
Abstract
Background:
Thyroglobulin (Tg), the major thyroidal protein, plays important roles in thyroid hormone biosynthesis and in autoimmune thyroid diseases (AITD). Tg also serves as a pre- and postoperative biomarker of differentiated thyroid cancer (DTC). The endogenous β-galactoside binding lectin galectin-3 (Gal-3), secreted by transformed thyroid cells, has been shown to be another useful biomarker of DTC. Tg contains covalently linked complex-type glycans that can serve as binding epitopes of Gal-3. The objective of the study is to investigate the interaction between Tg and Gal-3 and discuss its potential consequences.
Methods:
Binding interaction between Tg and Gal-3 was first studied by hemagglutination inhibition assays. Subsequently, a detailed analysis of binding thermodynamics was carried out by isothermal titration calorimetry. Quantitative precipitation was performed to study the complex formation between Tg and Gal-3 and to determine the binding stoichiometry. The concentration-dependent rate and amount of complex formation between Tg and Gal-3 was examined spectrophotometrically. A similar approach was taken to study the effect of free Tg and Gal-3 on preformed Tg–Gal-3 complex.
Results:
Quantitative biochemical and biophysical data show that these two biomarkers produced by thyroid cancer cells interact with each other with submicromolar affinity and form an insoluble complex at their stoichiometric concentration. One Tg molecule could bind up to 14 molecules of Gal-3. Such complex formation mutually sequestered both Tg and Gal-3, decreasing the concentration of their freely available forms. Formation of the Tg–Gal-3 complex was reversible as the preformed complex was dissolved by free Tg as well as free Gal-3. While free Tg rapidly dissolved preformed Tg–Gal-3 complex in a concentration-dependent manner, Gal-3 was found to be much less efficient and slowly dissolved only a fraction of the preformed complex at a relatively higher Gal-3 concentration.
Conclusions:
Complex formation between Tg and Gal-3 through high affinity binding and the sensitivity of the complex to free Tg and Gal-3 can potentially influence their biological functions. Interactions between Tg and Gal-3 might also interfere with their clinical detection, the same way Tg autoantibody (TgAb) is reported to interfere with Tg assays. The data support a model of Gal-3-mediated homeostatic process of Tg.
Introduction
T
Due to its cancer-associated expression, the endogenous β-galactoside binding lectin galectin-3 (Gal-3) is a promising marker of thyroid carcinomas (12 –17). The levels of Gal-3 in fine-needle aspiration (FNA) samples and in serum samples have been investigated in many studies (18 –21). Immunohistochemical analysis revealed the sensitivity of Gal-3 as a thyroid cancer biomarker (22). A test method has been proposed (Gal-3 thyrotest) to enhance the diagnostic accuracy of thyroid FNA cytology. This method determines the expression level of Gal-3 in FNA-derived cells (23).
Gal-3 recognizes complex-type glycans terminating in either α2–3 or α2–6 residues. Gal-3 is capable of binding internal LacNAc (N-acetyllactosamine) of poly-LacNAc chains of complex-type glycans (24,25). Tg contains multiple copies of such glycans as covalent attachments. Therefore, Tg appears to be a potential multivalent ligand of Gal-3. The present study found that Gal-3 interacts with Tg with high affinity. This multivalent glycan-dependent interaction between Tg and Gal-3 leads to the formation of reversible cross-linked complexes. The addition of free Tg or Gal-3 dissolves the complex in a concentration-dependent manner. Given the physiological roles of Tg and Gal-3, their clinical importance as biomarkers, and their physical proximity after their synthesis, these interactions might have important clinical and physiological significance, as discussed.
Materials and Methods
Reagents and glycoproteins (including Tgs) were obtained from Sigma Chemical Co., unless mentioned otherwise. Tg samples (with highest purity available) were extensively dialyzed against 20 mM phosphate-buffered saline (PBS) containing 150 mM NaCl. Recombinant human Gal-3 was provided by Prof. Hans-Joachim Gabius.
