
Abstract
Background:
The B cell activating factor (BAFF) is a member of the tumor necrosis factor family, which controls the survival/proliferation of B cells and is involved in the pathogenesis of a number of autoimmune diseases. The objective of the present study was to investigate the expression of BAFF and BAFF receptor (BAFF-R) in the thyroid tissue of patients affected with autoimmune thyroid disorders (AITD) or multinodular goiter (MNG) compared with those with normal thyroids.
Methods:
Immunohistochemistry was performed using a panel of antibodies against BAFF, BAFF-R, CD3, CD4, CD8, CD20, CD34, CD79a, CD1a, CD68, and CD163 on the thyroid sections of 27 patients affected with Graves' disease (GD), 23 with Hashimoto's thyroiditis (HT), 16 with nontoxic nodular goiter (NTG), and 15 with toxic nodular goiter (TG), submitted to total thyroidectomy between 2000 and 2011.
Results:
The overall BAFF-R expression in thyrocytes was weak and not different in AITD and MNG. Conversely, a stronger BAFF expression was observed in MNG compared with AITD. BAFF and BAFF-R expression in the infiltrating lymphocytes was higher in AITD compared with MNG. Interestingly, in lymphocytes of follicular-like structures observed in HT, BAFF and BAFF-R were localized in the germinal center or in the mantle, respectively.
Conclusions:
This study shows that BAFF and BAFF-R are expressed in the thyrocytes derived from patients with either AITD or MNG, in addition to the expected expression of BAFF and its receptor in the infiltrating immune cells of GD and HT. These findings suggest a possible involvement of BAFF and its receptors in the pathophysiology of AITD.
Introduction
T
BAFF is likely involved in the pathogenesis of a number of autoimmune diseases (2). BAFF overexpression has been reportedly associated with the breakdown of B cell tolerance and autoantibody production. Serum BAFF concentrations are increased in systemic lupus erythematosus (SLE), rheumatoid arthritis, Sjögren's syndrome (SS), autoimmune hepatitis, primary biliary cirrhosis, and Wegener's granulomatosis (3 –6). BAFF is also involved in the repopulation of peripheral B cells after depletion by the anti-CD20 monoclonal antibody rituximab (7,8).
A possible role of BAFF in the pathogenesis of autoimmune thyroid diseases has been suggested by Gilbert et al. (9) who, after immunizing BALB/c mice with a recombinant adenovirus expressing the TSH-receptor A subunit, induced experimental autoimmune hyperthyroidism. In this experimental model, the administration of BAFF-R-Fc, which blocks BAFF activity, induced an improvement of the hyperthyroidism with a significant reduction in serum T4 in 78% of the animals, compared with pretreatment levels, suggesting that BAFF may be involved in the pathogenesis of Graves' disease (GD). In addition, the authors have previously shown that serum BAFF concentrations are elevated in GD patients, independently of the presence of Graves' orbitopathy (GO) (10). Moderately increased serum BAFF concentrations were also observed in patients with HT, although the elevation was less pronounced when compared with patients with GD (11,12).
Previous data obtained in patients with SS, an autoimmune disease that shares histological and clinical similarity with HT, being characterized by B-lymphocyte infiltration and tissue functional impairment (or functional impairment), suggest that BAFF may be responsible for an abnormal distribution of B cell subsets within the salivary gland, associated with an increased number of transitional autoreactive B cells (13). BAFF overexpression is associated with an atypical B cell hyperactivity in SS (14), due to increased expression of co-stimulatory molecules (15).
At present, the tissue expression of BAFF in the thyroid gland of subjects affected with autoimmune disorders is completely unknown. To determine whether BAFF and its specific receptor BAFF-R are expressed in the thyroid of patients affected with autoimmune or nonautoimmune thyroid disorders, immunohistochemistry was performed on 81 thyroid samples from patients affected with GD, HT, or toxic and nontoxic multinodular goiter (MNG).
