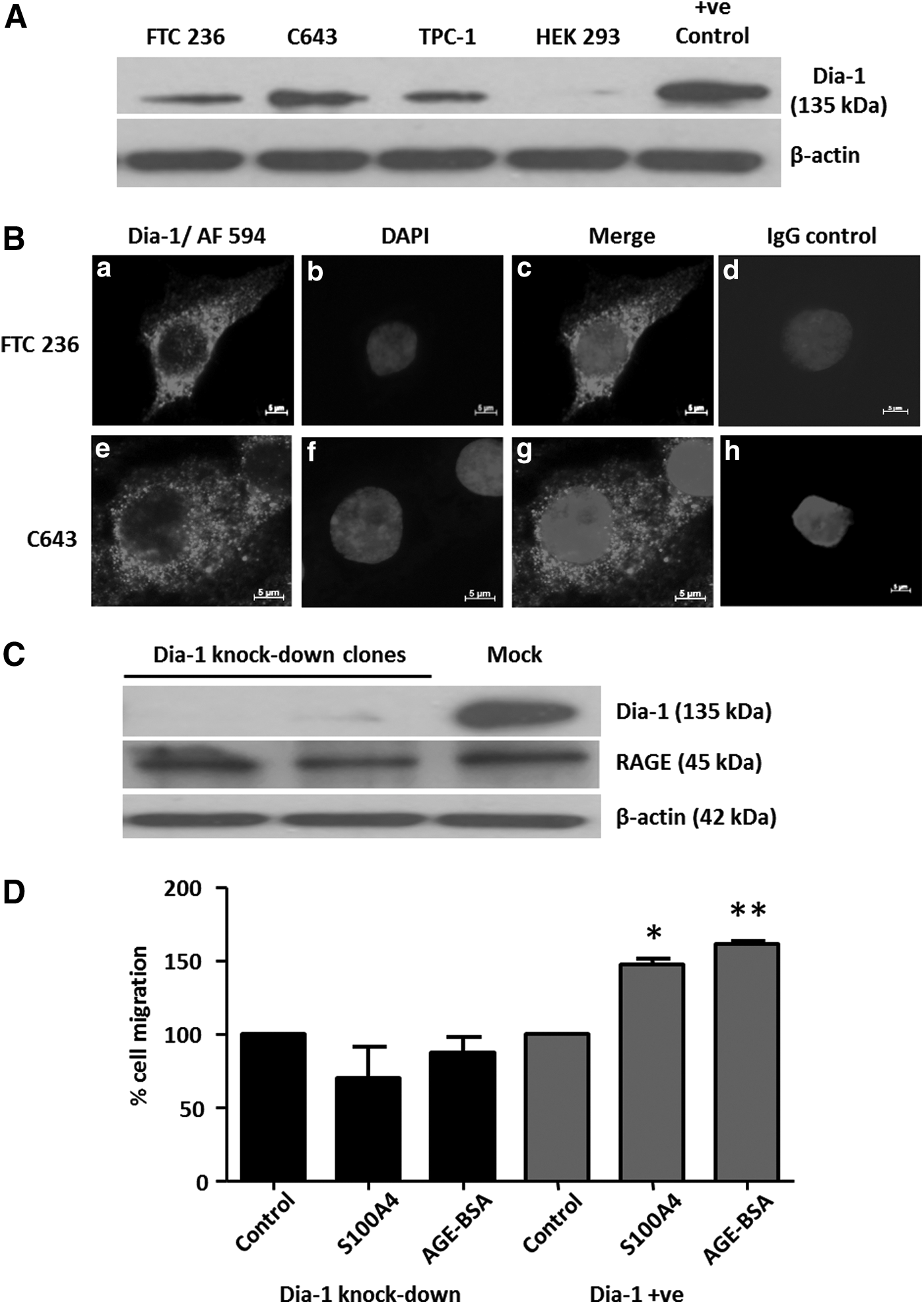
Cell migration by S100A4/RAGE is mediated by Diaphanous-1 (Dia-1) in TC cells. Human TC cells expressed the 135 kDa Dia-1 protein, as shown by Western blot (
S100A4/RAGE-mediated increased cell migration involves Cdc42 and RhoA kinase
This study investigated whether activation of the S100A4-RAGE/Dia-1 signaling pathway in TC cells included the small GTPases Cdc42 and RhoA as potential downstream RAGE/Dia-1 targets. S100A4 increased the active GTP-bound fractions of both RhoA and Cdc42 (Fig. 8A). The reduction of RAGE protein to approximately 50% of normal cellular levels coincided with a markedly diminished fraction of active GTP-RhoA but did not affect Cdc42 activation (Fig. 8B). To determine whether the S100A4-induced cell migration was dependent on the activation of small GTPases, the Rac-1/ Cdc42 inhibitor ML141 and the Rho kinase inhibitor H-1152P were employed at non-toxic concentrations of 10 μM each (Fig. 8C). Reduced levels of the GTP-bound RhoA and Cdc-42 indicated the successful inhibition of their GTPase activity (Fig. 8D). Inhibition of Cdc42 activation by ML141 almost completely abolished the ability of S100A4 to induce TC cell migration, whereas treatment with the Rho kinase inhibitor H-1152P caused an approximately 50% reduction of TC migration (Fig. 8E). DMSO solvent had no effect on migration (Fig. 8E). Thus, the active forms of both Cdc42 and RhoA were identified as downstream mediators of the S100A4-RAGE/Dia-1 pathway leading to enhanced TC migration.

