
Abstract
Background:
Autoimmune thyroiditis (AIT) is a very common endocrine disorder in pregnancy. However, the effect of maternal positive thyroglobulin antibodies (TgAb) on brain development of offspring remains unclear. This study used an experimental autoimmune thyroiditis model in CBA/J mice and determined whether isolated positive maternal TgAb directly affected learning and memory abilities of offspring.
Methods:
An experimental autoimmune thyroiditis model was established in CBA/J mice through immunization with murine thyroglobulin (mTg). Measuring thyroid function and serum TgAb titer confirmed the presence of isolated positive maternal TgAb. Offspring serum TgAb titer, MCT8, Reelin, RC3, and BNDF mRNA expression in the brain, and brain histology were measured on postnatal days 0, 10, and 40 (PND0, PND10, PND40), and nerve cell migration (BrdU labeling) at PND40. Morris water maze, long-term potentiation (LTP), and LTP-related factor ERK1/2 levels were measured at PND40 to determine offspring spatial learning and memory development.
Results:
Maternal serum TgAb titers increased and remained elevated through pregnancy compared to controls. Thyrotropin and thyroid hormone levels were normal. The T group offspring (Tg immunized) had higher TgAb titers than the control (C) group. However, antibody titers time-dependently decreased. MCT8, Reelin, RC3, and BDNF mRNA expression in the whole brain were similar in the T and C groups on PND0, PND10, and PND40. Neuronal distribution and BrdU from the cerebral cortex and hippocampus were similar in the T and C group offspring. Morris water maze tests, excitatory postsynaptic field potentials, and ERK1/2 levels were also similar between the T and C groups.
Conclusions:
Isolated positive maternal TgAb did not clearly influence the learning ability and memory of offspring, or nerve cell migration, despite a transient increase in TgAb in immunized mice.
Introduction
A
An experimental autoimmune thyroiditis (EAT) model by immunization with thyroglobulin (Tg) is a classic method to induce AIT in mice, and is characterized by thyroid lymphocytic infiltration and high levels of serum thyroid antibodies (5).
The epidemiological data remain controversial regarding whether positive thyroid autoantibodies in pregnant women could affect the cognitive development of offspring (6,7). The present study was initiated to determine whether isolated positive maternal TgAb could exert direct effects on brain development in offspring, especially in terms of learning and memory, in an EAT model.
Materials and Methods
Animal studies
Sixty four-week-old CBA/J female mice (Beijing HFK Bioscience Co.) were divided into two groups: test group (T group; n=30) and control group (C group; n=30). All animal procedures were performed in accordance with the protocols approved by the Institutional Animal Care and Use Committees of the China Medical University.
Preparation of murine thyroglobulin
Murine Tg (mTg) was prepared from frozen mouse thyroids (KM mouse), as described by Imaizumi et al. (8).
Immunization and experimental procedures
Subcutaneous injection of 200 μg mTg with complete Freund's adjuvant (Sigma, F5881) was used to immunize T group mice, and the boosting injection was performed two weeks later with the same dose of mTg in incomplete Freund's adjuvant (Sigma, F5506). The same dose of phosphate buffered saline (PBS) instead of mTg was used to immunize C group mice. Mice (T group: n=7; C group: n=7) were euthanized within four weeks of the last immunization, marking the M1 time point. The remaining mice were mated with CBA/J male mice (aged 8–10 weeks). Mice with vaginal plugs were counted as originally pregnant. Female mice (T group: n=5; C group: n=5) were euthanized at the end of the pregnancy, marking the M2 time point. Pups were euthanized on postnatal days 0, 10, and 40 (PND0, PND10, and PND40), respectively.
Thyroiditis score
Thyroids were removed and fixed in 10% formalin in PBS and stained by hematoxylin and eosin assay. The thyroid grading was performed according to the thyroiditis classification standard published by Imaizumi et al. (8). Percentages of thyroiditis were calculated as percent of mice with thyroiditis, and the mean thyroiditis grades were calculated as the total thyroiditis grades divided by the total number of mice.
Enzyme-linked immunosorbent assay–detected mTg antibody
Murine sera were analyzed using 96-well plates, which were coated overnight with 1 mg/mL of purified mTg in carbonate–bicarbonate buffer (pH 9.6). After washing and blocking with 1% bovine serum albumin (BSA)/PBS, wells containing individual murine sera were incubated (maternal dilution=1:500; pup dilution=1:100). Bound antibodies were detected using alkaline phosphatase-labeled rabbit anti-mouse IgG (Sigma, A9044, 1:40,000 dilution), and finally TMB (Sigma, T0440) was added. At the appropriate time, 2 M HCl was added. Absorbance at the 450 nm filter (TECAN, F200PRO) was measured after termination.
Thyroid function tests
Free thyroxine (fT4) and thyrotropin (TSH) were measured by enzyme-linked immunosorbent assay (ELISA) kit (USCNLIFE, CEA185Ge, and CEA463Mu). All samples were measured twice, and specific experimental steps were performed in accordance with the kit specifications.