Materials and Methods
Patients
Thyroid tissue specimens were obtained from 81 patients affected with benign thyroid goiter or AITD, submitted to total thyroidectomy between 2000 and 2011.
Twenty-three women with Hashimoto's thyroiditis (HT) were euthyroid, of whom six were on L-thyroxine (L-T4) replacement therapy at the time of thyroidectomy. Six patients underwent surgery because of TIR3 cytology according to the classification of the Italian Societies of Pathology and Endocrinology (SIAPEC/SIE), with no malignancy found at histology. The remaining patients underwent surgery because of very large goiters causing compressive symptoms or because of cosmetic concerns. Seventeen of 23 HT patients had detectable serum thyroid peroxidase antibodies (TPOAb) and thyroglobulin antibodies (TgAb), respectively. Twenty-seven patients (5 men) were affected with relapsing GD after 12–18 months of methimazole (MMI) treatment and were therefore referred for thyroid surgery. Total thyroidectomy was preferred to radioiodine ablation due to the presence of a large goiter or the patient's choice. All patients were euthyroid on MMI at the time of surgery. Twenty-five of 27 GD patients had associated GO, of whom 12 were previously treated with steroids because of active moderate-to-severe disease. Papillary thyroid cancer <10 mm was incidentally found on histology in the thyroid of four patients with GD. All 27 GD patients had detectable serum thyrotropin receptor antibodies (TRAb), two had TgAb, and seven had TPOAb. Thirty-one patients (6 men) had MNG, 15 of whom had toxic goiter treated with MMI until surgery, while the remaining 16 patients had euthyroid goiters. No antibodies were detected in the serum of MNG patients (Table 1).
Ellissoid formula (4/3 π a × b × c).
Not assessed.
MMI, methimazole; L-T4, L-thyroxine; TgAb, thyroglobulin antibodies; TPOAb, thyroid peroxidase antibodies; TRAb, thyrotropin receptor antibodies.
In addition, 12 thyroid specimens without any histological abnormalities, obtained from the contralateral lobe of patients operated for papillary thyroid carcinomas (PTC), were studied as controls.
This study was approved by the local Ethics committee, and informed consent was obtained from all the patients. The clinical characteristics of the study patients are shown in Table 1.
Macroscopic analysis of surgical samples, histology, and immunohistochemistry
The volume of surgical thyroid samples was quantified by the ellipsoid formula (4/3 π h × w × d of each thyroid lobe). Thyroid tissue specimens were formalin fixed and paraffin embedded. Serial tissue sections (4 μm) were dewaxed, rehydrated, and stained with hematoxylin and eosin staining for histological analysis.
For immunohistochemistry, antigen retrieval was performed by treating the slides in a thermostatic bath (97.5°C) in citrate (pH 6) or EDTA (pH 8) buffer for 35 min. Endogenous peroxidase activity was quenched with 3% H2O2. After incubation in blocking buffer, the following primary antibodies were applied: CD3 (clone F7.2.38; dilution 1:30), CD20 (clone L26; dilution 1:300), CD79a (a pan-B marker clone HM57, dilution 1:40), CD8 (clone C8/144B; dilution 1:50), CD4 (clone 1F6; dilution 1:200), all supplied by DAKO Cytomation (Carpinteria, CA). BAFF and BAFF-R antibodies were supplied by ProSci, Inc. (San Diego, CA). The reaction was detected by the Novolink Max polymer detection system (Novocastra Laboratories Ltd., Leica Microsystem, Nussloch, Germany), following the manufacturer's instructions. Slides were counterstained with hematoxylin, mounted, and observed under a light microscope (Nanozoomer XR Hamamatsu). Expression of BAFF and BAFF-R was defined as the number of stained cells graded as 0, 1+, 2+, 3+, and 4+, when no cells, <5, 5–10, 10–30, and >30 cells, respectively, were counted per high magnification field (250 × ). The specificity of BAFF and BAFF-R antibodies was preliminary tested on a hypertrophic tonsil.
