
Abstract
Background:
Iodine is a crucial component of thyroid hormones, and several reports have shown that iodine per se is implicated in the physiopathology of other organs.
Methods:
Innovative ion chromatography detection following a four-step temperature ramp microwave digestion in 25–50 mM nitric acid was developed to measure total iodine in biological fluids and tissue samples from female Sprague-Dawley rats supplemented with 0.05% molecular iodine (I2) or 0.05% potassium iodide (I−) in drinking water.
Results:
The reported method allows the measurement of total iodine with a limit of quantification of 13.7 μg L−1, recoveries of 96.3–100.3%, and intra- and inter-assay variations, of 3.5% and 7.4% respectively. Analysis of biological fluids showed that after 48 hours, iodine-supplemented animals exhibited significantly higher levels of total iodine in both serum and urine compared with those supplemented with iodide. The half-life of iodine in serum and urine measured over the first 48 h showed similar patterns for both the I2 (7.89 and 7.76 hours) and I− (8.27 and 8.90 hours) supplements. Differential uptake patterns were observed in tissues after 6 days of supplements, with I− preferentially retained by thyroid, lactating mammary gland, and milk, and a slightly but significantly higher capture of I2 in pituitary, ovary, and virgin mammary gland.
Conclusions:
We developed a rapid, selective, and accurate digestion method to process fluid and tissue samples that permits reproducible measurements of total iodine by ion chromatography; iodine or iodide supplement show a similar serum and urine half-life, but organ-specific uptake depends on the chemical form of the iodine supplement.
Introduction
I
At present, the iodine content in biological samples is determined almost exclusively by one of two methods: one is a simple kinetic spectrophotometric method based on the Sandell-Kolthoff reaction, in which iodide catalyzes the reduction of ceric ammonium sulfate (yellow) to the colorless cerous form in the presence of arsenious acid. This reaction is catalyzed by trace amounts of iodide, but many organic and inorganic substances can potentially interfere, affecting the reaction rate. Complete mineralization of the sample to digest the organics is thus a necessity. This digestion step typically involves perchloric acid, which requires special hoods and precautions to prevent explosion. The other method used is at the opposite end of sophistication: inductively coupled mass spectrometry provides superb sensitivity and allows direct analysis of diluted biological fluids (urine or serum). Nevertheless, for tissue samples, predigestion is still necessary (12). We developed a rapid and simple method to determine total iodide by ion chromatography after microwave-assisted digestion with 25–50 mM nitric acid (HNO3) of biological fluids and several tissues from female rats supplemented with 0.05% molecular iodine (I2) or 0.05% potassium iodide (I−) in drinking water. This method allows rapid, selective, and accurate analysis of fluid and tissue samples, permitting reproducible determinations of total iodine content. The results obtained demonstrate a similar half-life for iodine in serum and urine but differential tissue uptake, depending on the chemical form of the iodine supplement.
Materials and Methods
Reagents
All solutions were prepared with deionized water (NANOpure Infinity Unit; Barnstead Thermolyne, Dubuque, IA), and all reagents were high-performance liquid chromatography (HPLC) grade. Nitric acid and iodine sublimate (molecular iodine; I2) were purchased from J.T. Baker (Phillipsburg, NJ, and Edo. de México, México respectively). Potassium iodide was obtained from Sigma-Aldrich (St. Louis, MO), ketamine and xylazine from Cheminova (Mexico City, México), and oxytocin from Novartis (Sao Paulo, Brazil).
Digestion and iodine quantification
Fluids and tissues were digested in a MARSXpress microwave system (CEM Corporation, Matthews, NC) programmed for a four-step temperature ramp from 60°C to 200°C over 85 minutes as summarized in Table 1. Total iodine was quantified in a Dionex ICS-2500 HPLC/IC chromatograph (Sunnyvale, CA) with the configuration and parameters summarized in Table 2. A stock solution of potassium iodide (100 mg/L) was prepared in deionized water. Working standards were prepared by further diluting the stock solution with deionized water to the required concentration.
HNO3, nitric acid.
Animals and protocols
Virgin or lactating (day 4 postpartum, 8 pups) female Sprague-Dawley rats were born in a colony maintained at the Instituto de Neurobiología, Universidad Nacional Autónoma de México (UNAM, Querétaro, México). Animals were kept under regulated temperature conditions (22±1°C) at 50% humidity on a 12:12 hour light–dark cycle and permitted ad libitum access to food (Purina Certified Rodent Chow) and water. Animals were housed and handled in accordance with UNAM Animal Use Committee Guidelines. To analyze the half-life of I2 or I− in serum or urine, groups of virgin animals were supplemented with one of these chemical forms of iodine in drinking water for 48 hours; after this period, the iodine source was removed, and animals in groups of six were sacrificed immediately (time 0) and after 6, 12, 24, and 48 hours. To analyze the uptake and metabolism of each chemical form of iodine in different fluids or tissues, virgin and lactating animals were assigned to the following groups: Control (deionized water); iodine (0.05% I2), and iodide (0.05% KI), and sacrificed on day 6. The thyroid status and general health were monitored by measuring serum triiodothyronine (T3) by the homologous radioimmunoassay method (11), serum thyrotropin by the Biotrack assay system (rat radioimmunoassay method, Amersham Co., New Jersey), and body weight (reported as % change between day 0 and day 6).