Morris water maze
The Morris water maze consists of a white circular swimming pool divided into four quadrants of equal area: I, II, III, and IV. The pool was filled with milk diluted with water (20±2°C). A circular platform (diameter 10 cm) was located 1 cm below the water surface in the middle of the II quadrant. Pups at PND40 (T group: n=8; C group: n=10) swam to accommodate the water maze and platform on the first day. Training began from the second day and continued until the fifth day, with each pup undergoing four trials each day. In each trial, pups were placed into the water and allowed to search for the platform with a time limit of 60 sec, and the time spent by the pups finding the platform was recorded and assigned as escape latency time. After staying on the platform for 10 sec, pups were gently picked up, returned to the cage, and dried under a heat lamp. On the sixth day, the platform was removed, the pups were put into the water, and the time taken by each pups to reach the place where the original platform was located within 60 sec was recorded (probe trial test). Each pup swam twice.
Long-term potentiation induction in vivo
The excitatory postsynaptic field potentials (fEPSPs) were recorded as the CA3 area of the hippocampus of anesthetized (urethane 1.2 g/kg, administered intraperitoneally [i.p.]) pups. The pups at PND40 (C group: n=10; T group: n=8) were placed in a stereotaxic apparatus (SR-5M, Narishige Inc.), and holes were drilled in appropriate regions of the skull. To record the field potential response, a concentric bipolar stimulating electrode was placed in the CA1 area of the left hippocampus (AP, 1.82; L, 0.5; D, 1.75), and a glass capillary recording electrode, filled with 1% Fast Green in 3 M KCl, was placed in the CA3 of the left hippocampus (AP, 1.7; L, 2.0; D, 2.0). The position of the electrode referred to the mouse brain in stereotaxic coordinates (9). Stimulus intensity (Nihon Kohden, SEN-3201) was set at 30% of the maximum response by construction of input/output curves to evoke test responses every 30 sec at 0.04 Hz throughout the experiment. Long-term potentiation (LTP) was evoked by multiple high-frequency stimulation (MHFS), which consisted of three trains given at 2.5 min intervals. Each train consisted of 10 pulses (400 Hz) repeated every 10 sec for a period of 30 sec. The fEPSP slope was measured from the early rising phase of the recorded traces using a microelectrode recording signal amplifier (MP150, BIOPAC SYSTERMA). Recordings were normalized to the average baseline values of 20 min. LTP was induced by MHFS and measured as an increase in fEPSP slope, which was expressed as a percentage baseline of the fEPSP (fEPSP %) slope after MHFS in the two groups. Based on the pooled data, the means of the slope of fEPSP % were expressed as a percent of the corresponding pre-stimulation control.
BrdU immunohistochemistry
Mice at 13–15 days of pregnancy were injected i.p. with BrdU (Sigma, B5002) at a rate of 50 μg/g body weight. A series of 40 μm-thick brain sections of pups at PND40 were cut using a freezing microtome (Leica, CM1900) and fixed with acetone. The experimental process was in accordance with anti-BrdU antibody specification (1:500 dilution, Sigma, B8434). To perform a quantitative analysis of the cell number, Image-Pro-plus 6.0 software was used, and data were expressed as the number and location of BrdU labeling cells. Percent of positive cells=average IOD of positive cells/average IOD of all cells in the field of view (10).
Nissl staining
Whole brain specimens (five per group) were immediately fixed in 4% paraformaldehyde for 24 h, routinely dehydrated, embedded in paraffin, and sliced into 5 μm sections (Leica, RM2135). The coronal sections were stained with 1% toluidine blue. The light microscope (Olympus, BX51) was used to measure the thickness of the cerebral cortex and CA1 pyramidal neurons of hippocampus, and to observe the distribution of the neurons from the cerebral cortex and hippocampus at PND0, PND10, and PND40.
Western blot
The whole brain protein extraction from pups at PND40 was made according to the Total Protein Extraction Kit (KeyGen Biotech, KGP210). The supernatant was collected and the protein concentration measured by the BCA Protein Assay Reagent Kit. REK1/2 Western blots were performed as described by Zhang (11).
Real-time PCR
Total RNA of whole brain tissue from pups at PND0, PND10, and PND40 was extracted with TRIzol (Life Tech) according to a standard procedure, and Nano Drop2000C (Thermo Scientific) was used to determine RNA concentration and purity. Total RNA (1 μg) was reversely transcribed with random primers using the Reverse Transcription Reagent Kit (Promega) following the manufacturer's protocol. All amplification reactions were performed in triplicate on the ABI PRISM 7500 sequence detection system (Applied Biosystems). For quantitative real-time PCR, 25 ng of cDNA was used, and single transcript levels of genes were detected with a QuantiTect SyBR green master mix (TaKaRa). Primers used to detect synaptic transcripts were as follows:
β-actin:
5′-GGT CAT CAC TAT TGG CAA CG-3′(forward),
5′-TCC ATA CCC AAG AAG GAA GG-3′(reverse);
Reelin:
5′-GGA CGT ATC GGC AGT GAG AAC A-3′(forward),
5′-CAT CTC GTG AAG CAA GGT CCA A-3′(reverse);
BDNF:
5′-ATC GGT TCA CAG GAG ACA T-3′(forward),
5′-TCA GGT CAA CAT AAA CCA CCA-3′(reverse);
MCT8:
5′-CTT CTA CTT TGC GGG TGT CC-3′(forward),
5′-CCT TGC TGG AAT CTC TCT GC-3′(reverse);
RC3:
5′-AGG AGA GAG GCT GGT TGC-3′(forward),
5′-TAT CGT CGT CTG GCT TGG AG-3′(reverse).
Data were analyzed using the comparative CT method.