Subunit Gal-3 concentrations were determined spectrophotometrically at 280 nm using specific extinction coefficient of the protein (E1%1cm). A value of 6.1 (E1%1cm) was used for Gal-3, as determined from isothermal titration calorimetry (ITC) experiments described previously (26). The molecular mass of the Gal-3 subunit was 29,000 Da.
Hemagglutination inhibition assay
The assay was performed at room temperature using a twofold serial dilution technique and 2% (v/v) rabbit erythrocytes in 20 mM PBS containing 150 mM NaCl. The minimum concentration of human Tg (hTg), bovine Tg (bTg), and porcine Tg (pTg) required for complete inhibition of three hemagglutination doses was determined. Inhibition of Gal-3 by complex-type glycan-containing glycoproteins asialofetuin and transferrin were used as positive controls. RNaseB, a glycoprotein that contains exclusively high mannose glycans, was used as a negative control.
ITC
ITC experiments were performed using a VP-ITC instrument from Microcal, Inc. (Northampton, MA). Injections of 4 μL of a glycoprotein solution were added from a computer-controlled microsyringe at an interval of 4 min into the sample solution of lectin (cell volume 1.43 mL) with stirring at 310 rpm. An example of an ITC experiment is shown in Figure 1 for hTg and hGal-3 at 27°C. Control experiments were performed by making identical injections of Tg into a cell-containing buffer. Titrations were carried out at pH 7.4 using 20 mM PBS buffer. The experimental data were fitted to a theoretical titration curve using software supplied by Microcal, with ΔH (binding enthalpy in kilocalories per mole), Ka (association constant), and n (number of binding sites per monomer) as adjustable parameters. The quantity c = KaMt(0), where Mt(0) is the initial macromolecule concentration, is important in titration microcalorimetry. All experiments were performed with c values between 1 and 200. The instrument was calibrated using the calibration kit containing ribonuclease A (RNaseA) and cytidine 2′-monophosphate (2′-CMP) supplied by the manufacturer. Thermodynamic parameters were calculated from the Gibbs free energy equation ΔG = ΔH – TΔS = RT ln Ka, where ΔG, ΔH, and ΔS are the changes in free energy, enthalpy, and entropy of binding, respectively. T is the absolute temperature, and R = 1.98 cal/mol/K.

Binding interaction between human thyroglobulin (hTg) and human galectin-3 (hGal-3) studied by isothermal titration calorimetry at 27°C (pH 7.4). The top panel shows data obtained for automatic injections (4 μL each) of hTg into hGal-3. The integrated curve shows experimental points, and the best fit is shown in the bottom panel.
Multivalent binding often causes precipitation of the reactants. Such precipitation during an ITC experiment adversely affects the titration as well as the quality of the data. Previously, it has been shown that unambiguous ITC data of multivalent binding could be obtained by using low concentrations of lectins and their glycoprotein ligands (27). Tg contains multiple complex-type glycans. Therefore, it is a multivalent glycoprotein ligand for Gal-3. Interaction between Tg and Gal-3 leads to insoluble complex formation at stoichiometric concentration. Following the previous approach (27), the complex formation was arrested. As a result, unambiguous ITC data were obtained (Fig. 1 and Table 1).
The data represent the mean value and standard errors of a particular parameter obtained from three different experiments. Ka, −ΔH, and n values are the mean values of respective parameters from three experiments. −ΔG and −TΔS were calculated separately for each experiment, and then the mean values of −ΔGs (from three experiments) and TΔSs (from three experiments) were determined.
Tg, thyroglobulins.
Quantitative precipitation assays
The assays were performed in 20 mM PBS, pH 7.4, containing 0.15 M NaCl. Increasing amounts of hTg or bTg were added to a series of tubes containing equal amount of hGal-3. The precipitation was allowed to occur for 5–20 h at room temperature. The optical density of the samples in each tube was measured at 420 nm. Lactose solution (200 mM) was added to the precipitates to check if the precipitation was glycan mediated. Experiments were repeated three times.