Biological samples
After each treatment, the animals were anesthetized with a mixture of ketamine/xylazine (30 mg/6 mg per kg body weight), the milk was removed by administering oxytocin (30 mU; intraperitoneal), and the animals were sacrificed by decapitation. Fluids (urine, serum, and milk) and tissue (thyroid, mammary gland, pituitary, and ovary) samples were frozen in dry ice and kept at −70°C. Urine and serum (100 μL each) were analyzed both directly after centrifugation (1300g, 15 minutes) and after microwave-assisted digestion in 3 mL of 25 mM HNO3. Tissues were digested in 5 mL 50 mM HNO3 in a microwave system. For the pituitary assay, each pool contained four to five glands, and four pools from each group were analyzed. Thyroid glands were processed individually, and 50 mg samples of all other tissues were digested.
Statistical analysis
All results are expressed as the mean±standard deviation. Data were analyzed using a one-way analysis of variance. Differences between means were tested by Tukey's multiple comparisons test. Values with p<0.05 were considered statistically significant. The regression equation, correlation coefficient, and half-life (exponential decay equation) were analyzed using the statistics software package Graph Pad Prism 6.
Results
Method validation
Selectivity
Selectivity was validated through the repeated measurement of blanks (samples without iodine), standards of potassium iodide in concentrations between 12.5 and 300 μg/L, and samples of fluids or tissues. As shown in Table 3 and Figure 1, standards and samples exhibited one distinct, well-resolved peak at average retention times of 3.50 minutes, with a relative standard deviation of 1.6%. This peak, which is absent in the chromatogram of the control blank (Fig. 1), corresponds to iodide. Chromatograms of samples also show that nonretained solutes elute well before iodide (peak at ca. 1.5 minutes). Slight variations in the retention time of iodide among the different sample types are within the error of standard measurements (Table 3) and do not affect selectivity, given the good resolution of the peak.
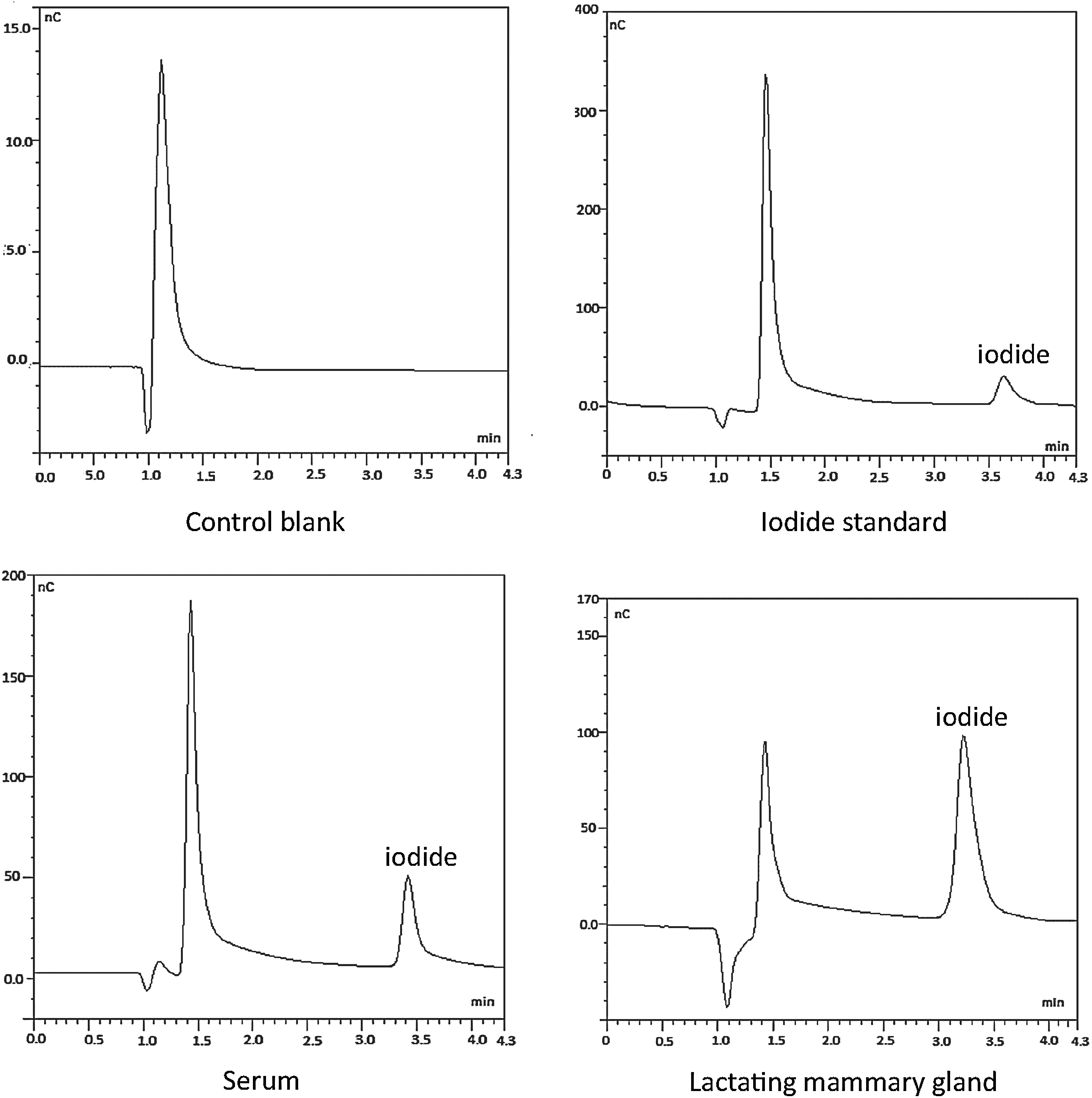
Chromatograms of iodide in a water sample (control blank), an iodide standard (100 μg/L), and digested samples of serum and lactating mammary gland. Measurement conditions are summarized in Table 2 and described in the Materials and Methods section. For standard and samples, iodide elutes at retention times of around 3.50 minutes; this peak is absent in the control blank. Note the different scales on the y-axis.