Data analysis
Data are presented as mean±standard error (SE). The t-test was applied to assess the statistical significance of the differences between paired groups of data, and the differences among multiple groups were assessed by one-way analysis of variance (ANOVA). The Morris water maze escape latency data were also assessed by repeated-measures ANOVA. SPSS 20.0 (IBM Corp.) was used for all data analyses. Differences were considered statistically significant at p<0.05.
Results
Building a murine model of isolated positive maternal TgAb
Thyroid histology and the thyroiditis score showed that mice in the C group had uniform follicular thyroids with even distribution and absence of atrophy, and no lymphocytic infiltration was found in either maternal thyroids (M1 and M2). On the other hand, mice in the T group manifested a disorderly arrangement of the thyroid follicle. Lymphocytic infiltration was found more or less, and the structure of a portion of follicular material was destroyed. In the T group, the prevalence of thyroiditis was 85.70% at M1 and 90.90% at the M2; the mean thyroiditis score was 2.29 at M1, and 1.25 at M2. The degree of thyroiditis decreased as time progressed. Additionally, a serum TgAb ELISA test for the T group showed that TgAb of the mice at M1 and M2 was enhanced and significantly higher than those in the C group, respectively (p<0.01). No statistical difference in TgAb OD value was found when comparing M1 with M2 in the T group (p>0.05), indicating that the serum TgAb titer was on the higher side during the whole pregnancy (Fig. 1, Table 1). Serum fT4 and TSH in maternal mice, as seen in Table 1, showed no significant differences between the two groups (p>0.05).
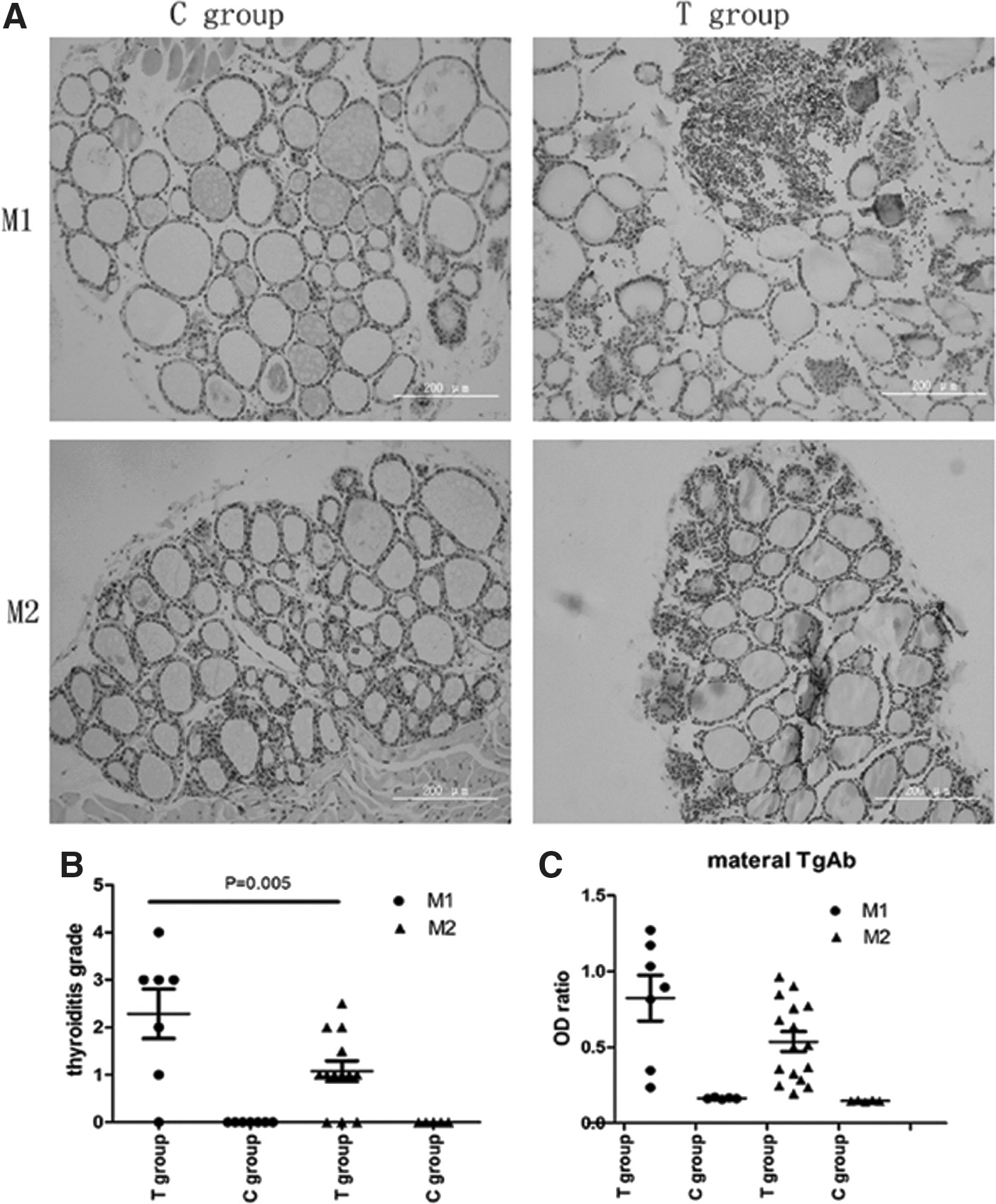
(
Values are expressed as mean±SD.