Kinetics of precipitation
Measured volumes of hTg (or bTg) and hGal-3 solution at different ratios were mixed in a 1 mL quartz cuvette, and the time-dependent development of turbidity was measured at 420 nm. The buffer was 20 mM PBS (pH 7.4). All experiments were done at room temperature and repeated at least three times. Absorbance was monitored continuously until it remained constant for 30 min. After each experiment, a portion of the precipitate was treated with 200 mM lactose solution to check whether the precipitation was glycan dependent.
Kinetics of dissolution of preformed Tg–Gal-3 complex
The Tg–Gal-3 complex was generated by mixing the components at their stoichiometric concentration. A measured amount of hTg (or bTg) solution was added to the preformed Tg–Gal-3 complex in a 1 mL quartz cuvette. Similarly, Gal-3 with known concentration was added to the preformed complex in separate experiments. The time-dependent dissolution of the complex was measured at 420 nm. All experiments were repeated three times.
Results
Gal-3 binds to human thyroglobulin with high affinity
The present study found that hGal-3 binds to hTg with high affinity. Binding of Gal-3 to hTg, bTg, and pTg was studied by hemagglutination inhibition assays and ITC (Fig. 1). The association constants (Ka) obtained through ITC of Gal-3 for hTg, bTg, and pTg are 1490 (M−1 × 10−4), 1097 (M−1 × 10−4), and 423 (M−1 × 10−4), respectively (Table 1). The Kd values of Gal-3 calculated from these Ka values are 0.067 μM for hTg, 0.091 μM for bTg, and 0.24 μM for pTg. The affinities (Ka or Kd) of Gal-3 for hTg, bTg, and pTg determined by ITC agree with those obtained with hemagglutination inhibition experiments. Gal-3 was able to interact with hTg, bTg, and pTg, despite the presence of terminal sialic acids on their N-linked complex-type glycans because Gal-3 is capable of binding to internal LacNAc residues that contain terminal sialic acids. The complex-type glycans of pTg contain fucose residues attached to the core GlcNAc and sulfate groups on the LacNAc residues (28,29). Such modifications in the glycan structures possibly contribute to the relatively lower affinity (Ka) of hGal-3 for pTg (compared with hTg and bTg; Table 1). The iodination levels of bTg and pTg are comparable, but their affinities (Ka; Table 1) for Gal-3 are different. Therefore, it seems that the affinities of Tgs are not correlated with their iodine levels. Gal-3 shows enhanced affinity for poly-LacNAc chains compared with LacNAc (24,25). Tg produced by transformed thyroid cells contains poly-LacNAc units in their complex-type glycans (30). Therefore, it is likely that Gal-3 will bind to Tg produced by transformed cells with even higher affinity.
The valence of hTg for hGal-3, as calculated from the n values (valence = 1/n) (27) obtained through ITC experiments (Table 1), is 14. Therefore, the stoichiometry of hTg:Gal-3 is 1:14 (1 molecule of hTg binds to 14 molecules of hGal-3). ITC-driven stoichiometry is consistent with the value determined by quantitative precipitation assays (shown below).
Binding of Gal-3 to Tg leads to complex formation at stoichiometric concentration
Quantitative precipitation assays showed that binding of Gal-3 to Tg resulted in insoluble complex formation at stoichiometric concentration. This complex formation is glycan mediated, as the complex was dissolved by lactose. Quantitative precipitation assays were done by adding varying concentrations of hTg to a fixed amount to hGal-3 (Fig. 2A). Alternately, varying concentrations of hGal-3 were added to a constant amount of hTg (Fig. 2B). In both cases, robust precipitation curves were observed with the same stoichiometry at the peak (hTg:hGal-3 = 1:14). As evident from Figure 2, the amount of complex formation is very sensitive to the relative ratio of hTg and hGal-3. To investigate further how the relative concentrations of hTg and hGal-3 regulate the precipitation process, the kinetics of hTg/hGal-3 complex formation were measured at different hTg concentrations. At the stoichiometric concentration (Tg:Gal-3 = 1:14), the rate and amount of precipitation were the highest (Fig. 2C). The kinetics of complex formation was considerably fast. Most (88%) of the total precipitation was formed within three minutes, and the precipitation maxima were reached within 12 minutes. When hTg concentrations were lower or higher than the stoichiometric concentration, the amount and rate of complex formation were significantly lower (Fig. 2C). For example, at 3.4 μM Tg concentration, the amount of complexes was reduced by 72% compared with the amount obtained with 1 μM of hTg. This result indicates that if the proper ratio between hTg and hGal-3 concentrations is not achieved, the rate and amount of complex formation will be insignificant, even at a higher concentration of hTg or hGal-3. A similar trend of stoichiometry-dependent complex formation was observed with bTg and hGal-3 (Fig. 2D). Such complex formation between Tg and Gal-3 reduces the concentration of freely available Tg and Gal-3. The potential biological significance of this result is described below in a later section.