Linearity and sensitivity
Seven iodine calibration standards with concentrations ranging from 12.5 to 250 μg/L were measured in six different assays. Peak areas were plotted against the concentration of the calibration standards, and the calibration curve was fitted by linear least-squares regression (Fig. 2). The average slope and y-intercept of the regression equation are 0.0127 and −0.0536 respectively, and the correlation coefficient (r) was 0.9982. A limit of quantification of 13.7 μg/L was estimated for the method; this concentration corresponds to 10 times the standard deviation of repeated blank measurements (Table 4).

Average calibration curve over the range from 0 to 250 μg/mL obtained from six independent assays. The regression equation and the correlation coefficient (r) are also included.
The table includes parameters of the regression equation and the estimated limit of quantification of the method.
b, y-intercept; LOQ, limit of quantification; m, slope; R 2, coefficient of determination; SD, standard deviation.
Recovery
Recovery experiments were carried out in order to assess the accuracy of the method. This parameter was determined by measuring three different samples each of serum from the control group, serum from an I2-supplemented group, and mammary gland before and after the addition of a known iodide concentration. Recoveries of 96.3–100.3% demonstrate the accuracy of the method, especially for serum samples, with recoveries above 99.7% (Table 5).
Precision
The intra-assay variation was tested by performing five replicate analyses of two serum samples from the control group (analyzed direct and after digestion) and one serum sample from the I2-supplemented group (measured directly). For the inter-assay variation, one digested urine sample from an I−-supplemented rat was measured in eight independent assays. The relative standard deviation was 3.5% for intra-assay and 7.4% for inter-assay variation. The absolute standard deviation, variance, and relative standard deviation are summarized in Table 6.
Intra-assay variation was evaluated on the basis of five measurements of two serum samples from the control group (analyzed directly and after microwave-assisted digestion) and one serum sample from the I2-supplemented group (measured directly). For the inter-assay variation, one digested urine sample from an I−-supplemented rat was measured in eight independent assays.
S, absolute standard deviation; RSD, relative standard deviation; S2, variance.
Specificity
To analyze the specificity for iodine components, serum and urine samples from control and I2- or I−-treated animals were quantified directly or after HNO3 digestion. Figure 3 shows that the iodine concentrations obtained for digested serum samples are higher than those obtained from direct measurements. The largest relative increases were found in serum samples from the control group, which yielded iodine contents of 10.6 μg/L by direct analysis and 429 μg/L by microwave digestion (Table 6). Differences were also observed in serum samples from iodine- or iodide-supplemented rats, with increases after digestion of ca. 80–100% (Fig. 3). We interpret the difference in iodine content in digested serum samples as the iodine released from the organic fraction, which would include thyroid hormones. Such dramatic differences before and after digestion were not observed for urine samples, indicating that iodine is present as free iodide in urine, as previously reported (12).