0.001, T group vs. C group in M1.
0.02, T group vs. C group in M2.
TgAb, thyroglobulin antibodies; M1, female mice at four weeks after the last immunization; M2, female mice on parturition day; TT4, total thyroxine; TSH, thyrotropin; SD, standard deviation.
TgAb titer, MCT8, RC3, and Reelin mRNA expression in pups
TgAb levels of pups in the T group at PND0, PND10, and PND40 were significantly higher than those from the C group (PND0: p=0.001; PND10: p=0.001; PND40: p=0.02), respectively. No statistical difference in TgAb titer was seen between PND0 and PND10 (p=0.351) in the T group. However, both groups had higher OD ratios than at PND40, p=0.00. Because determination of thyroid hormones in the brains of pups was not available, thyroid hormone responsive genes were analyzed; specifically, RC3, Reelin, and the thyroid hormone transporter MCT8 mRNA were determined in the whole brains of pups. The results showed that the expression of all three mRNAs had no statistical difference at PND0, PND10, and PND40 (p>0.05) between the T and C groups. These results indicate that maternal TgAb could transfer to pups, and with the extension of time, the TgAb titer of pups decreased significantly. Moreover, the maternal TgAb did not affect thyroid hormones levels or transfer to the brain in the pups (Fig. 2).
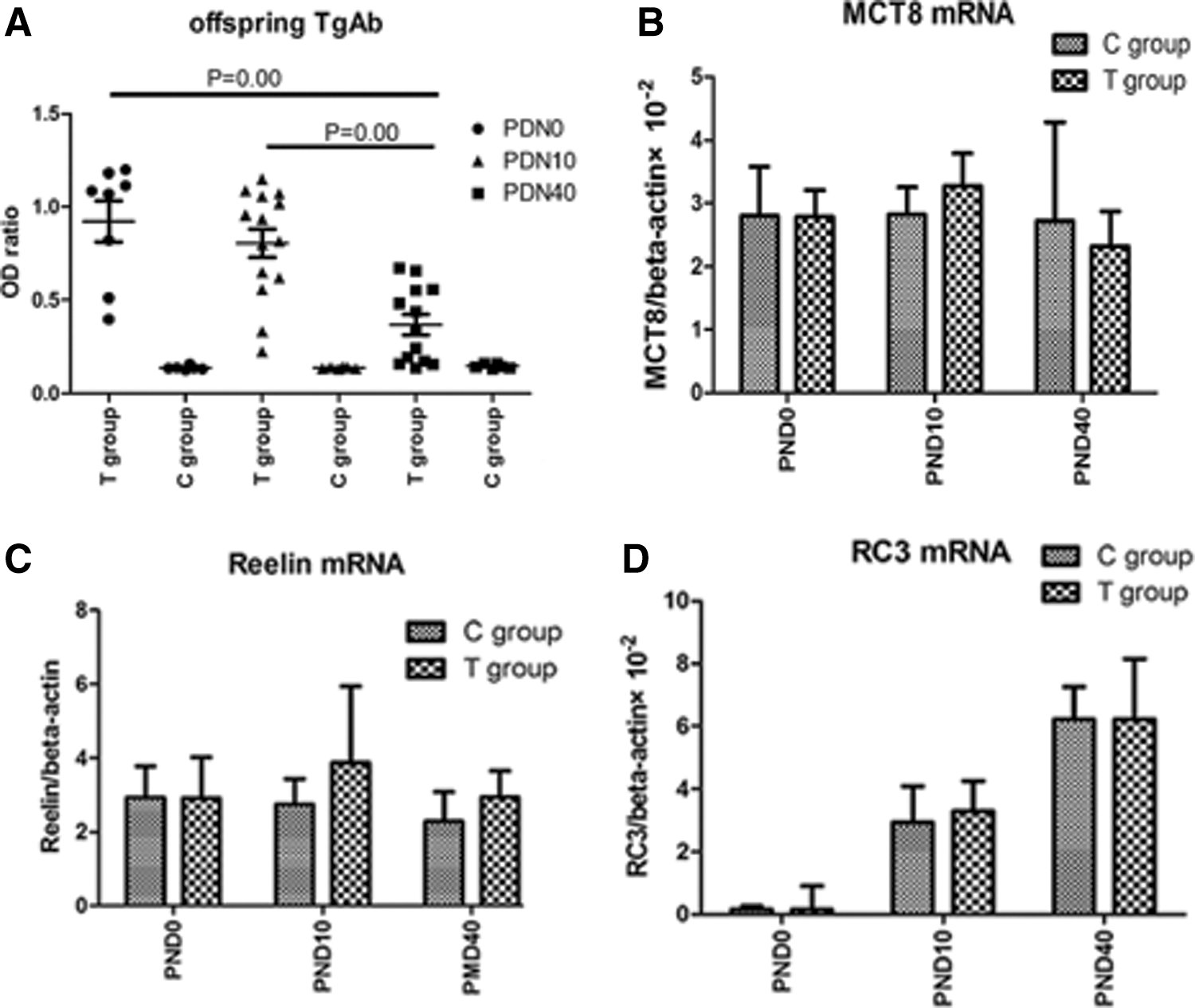
(
Brain histology and nerve cell migration
Nissl staining showed that in C group pups, the brain had well-arranged neurons in the cortex and hippocampus, a clear structure, normal size, and clear nucleoli. Compared to the C group, T group pups had no obvious difference. The thicknesses of cortex neurons and CA1 pyramidal neurons (PY) had no statistical differences either (p>0.05; Fig. 3). To determine whether maternal TgAb affected neuronal migration, BrdU IHC was performed. BrdU positive cells had claybank nuclei, which were richly expressed in the cerebral cortex and hippocampus. Photomicrographs of the immunohistochemistry-stained brain tissues at PND40 were used to calculate the IOD ratio of BrdU-labeled cells in every layer of the cortex, the CA1 pyramidal neurons of hippocampus (PY), and the radiation area (RA). When comparing the T group with the C group, no significant statistical differences were found in the percentages of positive cells (p>0.05; Fig. 4).