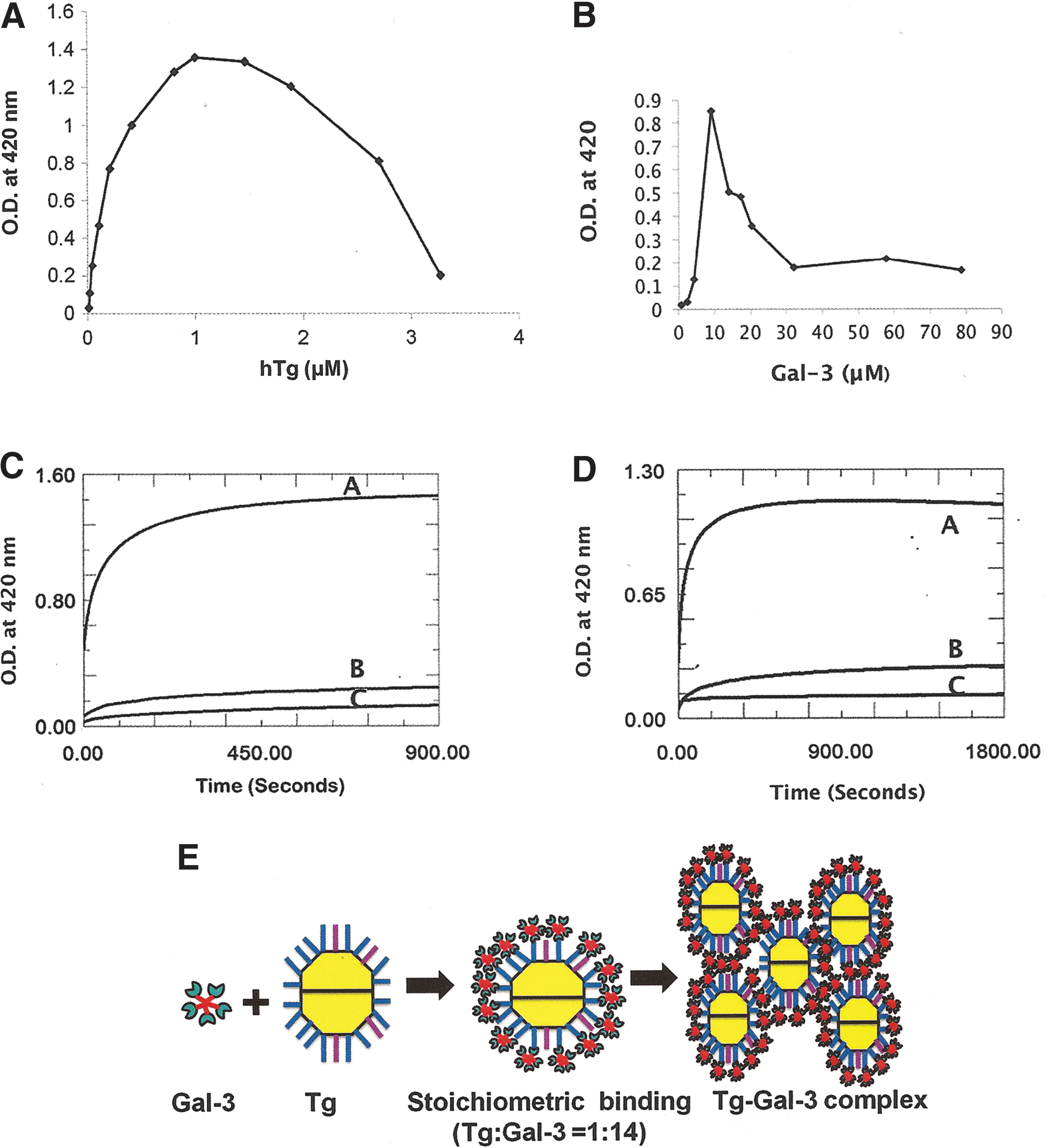
Complex formation between hTg and hGal-3.