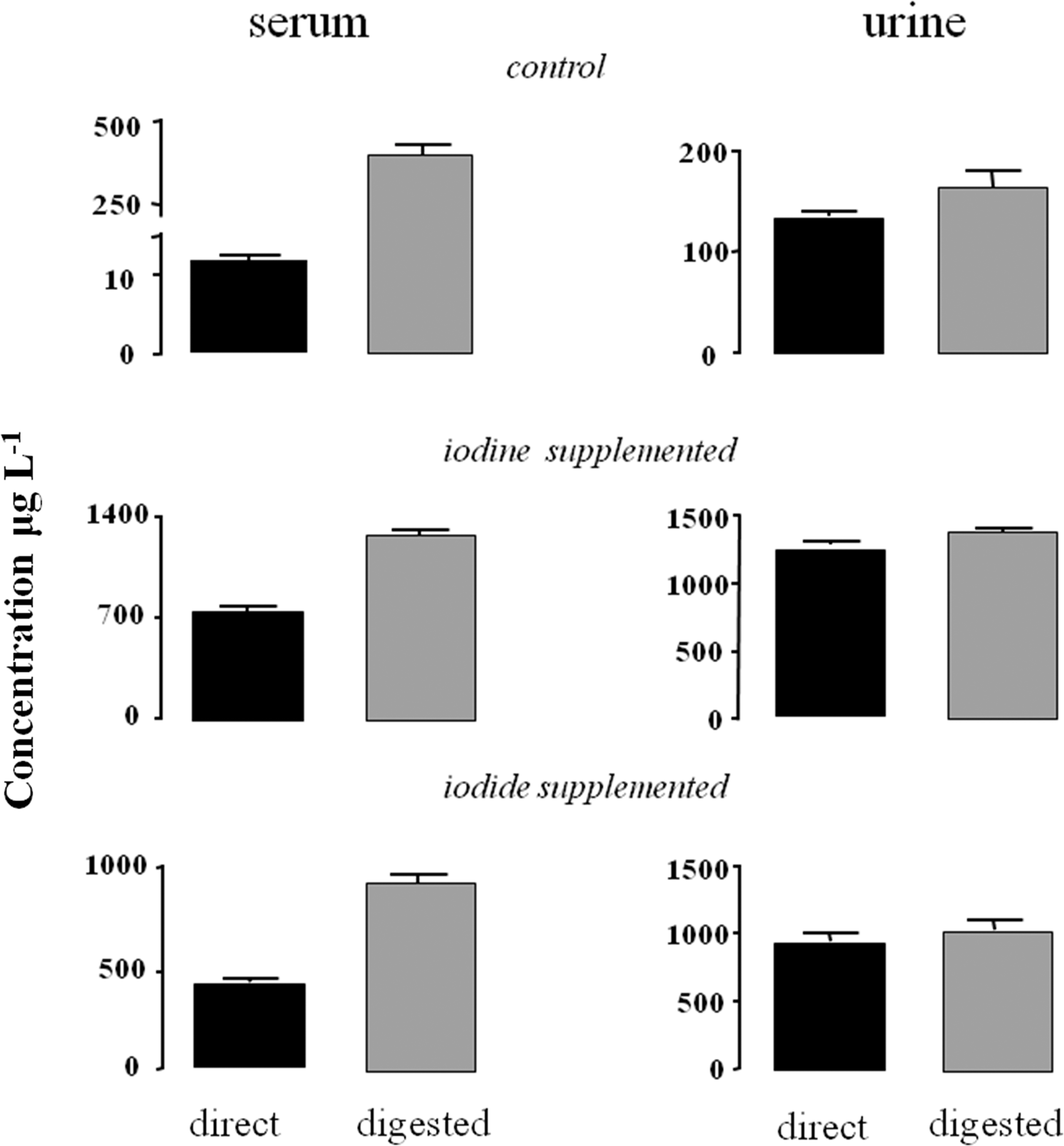
Concentrations (μg/L) of free iodide (direct) or total iodine (digested) contained in control and I2- or I−-treated animals.
Iodine versus iodide metabolism
The half-life of free iodide was obtained by analyzing serum and urine samples obtained from virgin animals after a 48 hour treatment with either I2 or I−. Iodide decay displays similar patterns in serum and urine samples of both groups (Fig. 4), which translates into similar half-lives of free iodide in serum and urine of animals treated with iodine (7.89 and 7.76 hours respectively) or iodide (8.27 and 8.90 hours respectively). These results indicate that iodine ingestion and intestinal absorbance were similar for both chemical forms of iodine. In animals supplemented for 6 days, similar patterns for free iodide were found in serum and urine from control, I2-treated, and I−-treated groups (Fig. 5). In both sample types, the highest free iodide concentrations were in I2-treated animals. The lowest values were found in control animals given a standard diet that contains 0.8 mg/kg iodide.