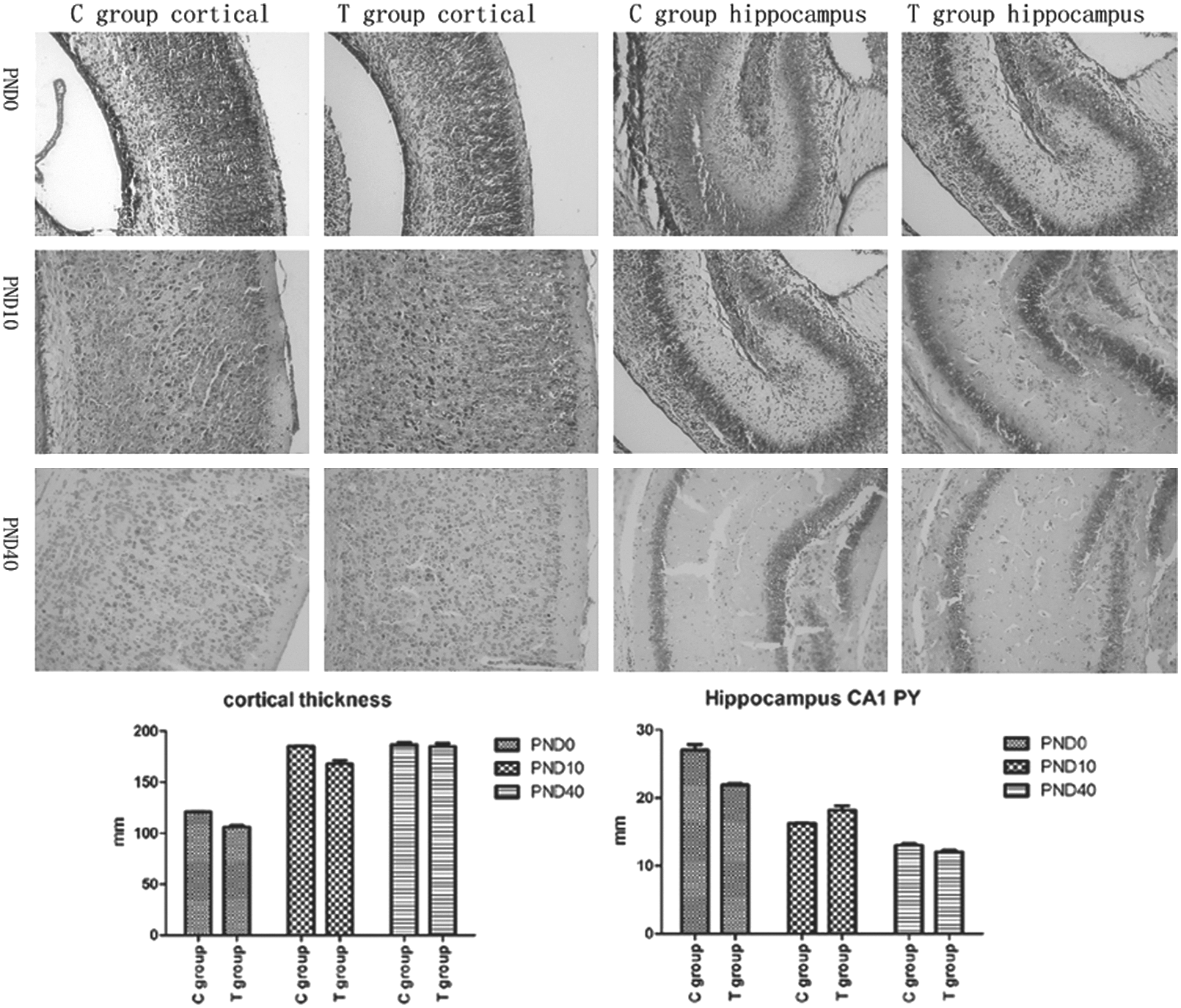
Nissl staining in offspring brain, magnification×200. Bar graphs show thickness of cerebral cortex and hippocampus of pups using Nissl staining. Every sample measured twice in magnification×200, p>0.05. PY, pyramidal layer of hippocampus; RA, radiation area of hippocampus.