S100A4/RAGE-induced cell migration required Cdc42 and RhoA. As shown by GTP pull-down assays, exogenous S100A4 increased the active GTP-bound fraction of RhoA and Cdc42 in C643 cells (
S100A4-induced p44/p42 MAPK signaling is independent of RAGE
Extracellular S100A4 induced a robust ERK1/2 phosphorylation in TC cells at concentrations of 250 nM of S100A4 (Fig. 9A), and the MEK inhibitor U0126 prevented this ERK1/2 phosphorylation (Fig. 9B). In cells treated with RAGE siRNA, rhS100A4 was still able to induce ERK1/2 phosphorylation (Fig. 9C), indicating a RAGE independent S100A4 signaling event. In Dia-1 knockdown cells, rhS100A4 mediated ERK1/2 phosphorylation was detectable but delayed compared to parental cells (Fig. 9D). Treatment with the MEK inhibitor U0126 at 10 μM failed to block the pro-migratory response in TC cells treated with exogenous rhS100A4, suggesting that ERK activation was not directly linked to cell migration in TC.

S100A4-induced ERK1/2 activation is independent of RAGE. Recombinant human S100A4 at 250 nM resulted in the phosphorylation of the p44/p42 MAPK (ERK1/2) in FTC, PTC, and UTC cells, as shown here for C-643 (
Discussion
This study reports the expression of RAGE in human TC tissues and in human primary and established TC cells. The study shows that the induction of cell motility in human thyroid carcinoma cells induced by extracellular S100A4 was RAGE-dependent and involved the cytoplasmic RAGE-interaction partner Dia-1 and the small GTPases of the RhoA/Cdc42/Rac-1 family. The data demonstrated that in TC cells, the small GTPases Cdc42 and RhoA, but not the MAP kinases ERK1/2, were activated by exogenous S100A4 downstream of RAGE/Dia-1 to increase cell motility.
The authors have previously shown that S100A4 is expressed and secreted in TC cells and increases TC cell motility (14,15), but the signaling pathway mediating this action in TC is not fully understood. Several S100 proteins were shown to bind to RAGE in their dimeric and oligomeric forms in a calcium-dependent manner (27,39,40). S100A4 was shown to bind to RAGE in vitro (30,33) and activate RAGE in human chondrocytes of patients with osteoarthritis (36). It was demonstrated in human colon cancer cells that RAGE mediates increased cell motility in the presence of S100A4 (33). On the other hand, RAGE-independent actions of extracellular S100A4 were reported in endothelial cells (41) and in hippocampal neurons (42). Intriguingly, it was found that TC cells and the majority of follicular thyroid cells isolated from thyroid adenoma expressed the pattern recognition receptor RAGE, whereas thyroid follicular epithelial cells derived from normal thyroid tissue lacked RAGE expression. Extracellular S100A4 had a potent auto-/paracrine effect on TC cells and enhanced cell motility via activation of RAGE.
To the best of the authors' knowledge, this is the first report on the expression of the cytoplasmic RAGE-interaction partner Dia-1 in TC cells. Dia-1 was essential for extracellular S100A4 to induce increased migration via RAGE in TC. Previously, glycosylated albumin species were used as RAGE ligands to induce RAGE-mediated cell migration in the C6 rat glioma cell line through the involvement of Dia-1 (38). In TC cells, the presence of S100A4 and activation of the S100A4-RAGE-Dia-1 signaling pathway coincided with an increase in cellular active GTP-bound protein fractions of Cdc42 and RhoA and increased TC migration. Blocking Cdc42 and the Rho kinase activation resulted in markedly reduced cell migration, and exogenous S100A4 was unable to rescue the reduced TC migration. Different from TC cells, RAGE-mediated rat glioma cell migration was independent of RhoA activity (38). The inhibitor H1152P was used, which acts on Rho-associated kinase (ROCK), a downstream mediator of RhoA, and can indirectly modulate the activity status of RhoA. H1152P was demonstrated in vitro to bind to and inhibit PKA at higher concentrations (43,44). PKA can phosphorylate and increase the level of active GTP-RhoA at the leading edge of migrating cells (45). At the H1152P concentrations used here in TC cells, the reduction in GTP-bound RhoA likely resulted from PKA inhibition (Supplementary Fig. S2). This coincided with a 50% reduction of TC migration and could not be rescued by exogenous S100A4. Intriguingly, cellular RAGE levels had a profound impact on the level of GTP-bound RhoA in TC. Incomplete knockdown of RAGE in TC cells significantly reduced active GTP-bound RhoA but had little effect on GTP-Cdc42 levels in TC cells expressing S100A4. Thus, Cdc42 may be the major small GTPase targeted by the S100A4-RAGE-Dia-1 signaling pathway, whereas S100A4-mediated RhoA activation likely is indirect in TC cells.