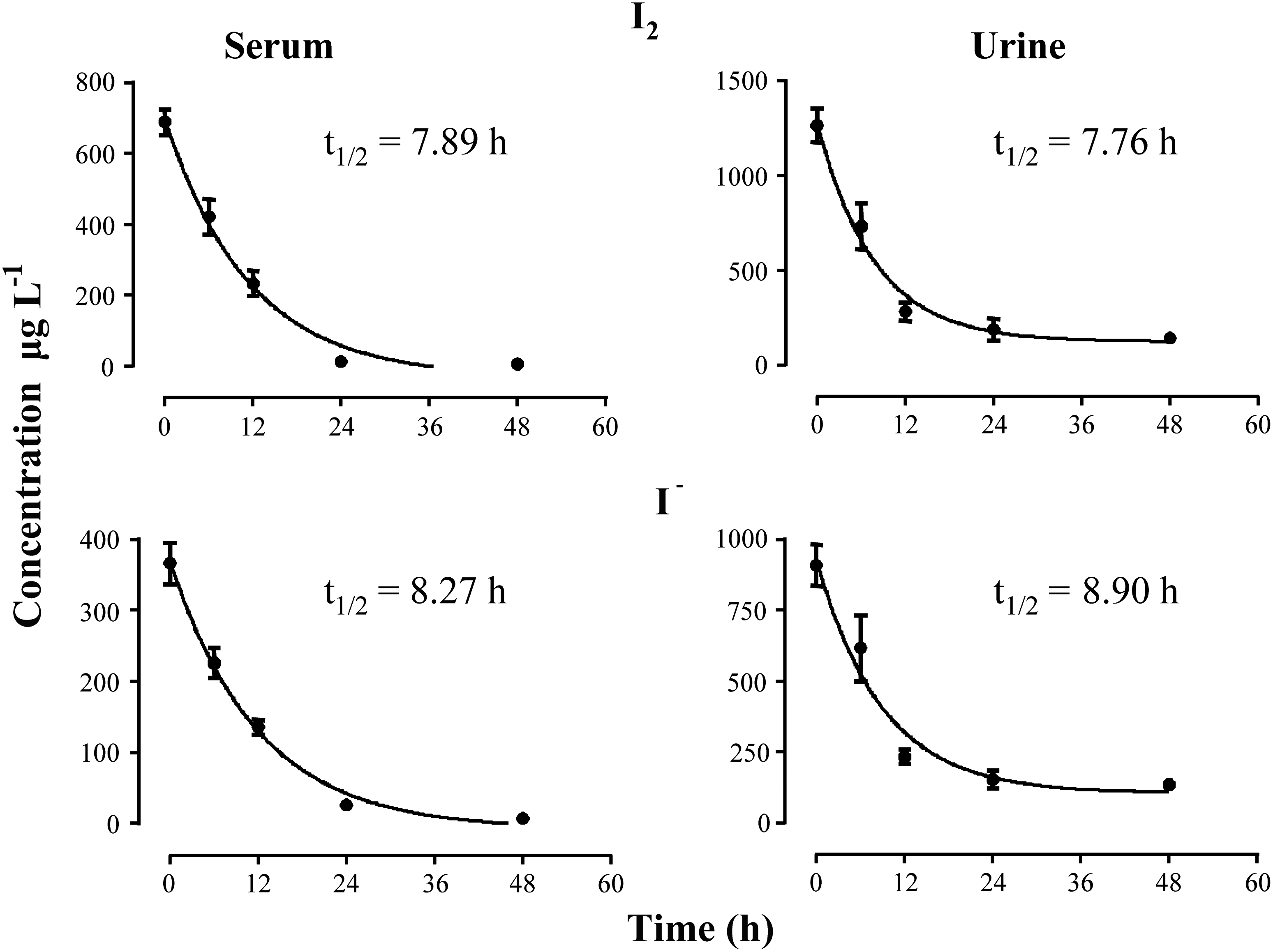
Free iodide content in serum and urine at different times after 48 hour treatment with I2 or I− supplementation. Half-life (t 1/2, hours) was determined using the software package Graph Pad Prism 6.
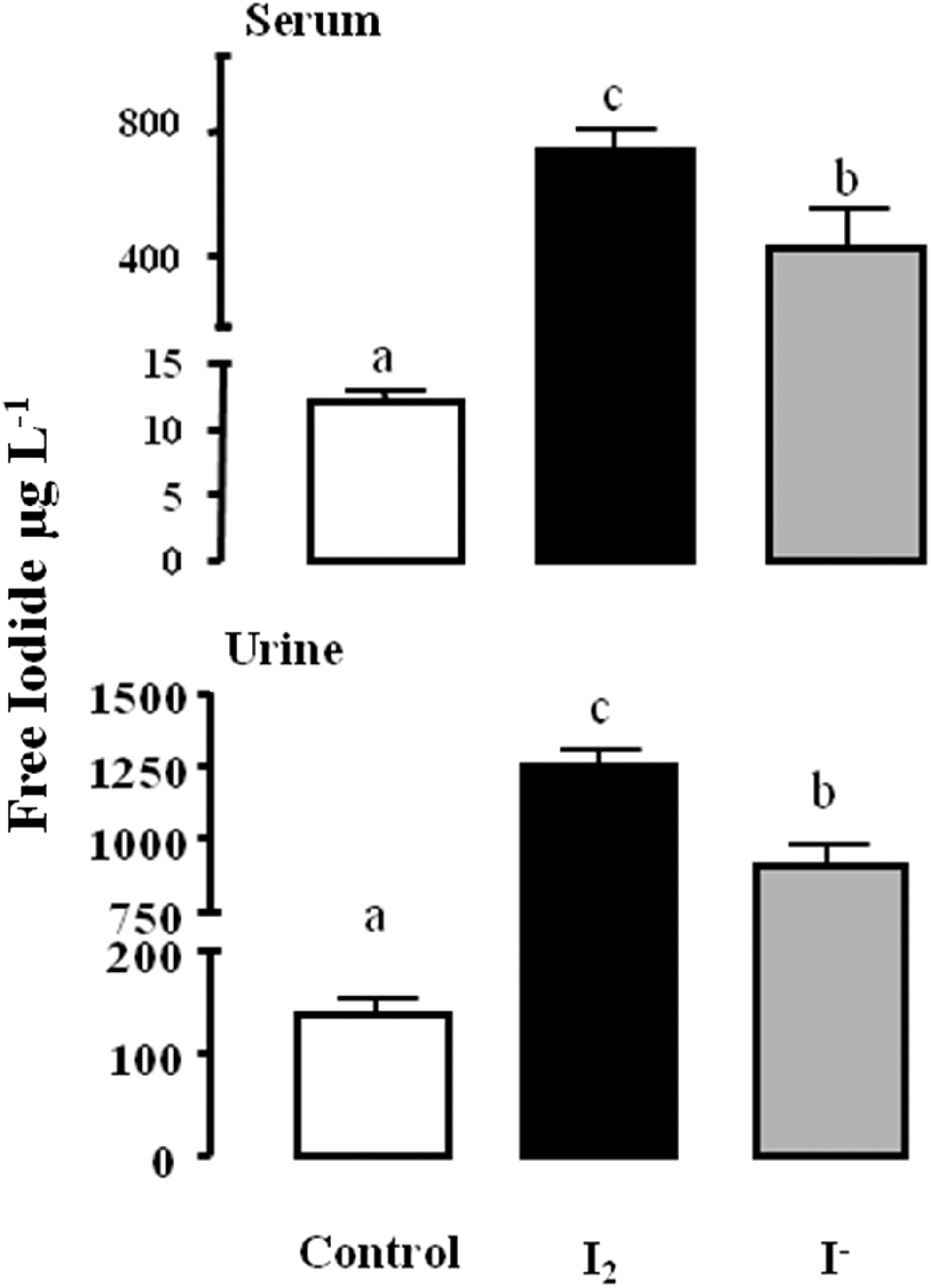
Concentration of free iodide in serum and urine of control and treated animals after 6 days of I2 or I− supplementation. Data are expressed as the mean±standard deviation (SD) (n=6). One-way analysis of variance (ANOVA) and Tukey's multiple comparisons test were used to determine statistical significance. Means with different letters are significantly different (p<0.05).
Analysis of total iodine content in tissues and milk after 6 days of I2 or I− supplement shows that I− is preferentially taken up and retained by thyroid, lactating mammary gland, and milk (Fig. 6). In contrast, pituitary, ovary, and virgin mammary gland take up both types of iodine, with a small but significant preference for I2. Figure 7 shows that the iodine supplement, in either chemical form, did not modify the thyroid status (serum T3 and thyrotropin levels) or the general health (body weight) of the animals after 6 days of treatment.
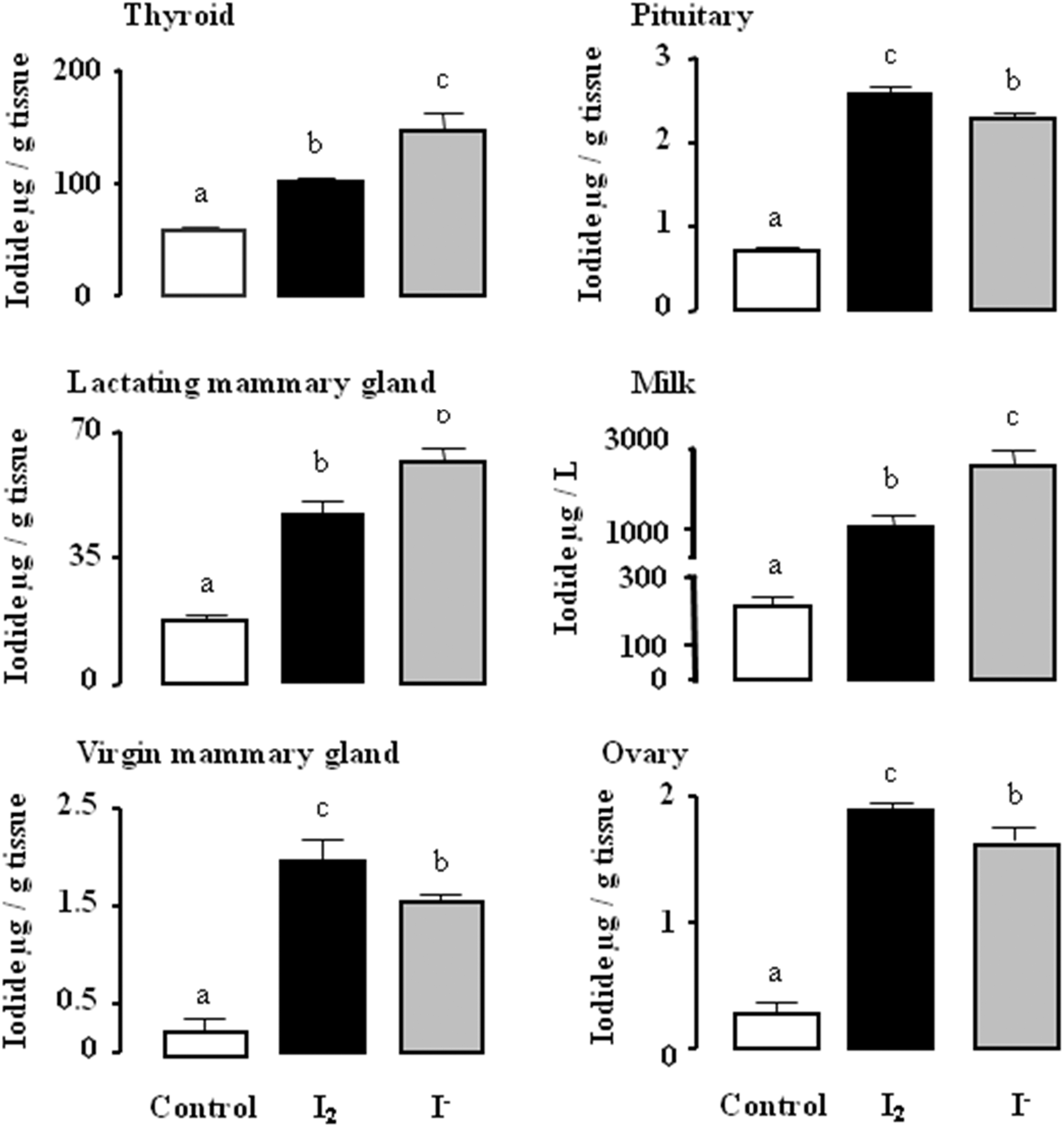
Total iodine content of tissues and milk after six days of I2 or I− supplementation. Data correspond to digested samples and are expressed as the mean±SD (n=6). One-way ANOVA and Tukey's multiple comparisons test were used to determine statistical significance. Means with different letters are significantly different (p<0.05).

Thyroid status: body weight and serum levels of triiodothyronine and thyrotropin do not change in control or treated animals after 6 days of I2 or I− supplementation. Data are expressed as the mean±SD (n=6).