Flow cytometry leukocyte and thyrocyte characterization
Infiltrating leukocyte characterization was performed on fresh dissociated thyroid tissues obtained from four MNG, one HT, and three GD patients.
BAFF-expressing cells were identified with the following fluorochrome-conjugated monoclonal antibodies: BAFF-PE and BAFF-R-PE (Biolegend, Inc., San Diego, CA), CD19-PE-cy7, CD14-PE-cy7, CD4-APC, CD8-FITC, CD45-APC-cy7, cytokeratin 8 (CK8-FITC, a specific epithelial marker) supplied by Becton Dickinson Biosciences (BD; San Jose, CA). 7-Amino Actinomycin-D, 7-AAD (BD) was used to exclude dead cells from the analyses. To evaluate intracellular markers, cells were permeabilized with 0.5% saponin for 30 min at room temperature before staining. The fluorescence threshold between negative and positive cells was set on the basis of the reactivity of appropriate nonspecific fluorochrome-conjugated isotypical controls. Data were acquired and analyzed with a FACSCanto II flow cytometer equipped with a FACSDiva software (BD).
The specificity of the BAFF and BAFF-R antibodies was first tested on the whole blood of a healthy donor and of a patient treated with rituximab.
Primary thyrocytes were separated for BAFF and BAFF-R mRNA expression by means of fluorescence-activated cell sorting. Briefly, primary live thyrocytes were defined as CD45–/7AAD–/CK8+ cells and sorted using a FACSAria II equipped with a FACSDiva software (BD).
RNA extraction and expression
Total RNA was extracted with Trizol reagent according to the manufacturer's instructions, and reverse transcribed to cDNA with the RevertAid First Strand cDNA Synthesis Kit (Thermo Scientific, Waltham, MA). The following primers were used to amplify either BAFF or Δ-BAFF (splice variant lacking exon 3): BAFF forward: 5′-CAG AAA TAA GCG TGC CGT TC-3′; BAFF reverse 5′-GTC CCA TGG CGT AGG TCT TA-3′. Reverse transcription polymerase chain reaction (RT-PCR) was resolved in 3% agarose gel stained with EtBr 0.5 μg/mL. N-THY cells, which are normal human primary thyroid follicular epithelial cells immortalized with SV40 ori plasmid that retain the functions of normal thyrocytes, were kindly provided by Dr. M.G. Borrello (Fondazione IRCCS, Istituto Nazionale dei Tumori, Milan, Italy).
Cell culture
N-THY cells were cultured in RPMI medium with phenol red, supplemented with 10% fetal bovine serum (FBS) and maintained in a 5% CO2–95% air atmosphere at 37°C with a change of medium every second or third day, and passed every seven days. Cells were seeded in 6-well plates and grown for 24 h in RPMI medium with 10% FBS. rhTSH (Thyrogen®, supplied by Genzyme, San Diego, CA) was added to the media at a final concentration of either 10 ng/mL or 20 ng/mL as previously described (16). Forty-eight hours after stimulation, cells were trypsinized, and BAFF and BAFF-R expression was analyzed by cytofluorimetry. Each condition was tested in triplicate.
Results
Expression of BAFF and BAFF-R in thyrocytes and lymphocytic infiltrates
In control thyroid specimens, no staining was found for BAFF or BAFF-R (Fig. 1A and B). In contrast, significant BAFF expression of various staining grades was found in thyrocytes from MNG and AITD and the lympho-monocytic cells infiltrating the thyroid (Fig. 1). Of note, specific immunoreactivity was observed both as cytoplasmic or perinuclear staining. Interestingly, a greater expression of BAFF was noticed on the thyrocytes from MNG compared with either GD or HT thyroid tissues (Table 2 and Fig. 1). Specifically, grade 4+ BAFF staining was found in 25/31 MNG, 8/27 GD tissues, and 11/23 HT thyroid tissue sections (Table 2). The expression of BAFF on GD did not differ from that on HT thyrocytes (Table 2). Since the expression of BAFF in the thyrocytes was a novel finding, other experiments were performed to confirm these results. First, immunohistochemistry was repeated using a different antibody, a mouse monoclonal antibody (Mouse Monoclonal anti-BAFF antibody clone T7-241 from LifeSpan BioSciences, Inc., Seattle, WA), and a similar pattern of staining was found. In addition, mRNA expression of BAFF in N-THY SV40 ori cells and primary thyrocytes was studied, sorted by flow cytometry as CK8+ cells.