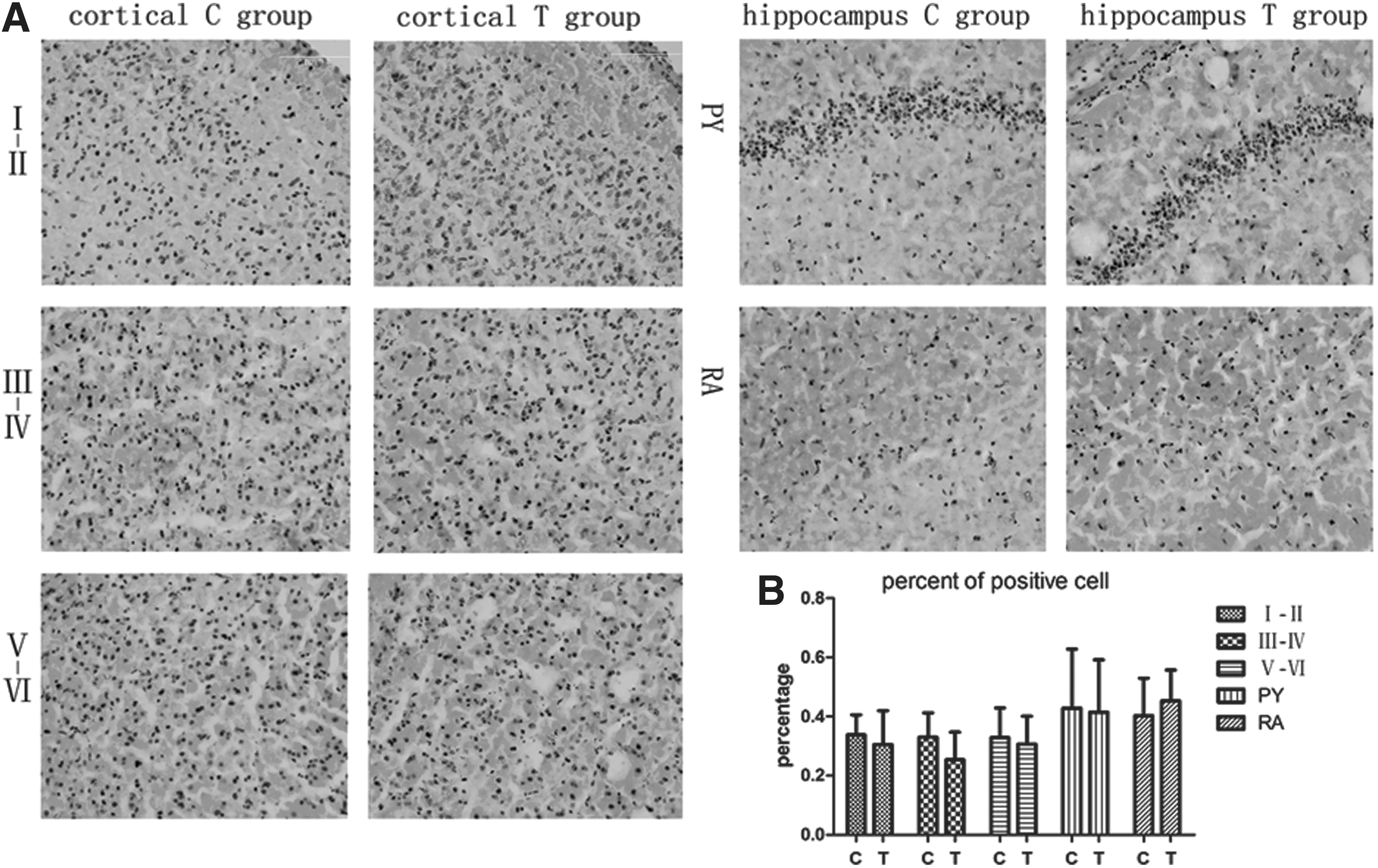
(
Morris water maze
The Morris water maze is a classic test for evaluating spatial learning and memory ability for rodents. The escape latency time of all training groups became shorter as training days increased. Meanwhile, there was no significant statistical difference between the T and C groups after five days of training (p>0.05). A probe trial test recorded the number of times each mouse crossed the platform area: the C group crossed 2.6±1.83 times, and the T group 3.37±0.91 times in 60 sec. The results suggest that no difference exists between the two groups (p>0.05; Fig. 5).