Addition of free Tg dissolves insoluble Tg–Gal-3 complex
When free hTg solution was added to preformed hTg-hGal-3 complex obtained at their proper stoichiometric concentrations, the complex dissolved. The process was found to be dependent on the concentration of free hTg added to the complex (Fig. 3A). The addition of 0.67 μM, 1.8 μM, and 2.9 μM hTg dissolved 14%, 74%, and 91%, respectively, of preformed hTg-hGal-3 insoluble complex. Although the dissolution process is concentration dependent, the relationship between the concentration of added free hTg and the percent of dissolved complex was found to be nonlinear (Fig. 3B). For example, when the free hTg concentration was increased by 2.7 times (from 0.67 to 1.8 μM), the amount of dissolved complex increased by 5.3-fold (14–74%). When the hTg concentration was increased by 4.3-fold (from 0.67 to 2.9 μM), the amount of dissolved complex increased by 6.5-fold (14–91%; Fig. 3B). The insoluble hTg-hGal-3 complex dissolved rather rapidly upon the addition of free hTg. Depending on the concentration of added free Tg, up to 91% of the complexes dissolved in less than two minutes. Interestingly, the addition of free Gal-3 was also found to dissolve preformed hTg-hGal-3 complex (Fig. 3C). However, the ability of free hGal-3 to dissolve preformed hTg-hGal-3 complex was less than that of free hTg. It took a much higher concentration of free hGal-3 (41 μM of free hGal-3 compared with 0.67–2.9 μM of free hTg) to dissolve only 25% of preformed hTg-hGal-3 complex, while 2.9 μM of free hTg dissolved 91% of the complex.
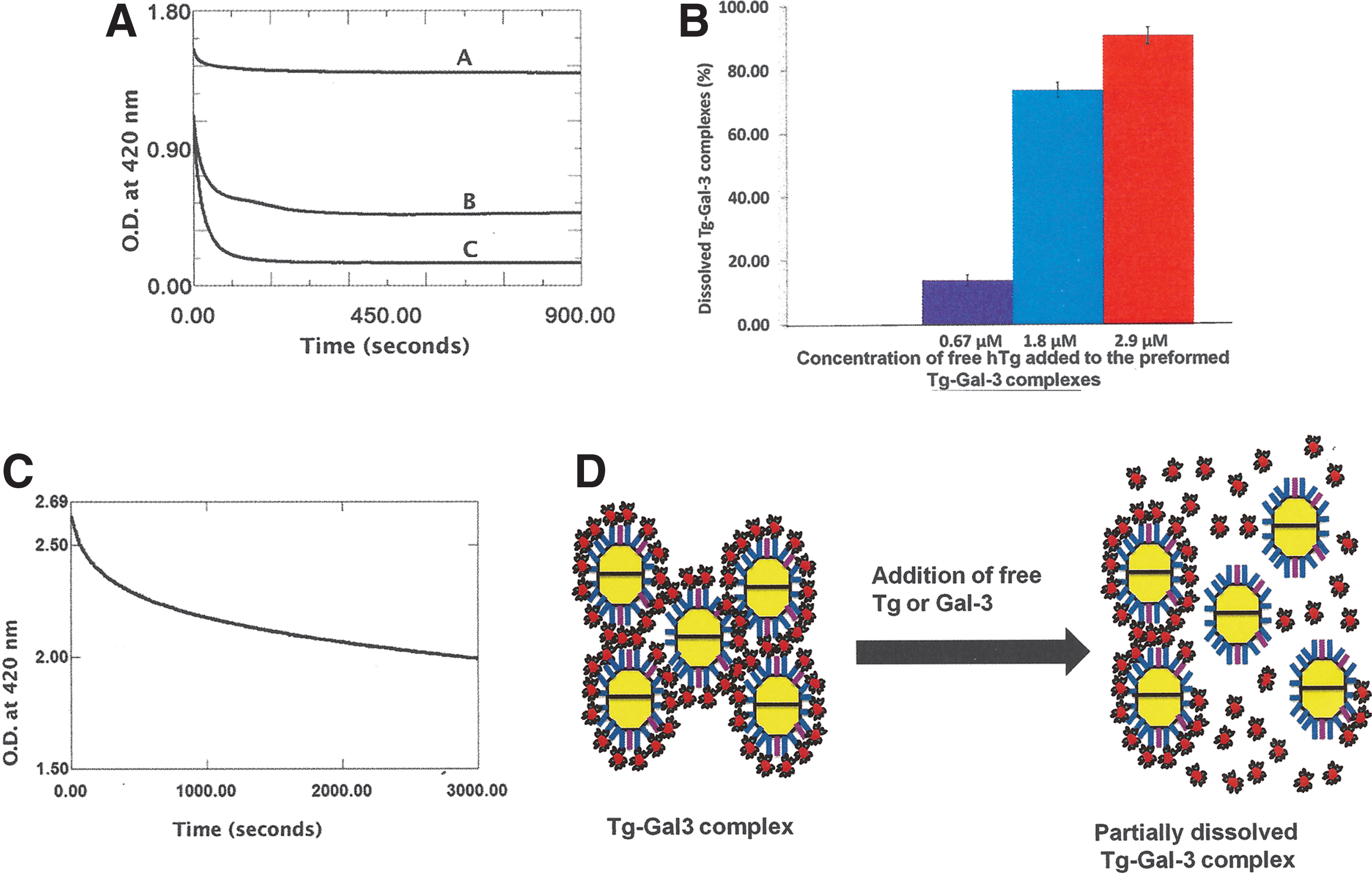
Free hTg dissolves preformed hGal-3–hTg complexes in a concentration-dependent manner.
Discussion
Interaction between Gal-3 and Tg may influence their detection in clinical samples