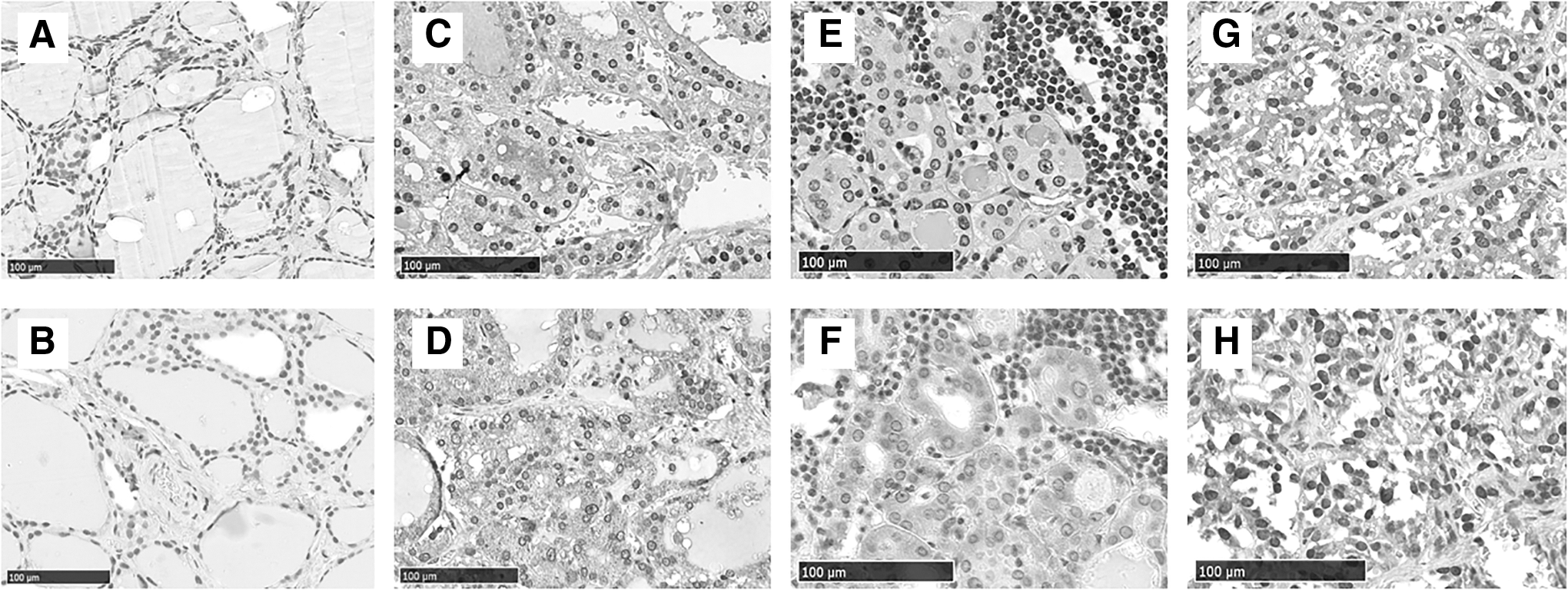
B cell activating factor (BAFF) and BAFF receptor (BAFF-R) expression in different thyroid diseases. Four representative cases of BAFF (upper row) and BAFF-R (lower row) stainings in different thyroid disorders: normal thyroid
The number of cases with different grades of BAFF and BAFF-R expression is summarized. Positive staining was graded as 0, 1+, 2+, 3+, and 4+, when no cells, <5, 5–10, 10–30, and > 30 cells, respectively, were counted per high magnification field (250 × ).