Discussion
In this study, a simple, rapid, and highly sensitive method is proposed for the determination of total iodine in biological samples. This ion chromatography method does not require any separation or preconcentration steps and was applied directly to determine free iodide in serum or urine, and after safe and reproducible microwave-assisted acid digestion, it permits total iodine to be measured in tissue samples. The direct quantification of free iodide in urine or serum is similar in sensitivity to other methods, ensuring the accurate measurements of iodine above 13.7 μg/L (13). Urinary iodide is the epidemiological indicator used to monitor the iodine nutrition status in a population, and the median levels are used to categorize a population as severely deficient (<20 μg/L), moderately deficient (20–49 μg/L), mildly deficient (50–99 μg/L), adequate (100–199 μg/L), above requirements (200–299 μg/), and excessive intake (>300 μg/L) (1). Our method is applicable for all these ranges with an excellent selectivity, accuracy, and precision. For tissue samples, our innovative digestion method using nitric acid and a temperature ramp protocol results in the total organic digestion in 85 minutes, as well as the complete conversion of iodine to hypoiodous acid, 96–100% of which can be recovered due to its high solubility in water, and there is no need to include specific solutions to absorb gaseous iodine products like hydrogen iodide or I2 released during combustion; it also eliminates the use of toxic or dangerous reagents like perchloric acid (14). Moreover, the differences in values obtained with this method in serum or urine samples before and after digestion agree with previous reports indicating that the iodine bound to thyroid hormones is 300–400 μg/L in serum and 20–40 μg/L in urine (15,16).