Despite current technical limitations, serum Tg measurement is the cornerstone of long-term monitoring for most thyroid cancer patients. For the diagnostic interpretation of serum and tissue Tg levels, it is important to understand the pathophysiology of Tg secretion and the limitations of Tg detection methods (31). The accuracy of the Tg test is compromised by several factors, including “hook” effects (when measuring high concentrations) and Tg autoantibody (TgAb) interference (32). Being one of the main thyroid autoantigens, Tg elicits the generation of TgAb (33). Binding of TgAb to Tg (known as TgAb interference) can produce false-positive or false-negative results and thus can potentially mask the presence of a recurrent or persistent thyroid carcinoma (34). Because of its high affinity binding to Tg, Gal-3 may cause a similar interference problem. It seems imperative that sera sent for Tg estimation be checked for TgAb by sensitive immunoassays (32). A similar approach should be taken to detect any Gal-3 that may cause possible interference in the detection of Tg. At present, it is difficult to identify which TgAb-positive sera suffer from interference. Therefore, the detected level of Tg of TgAb-positive sera should be interpreted with caution (32). A similar precaution should be taken for Gal-3 positive sera. As mentioned before, Gal-3 itself is a clinical biomarker of thyroid cancer. Formation of Tg–Gal-3 complex will reduce the amount of measurable free Gal-3 in the clinical samples and will introduce errors in the detection assays of Gal-3. Both FNA (18) and serum (19 –21) samples of thyroid cancer patients have been examined for the presence of Gal-3. Gal-3 levels in nonserum samples differ significantly between thyroid cancer patients and healthy subjects. However, such differences in Gal-3 levels in serum samples are not apparent (19 –21). Besides being expressed by malignant thyroid tissue, human Gal-3 is produced by various other normal tissues in different locations. It is possible that Gal-3 from other tissues contributes to the serum Gal-3 level as well.