MNG, multinodular goiter; GD, Graves' disease; HT, Hashimoto's thyroiditis.
The presence of BAFF transcripts was confirmed in both cell types. Interestingly, N-THY cells mostly expressed the full-length BAFF transcript compared with ΔBAFF, a splice variant lacking exon 3, whereas primary thyrocytes equally expressed both variants (Fig. 2). Finally, flow cytometry was performed on a limited number of specimens. In the first six samples studied, no BAFF staining was found on the membrane of the thyrocytes (CK8+ cells), which became BAFF positive only after permeabilization with 0.5% saponin, suggesting the presence of intracellular staining. The lack of membrane BAFF staining was unexpected. Therefore, the dissociation protocol was modified, since collagenase digestion has been reported to alter the membrane expression of a number molecules. Consequently, the collagenase concentrations and the time of digestion were reduced. At the time of surgery, margins of resection are routinely stained with India ink followed by immersion in Bouin's fixative. By avoiding this step in the two additional surgical specimens analyzed, an increase of BAFF membrane staining was observed in both MNG and GD in 49–70% of CK8+ cells without the need for permeabilization. BAFF expression was also observed on the membrane of 6–11% of CD8 and on 2–6% of CD4 T lymphocytes in both GD patients and on 3% of B lymphocytes in the single HT patient examined, whereas lympho-monocytes infiltrating nodular goiters did not express BAFF either on the membrane or at the intracellular level. A weak BAFF-R staining was found to be expressed in the cytoplasm of thyrocytes from all tissue specimens (Table 2 and Fig. 1).
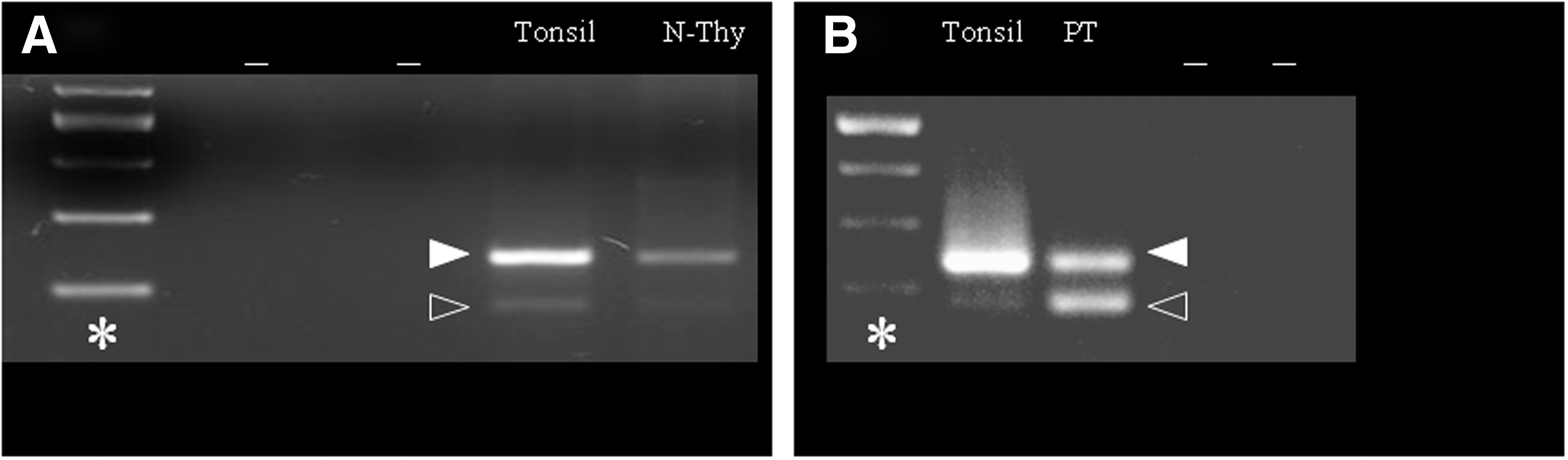
BAFF expression in N-Thy cells and primary thyrocytes. BAFF expression in N-Thy cells (
As expected, HT and GD tissues samples were infiltrated with more lymphocytes (>30 per high field magnification) when compared to samples from MNG, in which only few or no lymphocytes were found. While no BAFF or BAFF-R expression was found in the infiltrating cells of 22/31 (71%) MNG samples, with infiltration mainly characterized by macrophages and only few detectable T and B cells, grade 1+ to 4+ staining was found in all cells infiltrating GD and HT thyroids (Table 2). Lymphocytes from HT thyroids showed both a diffuse and a focal pattern of infiltration, the latter characterized by follicular-like structures (FLS) as commonly described, whereas in GD, lymphocytic infiltration was always diffuse (Fig. 1G and H). HT thyroids were characterized by marked B lymphocyte infiltration, either diffuse or in FLS, with greater expression of BAFF in the germinal center and of BAFF-R in the mantle. GD thyroids showed predominant T and B lymphocytic infiltration.