S100A8/9 proteins have previously been shown to bind to and activate TLR4 (46). Recently, S100A4 was also shown to serve as a ligand for TLR4 in mammary adenocarcinoma cells and to activate pro-inflammatory pathways (37). Although TLR4 was expressed in TC cells, siRNA-mediated knockdown of RAGE did not affect the expression of TLR4 indicating that TLR4 likely plays no significant role in mediating the pro-migratory response of extracellular S100A4 in human TC cells.
The absence of RAGE in normal follicular thyroid cells and its presence in TC suggest that in their tumor microenvironment, TC cells can respond in an autocrine/paracrine manner to secreted S100A4. Cancer-associated fibroblasts in several cancer entities have been described to produce S100A4 (47,48), and several immune cells such as macrophages and activated lymphocytes were identified as a source for S100A4 in the vicinity of tumors (49). The authors have previously shown that S100A4 is secreted from TC cells in vitro (14,15), further emphasizing the role for S100A4 in paracrine intratumoral signaling. Similar to the previously described induction of migration of endothelial cells (50) and in colorectal cancer cells (33) by S100A4, the migration-stimulating effect on TC cells was only seen at S100A4 concentrations ≥500 nM. The current model of RAGE–ligand interaction proposes the organization of RAGE as unstable preassembled receptor clusters, which requires stabilization through ligand binding to sustain RAGE signaling (25,40,51). RAGE clustering favors binding of multimeric S100 proteins (40,51), and this, in turn, enables stronger activation of RAGE (23). In agreement with previous work, the present data suggest that high intratumoral S100A4 concentrations activate RAGE and cause Cdc42/RhoA-mediated cytoskeletal rearrangements that facilitate enhanced TC cell migration.
Ligand-induced RAGE signaling was shown to activate the p44/42 MAPK pathway in several cell types (25,30,52 –54), and a robust upregulation of the phosphorylated species of MAP kinases ERK1/2 was also observed in TC cells exposed to S100A4. Was this ERK1/2 activation contributing to S100A4-mediated TC migration? S100A8/9 were shown to elicit ERK1/2 phosphorylation via RAGE in cancer cells (27,55), and S100A4-induced cell migration involved activation of ERK1/2 in vascular smooth muscle cells (56) and in human colon cancer cells (33). However, different experimental data are presented here, strongly suggesting that S100A4-induced ERK1/2 phosphorylation was not mediated through RAGE in human TC cells. Neither RAGE silencing nor Dia-1 knockdown was able to abolish ERK activation by rhS100A4. In addition, MAPK signaling in TC cells was robustly induced at concentrations of rhS100A4 well below those required to induce TC migration. Furthermore, exogenous S100A4 was still able to increase cell migration when ERK1/2 phosphorylation was inhibited with the MEK inhibitor U0261. In light of previous research showing that extracellular S100A4 interacts with endogenous EGFR ligands such as amphiregulin, betacellulin, and EGF and induces ERK1/2 phosphorylation in murine embryonic fibroblasts (57), it is tempting to suggest that S100A4 may utilize more than one receptor in TC cells. By blocking EGFR phosphorylation, the activation of the p44/p42 MAPK pathway could be abolished (Supplementary Fig. S3). The authors are currently investigating the role of extracellular S100A4 in the activation of EGFR signaling in TC cells.
RAGE is considered a pattern-recognition receptor (58) recognizing several different ligands, among them HMGB1, which serves as mediator of inflammatory responses (23). The expression of RAGE in primary cells isolated from TC and follicular adenoma specimens contributes to the understanding of inflammatory processes within the thyroid gland as a potential mechanism to promote cancer development and progression, especially for PTC (59), and supports a role for S100A4 during the early stages of PTC development (11).
In summary, secreted S100A4 targets RAGE, which is exclusively present on human TC derived from follicular epithelial cells. Within the tumor microenvironment, S100A4-induced RAGE/Dia-1/small GTPases signaling is likely an important promoter of TC migration. New therapeutic strategies targeting this signaling pathway may prevent TC invasion and metastasis.
Footnotes
Acknowledgments
S.H.K. and T.K. are grateful for financial support from the Natural Science and Engineering Research Council of Canada (NSERC) and the Department of Surgery Research Fund (A.P., S.H.K.). We thank Mr. Ron Brereton and his team for excellent assistance with the surgical thyroid tissue specimens.
Author Disclosure Statement
There are no competing interests to disclose for any of the authors of this manuscript.