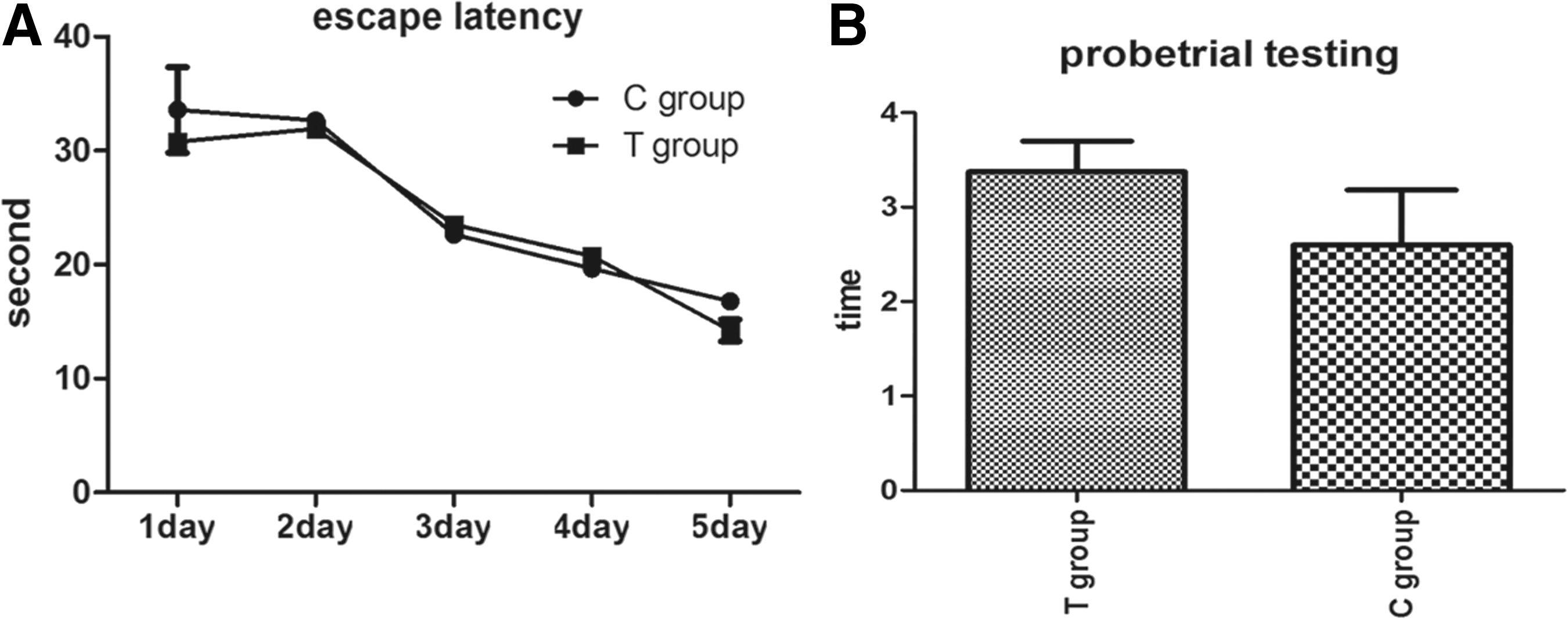
Performance of PND40 pups in the Morris water maze test. Data were expressed as the mean±SD (C group, n=10; T group, n=8). (
LTP and its related factors
Learning ability and memory, characterized as the LTP of fEPSPs, require synaptic transmission. The amplification percentages of the slope fEPSPs of the two groups were compared, and the findings were: the fEPSP % slope at baseline (n=10) in C group pups was 213.59±73.24%, and the fEPSP % slope in T group pups was 167.72±22.71% (n=8). There was no significant difference between the two groups (p=0.17), but in contrast to the C group, the T group values showed a downward trend. Both Phospho-ERK1/2 and BDNF played important roles in induction and maintenance of LTP (11). According to the Western blot and real-time PCR test, both ERK1/2 protein and BDNF mRNA expression showed no significant difference between the two groups (p>0.05; Fig. 6).
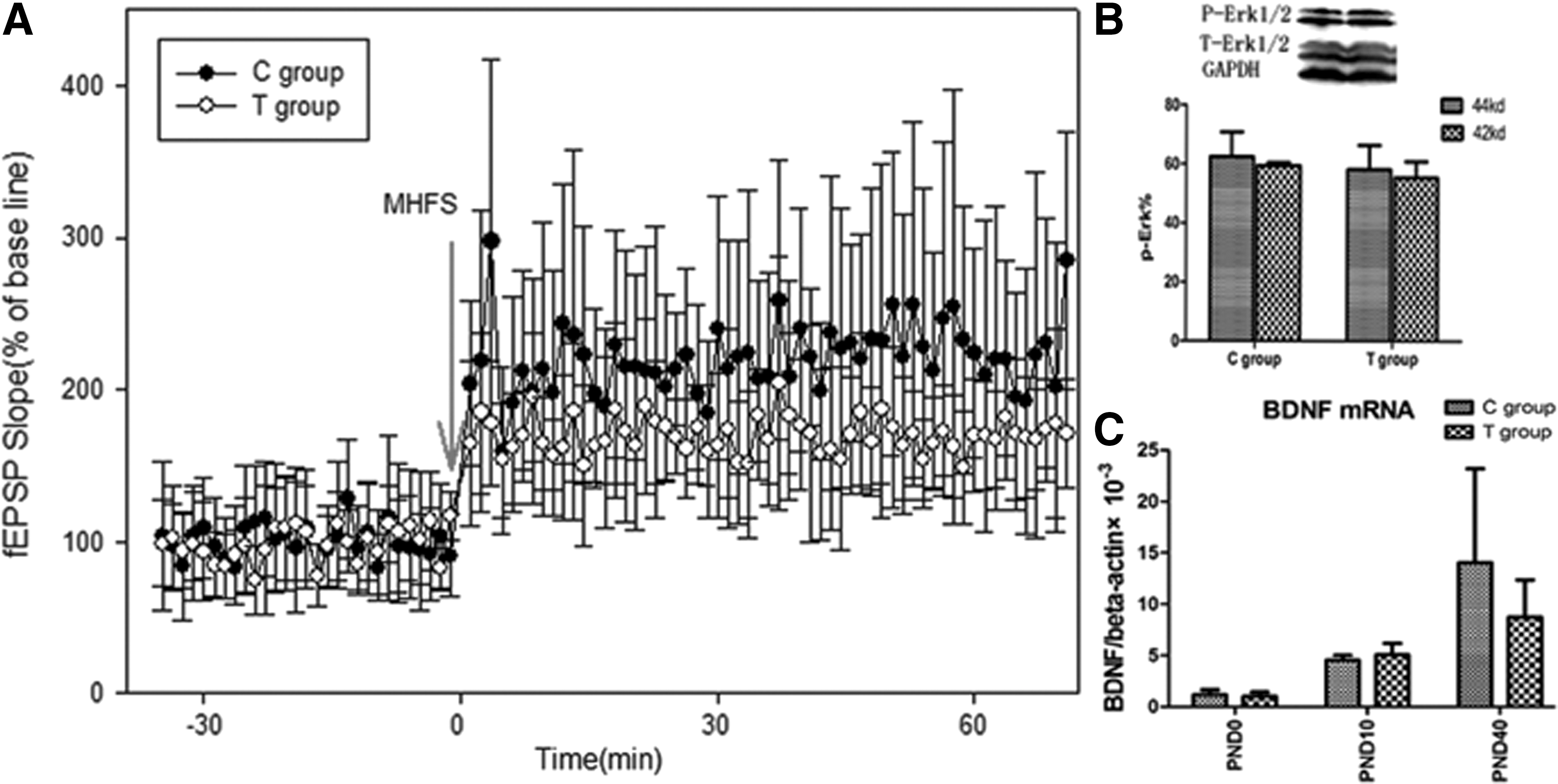
(
Discussion
EAT is a classic method for studying autoimmune thyroiditis. It is induced with mTg, a known thyroid autoantigen that is associated with the development of thyroiditis in both mice and humans. There are two parameters to evaluate EAT: one is histologic evaluation, which manifests mononuclear cell infiltration into the thyroid gland, and another is the serum thyroid antibody level increase (5). In the present study, the isolated maternal TgAb positive model met the above two parameters. To the authors' knowledge, this study is the first to use the model to explore the brain development in offspring.