With cytofluorimetry, BAFF-R was found in nearly 100% of B lymphocytes (CD19+ cells) in all the samples. In cultured N-Thy cells, a mean BAFF and BAFF-R positive expression was found in 4.7% and 0.1% of cells, respectively. The incubation with 10 or 20 ng/mL of hrTSH did not significantly change the expression of BAFF or BAFF-R compared with unstimulated cells.
BAFF and BAFF-R expression by immunohistochemistry and association with the clinical phenotype
Overall, BAFF and BAFF-R expression in thyrocytes and lymphocytes did not differ in men and women, and did not correlate with the patients' age, thyroid volume, or weight. In GD thyroid tissue, BAFF and BAFF-R expression in thyrocytes and infiltrating cells did not correlate with either disease duration or serum TRAb levels. In 12 patients with GD previously treated with steroids for active GO, BAFF and BAFF-R expression did not show any relationship with the time elapsed from immunosuppressive treatment or with treatment itself.
Discussion
In this study, a novel finding is reported that BAFF and BAFF-R are expressed in the thyrocytes derived from patients with either AITD or MNG, in addition to the expected increased expression of BAFF and its receptor in the infiltrating immune cells of GD and HT.
The expression of BAFF and its receptors on nonlymphoid tissues has been previously observed in adipocytes at different stages of differentiation from immature to mature adipose cells (17,18). Constitutive expression of BAFF has also been observed in the mammary gland (19) in both normal and neoplastic epithelial tissue and in the surrounding adipocytes. BAFF has also been found in the salivary ductal epithelium from normal subjects and from patients affected with SS (20,21), in normal and neoplastic renal tubules or Malpighian corpuscles (22), in the synoviocytes (23) of patients affected by rheumatoid arthritis, and in lower airway epithelium (24). BAFF and BAFF-R are expressed in mouse neurons (25), and BAFF-R deficiency has been reported to decrease the survival of primary cultured neurons.
In the present study, BAFF staining in thyrocytes was found in the cytoplasm, but was absent or very weak on the plasma membrane. At cytofluorimetry, BAFF expression on the cells surface was variable from 49% to 90% in the two samples analyzed. An explanation for this variable expression may be the intracellular sequestration of BAFF. It was found that in primary thyrocytes, the expression of full-length BAFF mRNA transcripts is co-expressed along with a shorter isoform, called ΔBAFF, which lacks 57 nucleotides of exon 3. It has been previously shown by Gavin et al. (26) that ΔBAFF can associate with BAFF in heteromultimers, limiting BAFF homotrimerization and its expression on the cell surface, by a mechanism of competitive co-association. In particular, this has been associated with the retention of ΔBAFF/BAFF multimers inside the cells. A similar mechanism has been evoked for other family members, such as TNF, which is tightly regulated by receptors being kept in intracellular stores (27).