The second objective of this work was to analyze the iodine content of several fluids and tissues from normal and lactating rats given 0.05% I2 or I− supplements for 6 days. It is well established that moderately high amounts (3–10 times the safe upper limit of 150–200 μg/day) of iodine exert, in euthyroid subjects, a transient decrease in the synthesis of TH within the first 24–48 hours. This inhibition of TH synthesis is called “the acute Wolff–haikoff effect,” and it is due to increased intrathyroidal iodine concentrations, which inhibit the iodination of tyrosyl residues of thyroglobulin by thyroperoxidase (17,18). After 48 hours of I− excess, the thyroid gland “escapes” from this effect by an adaptation that decreases the thyroid iodine trap, thereby decreasing the intrathyroid iodine concentration. The chronic I− excess inhibits the mRNA and protein expression of the sodium iodide symporter and thyroperoxidase, among others, and after this initial 48 hours, the synthesis of TH is reestablished (19). The Wolff–Chaikoff effect is abolished by inhibitors of I− organification, such as methimazole and propylthiouracil; therefore, the existence of a putative organic iodine compound that mediates this effect has been proposed (20). In previous reports, our group described that iodine and iodide supplements exert differential responses in the thyroid and normal or cancerous mammary glands. In the thyroid gland, I− but not I2 supplement induced the “Wolff–Chaikoff effect,” and neither of these iodine supplements modified sodium iodide symporter or lactoperoxidase expression in normal or tumoral mammary glands (11).