Gal-3 may influence multisite immunometric assay
Immunometric assay (IMA) typically underestimates the Tg concentration as a result of TgAb interference (4). To overcome this problem, multisite IMA has been developed that uses selected monoclonal antibodies (mAbs) that are specific for Tg epitope domain not involved in thyroid autoimmunity. In such selective epitope-targeting approach, the binding of mAbs to Tg will be inhibited by steric problems caused by TgAb that are already complexed with Tg. Galectin-3 bound to Tg may cause similar steric problems for mAbs and adversely affect the assays.
Any physiological or functional changes that alter Tg concentration or any structural alteration that blocks TgAb binding epitopes of Tg may potentially decrease the effectiveness of immunohistochemical staining with anti-Tg antibodies (30). Antibody binding sites may be sterically blocked by bound Gal-3 molecules on Tg. This, in turn, can interfere with immunohistochemical analysis.
Stoichiometric binding of Gal-3 and Tg leads to the formation of insoluble cross-linked complexes. However, when the binding is not stoichiometric, soluble and insoluble complexes of different sizes would still form between Gal-3 and Tg. In this situation, bound Gal-3 molecules on Tg can potentially interfere with other binding activities of Tg, including its interaction with TgAb.
Complex formation between hGal-3 and hTg may enhance their metabolic clearance
In vivo Tg concentration is influenced by (i) the rate of secretion of Tg from the tumor, (ii) the affinity of the TgAbs present, and (iii) the rate of complex formation between Tg and TgAb and the subsequent clearance of the complex (32). The half-life of free Tg is approximately three days, and it is suggested that the metabolic clearance of Tg–TgAb complexes may be faster compared with the metabolic clearance of free Tg (4). Similarly, Tg–Gal-3 complexes may potentially be metabolized faster compared with free Tg. While it takes around 24 hours to form Tg–TgAb complexes (4), Tg–Gal-3 complexes form in minutes (as shown by the current data). Therefore, the formation and subsequent metabolic clearance of Tg–TgAb complexes will be significantly impacted by the rapid complex formation between Tg and Gal-3. While the addition of free Tg as well as free Gal-3 dissolves Tg–Gal-3 complexes (as shown here), it is not apparent from the available literature if additional free TgAb is capable of breaking Tg–TgAb complexes. Gal-3 and TgAb bind to different epitopes on Tg. Gal-3 interacts with the complex-type glycans of Tg, while TgAb recognizes the protein backbone. It will be interesting to investigate how Gal-3 and TgAb compete for Tg and how the interaction between these three entities regulates their respective biological functions.
The data obtained in the present study support a model of Gal-3-mediated homeostasis of Tg (Fig. 4). Similar to Tg, biomarkers of many other cancers are glycoproteins (15,35 –38) that are potential ligands of Gal-3. Most of these cancer cells also express Gal-3 (15,39,40). Interestingly, Gal-3 is used as a biomarker of certain other cancers other than thyroid carcinomas (13,14,41). It is therefore tempting to speculate that Gal-3 could potentially bind and reversibly precipitate these glycoprotein biomarkers (as shown with Tg in the present study) and influence their clinical detection, homeostasis, and biological functions.