Surprisingly, in the present study, a greater expression of BAFF wes found in the thyrocytes of MNG when compared with AITD. This finding does not seem to be related to antithyroid drug treatment, since BAFF expression in the infiltrates and thyrocytes from toxic and nontoxic MNG was similar irrespective of whether the patients were treated with methymazole. In addition, BAFF and BAFF-R expression in the thyrocytes does not seem to be stimulated by the TSH receptor, since incubation with rhTSH did not significantly change the expression of BAFF and BAFF-r in cultured N-thy cells. Since BAFF has been related to intracellular anti-apoptotic signals in neurons and adipose tissue (22,25), one could speculate that the increased expression observed in MNG might be related to the tissue hyperplasia and the cellular growth characteristic of the goiter. This is also consistent with the lack of BAFF expression in normal thyroid tissue derived from the contralateral lobe of patients with PTC. The relatively reduced expression of BAFF in AITD, compared with MNG, may be explained by the predominance of factors that promote tissue disruption and atrophy (such as lymphocytic infiltration, and immune reactions against thyroid antigens).
Interestingly, the expression of BAFF in thyrocytes seems higher in tissues with a lower number of BAFF-expressing infiltrating lymphocytes. These preliminary observations might suggest a sort of paracrine/endocrine loop between the thyroid tissue and the immune cells. In particular, locally increased BAFF secretion by T-lymphocytes, macrophages, and dendritic cells might downregulate BAFF expression in the thyrocytes, following BAFF binding to its specific receptor, also expressed on thyrocytes. In addition, the elevated serum concentrations of BAFF, reported in both GD (10) and HT patients (11), may reduce BAFF expression in the thyrocytes of AITD. In humans, a similar paracrine cross-talk between the epithelium and the infiltrating cells has been described in prostate cancer tissues by Di Carlo et al. (28), who found reduced levels of BAFF in neoplastic tissue, leading to a reduced stimulation of the infiltrating B cells, thereby impairing immune surveillance against dysplastic and neoplastic epithelial cells. In vitro studies suggest that increased BAFF expression in the epithelium derived from the salivary glands allows survival of adjacent infiltrating B cells (22), thus representing a possible trigger for the development of SS. In autoimmune diseases, the relationship between BAFF expression in epithelial cells and immune infiltrates is still controversial. Daridon et al. (21) found an increased expression of BAFF in the total lysates of the salivary gland of patients with SS when compared with healthy controls without distinction between epithelium and infiltrates. In contrast, a study by Vosters et al. (20) found a similar BAFF expression in patients with SS, healthy volunteers, and patients with symptoms of sicca syndrome.
Flow cytometry has shown that, consistent with what has been reported in other autoimmune diseases, BAFF expressing infiltrating cells in AITD were mostly T lymphocytes, while BAFF-R expression was restricted to B lymphocytes. In only one HT thyroid sample expression of BAFF detected in 3% of B lymphocytes. This finding is somehow consistent with what has been observed in the infiltrates of the salivary gland of SS (21) and in hematologic neoplasias. In these tissues, BAFF seems to be atypically secreted by malignant or autoreactive B cells (21) that may confer increased survival potential to leukemic and autoimmune B cells. An additional similarity to SS is that, in HT, the FLS infiltrates express BAFF in the germinal center and BAFF-R in the mantle (29). The present study suggests that BAFF and BAFF-R are expressed in thyrocytes derived from patients with either AITD or MNG, in addition to the expected increased expression of BAFF and its receptor in the infiltrating immune cells of GD and HT. This finding, together with previous reports of increased serum BAFF levels in patients with AITD, suggests that BAFF may play a role in the pathogenesis of AITD. Activation of the BAFF–BAFF-R system may support the rationale for the use of B cell targeted therapies in patients with GD and GO (30).
Footnotes
Acknowledgments
This work is supported in part by MURST, Rome, and by Fondazione Cà Granda, IRCCS, Milan, Italy.
Author Disclosure Statement
The authors declare that there is no conflict of interest that could be perceived as prejudicing the impartiality of the research reported.