In the present work, we report the first comparison of these two chemical forms of iodine with respect to half-life, and organ-specific uptake after the “escape” from the Wolff–Chaikoff effect (6 days). Our results show that I2-supplemented animals exhibit higher total iodine concentrations in serum and urine in comparison with I−-treated animals. This result is a consequence of the fact that a 0.05% I2 solution contains more iodine atoms than a 0.05% KI solution. Nevertheless, over an interval of 48 hours, the half-life in serum and urine were practically equal for animals supplemented with either form of iodine, indicating first, that I2 and I− exhibit similar uptake and kinetic parameters and second, that after 48 hours of supplement, the serum compartment is saturated, and the excess iodine is excreted in the urine. This observation agrees with the measurement made after 6 days, when the median values of total iodine in serum remain equal in both the I2- and I−-treated animals.
Our results show that the thyroid, the lactating mammary gland, and milk concentrate significantly more I− than I2, whereas the pituitary, the virgin mammary gland, and the ovary exhibit higher concentrations of I2. Although the preference of the latter organs for I2 seems small, when organ size is considered, their total amount of I2 uptake is significant. In addition, it is well established that, with exception of the thyroid and lactating mammary gland, other tissues that take up iodine do not have the enzymatic system to oxidize and organify I−, resulting in its transitory uptake and release within minutes or hours (21 –23). By contrast, when I2 is captured, it is rapidly incorporated into lipids and other components within the cells (24). Some authors have proposed an interchange between I2 and I−. Molecular iodine can be reduced to I− by reacting with contents of the gastrointestinal tract and the high peroxidase activity in the small intestine could oxidize I− to I2 and increase its reactivity (25). However, these reactions could be minimal because first, Thrall et al. (9) and we (24) failed to detect radioactivity associated with lipids (by thin-layer chromatography) from animals or cells supplemented with 125I−, which was clearly observed in those supplemented with 125I2, and second, several pieces of data in the literature show that I− treatment does not exhibit the antiproliferative and apoptotic effects of I2 treatment in in vivo and in vitro cancer models (4,5,24,26). Our results corroborate previous data showing a differential distribution of iodine in organs known to take up iodine, such as the thyroid, mammary gland, and ovary (21 –23), and we also demonstrate iodine uptake in other organs like the pituitary, where it has never been reported before.
The physiological significance of extrathyroidal iodine uptake has been partially analyzed. In the lactating mammary gland, it is considered as a physiological adaptation to ensure an adequate iodide supply to the neonate through the milk (21,23). It has been demonstrated that iodine helps to maintain the normal integrity of the non-lactating mammary gland. Eskin et al. (7) showed that iodine deficiency alters the structure and function of mammary gland from virgin rats, and that I2 is effective in diminishing ductal hyperplasia and perilobular fibrosis secondary to this iodine deficiency. Similarly, I2 treatment of patients with benign breast disease is accompanied by a significant bilateral reduction in breast size, in addition to causing a remission of disease symptoms, which is not observed when I− or protein-bound iodide is administered (4,27).
In prostate gland, another tissue with iodine-uptake capacity, an iodine supplement prevented steroid-stimulated prostatic hyperplasia in male rats (8) and significantly improved symptoms, prostatic specific antigen levels, and urine flow in men with benign prostatic hyperplasia (28). Moreover, other authors have proposed that iodine is an antioxidant used since ancestral times, and showed that in different organs including the brain, it prevents lipoperoxidation (2,29). Finally, as we mentioned above, a significant body of data demonstrates that I2 exhibits antiproliferative and apoptotic effects in cancer models (2,18,25,30). All these data emphasize the importance of analyzing the physiological extrathyroidal effects of iodine.
In summary, we developed a rapid and simple ion chromatography method to determine free iodide or total iodine in biological fluids and tissues, and we demonstrated that iodine is distributed differentially among rat tissues depending on the chemical form administered.
Footnotes
Acknowledgments
The authors are grateful to Juana Cárdenas, Brenda Ugalde, Juan Gabriel Cano Ortega, Eliu Ayala de la Portilla, Martín García Servín, and Alejandra Castilla for technical assistance, Francisco Javier Valles and Rafael Silva for bibliographic assistance; Leonor Casanova and Lourdes Lara for academic support; Alberto Lara, Omar González, Ramón Martínez, and Sandra Hernández for computer assistance; and Dr. Dorothy Pless for proofreading. The work of our group was partially supported by grants UNAM/DGAPA-PAPIIT, IN200813, IN202513, and CONACYT 127368, and 176911.
Author Disclosure Statement
No competing financial interests exist.