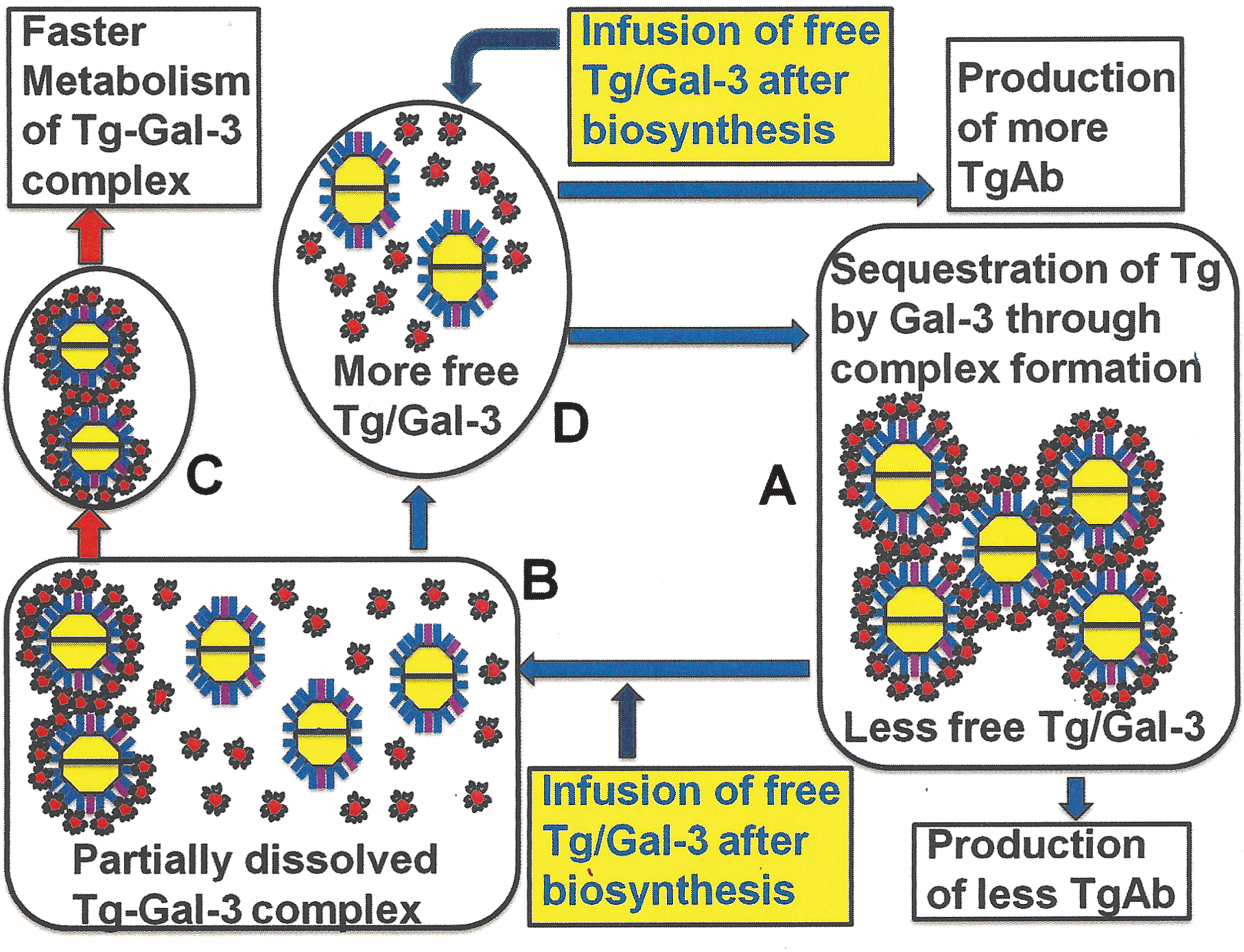
Proposed glycan-mediated biomarker homeostasis process. hGal-3 binds to hTg with high affinity (as shown in Fig. 1). Interaction between hGal-3 and hTg under proper stoichiometric conditions leads to complex formation
Footnotes
Acknowledgments
This work was supported by start-up funds (T.K.D., P.B.) and by Research Excellence Fund (T.K.D.) provided by Michigan Technological University.
Author Disclosure Statement
No competing financial interests exist.