After detecting TgAb titers in pups, it was found that pups were more likely to suffer from higher TgAb levels when their mothers had AIT. Nevertheless, TgAb titers decreased significantly as time progressed. This indicates that the increased TgAb of offspring originated from maternal antibodies transferred to the fetus through the placenta. TgAb, which belongs to the immunoglobulin IgG subtype, can cross the placental barrier (12). However, exogenous antibody levels will decrease gradually in offspring (13). This is confirmed, for example, by an epidemiological study from Korea (14).
Through behavioral experiments including the Morris water maze and electrophysiological experiments (LTP), it was found that the TgAb positive model did not affect the learning ability and the memory of offspring. The negative results led to this mechanism being studied further.
First, maternal thyroid hormones play an important role in early brain development in mammalian embryos (15). Two T3-responsive genes, Reelin and RC3 (16), were analyzed, and the results indirectly confirmed that thyroid hormone levels in the brain were normal in pups of EAT mice. MCT8 is one of the specific thyroid hormone transmembrane transporters and plays a major role in uptake of thyroid hormones across the blood–brain barrier, as well as in the transport between cells such as astrocytes and neurons within the brain (17). Unlike in humans, isolated MCT8 deficiency does not result in neurological impairment in the mouse, whereas combined MCT8 and OATP1C1, which mediate the uptake of thyroid hormones jointly, results in impaired neurological development (18). The present study did not find evidence that EAT had MCT8 deficiency, and OATP1C1 was not specifically studied.
Second, nerve cell migration is also an important aspect of brain development. Reelin and BrdU tests are classic methods of detecting nerve cell migration. Reelin is important for the correct positioning of neurons during the development of the brain cortex (19). Through the results of morphological and genetic detection, it was found that a TgAb positive model did not influence brain morphology and nerve migration of offspring.
Third, assessing learning ability and memory through the Morris water maze and LTP are classic methods (20,21). There are many pathways included in LTP where ERK1/2 appears to act as a point of convergence for several signaling cascades. The multiple and varied substrates of ERK1/2 predict the array of changes that follow its activation (22). Additionally, RC3 is highly concentrated in the cytoplasm and dendrites of selective neurons within the forebrain. It can enhance LTP and learning by promoting calcium-mediated signaling (23). BDNF is also a growth factor that increases dendritic spines and enhances memory function, and it plays an important role in learning and memory (24). In the present study, isolated maternal TgAbs had no effect on the expression of the selected analyzed genes and proteins that are involved in learning and memory pathways.
Clinical studies found AIT to be associated with abortion and adverse pregnancy outcomes (25,26). There was no consistent conclusion whether maternal thyroid antibodies influence the brain development in offspring. The present animal experiment shows that isolated positive maternal TgAb did not affect the brain development of offspring. However, the study has some limitations. The levels of TgAb within the immune group were variable, and we did not evaluate potential differences among the offspring by stratifying TgAb titers. In addition, the study focused on the hippocampus and cerebral cortex, but other parts of the brain such as the cerebellum play essential roles in some types of motor learning and cognitive functions. These limitations will be the focus of future studies.
In summary, isolated maternal TgAb did not have an obvious influence on learning ability and memory of offspring, or nerve cell migration in an murine model of EAT, despite a transient TgAb increase in offspring.
Footnotes
Acknowledgments
This work was supported by the 973 Science and Technology Research Foundation, Ministry of Science and Technology in China (Grant 2011CB512112); the Chinese National Natural Science Foundation (Grant 81170730, 30971400); the Program for Liaoning Excellent Talents in University (Grant LR2011022); the Program for the Construction of Endocrine Disease Laboratory (F11-244-1-00); Liaoning Province Science and Technology Plan Projects (2011225023); and the Institutions of Higher Learning in Liaoning Province Innovation Team Project (LT2012015).
Author Disclosure Statement
The authors declare that no competing financial interests exist.