
Abstract
Background:
In humans, the cause of arrested migration of the median thyroid anlage resulting in an ectopic sublingual gland is unknown. These ectopic glands have a normal follicular architecture but their thyrotropin-induced growth is insufficient, leading to congenital hypothyroidism in the vast majority of affected subjects. We hypothesized that arrested migration is due to premature differentiation [reflected by decreased telomere length (TL)], as observed in neural tube defects in mice.
Methods:
Absolute TL and telomerase reverse transcriptase (hTERT) expression was measured in four ectopic and six orthotopic thyroids. TL was measured by quantitative polymerase chain reaction of genomic DNA, whereas hTERT expression was measured by quantitative polymerase chain reaction of total RNA.
Results:
The mean±standard deviation TL (in kilobases per diploid genome) was 140.45±40.07 in ectopic and 97.50±30.48 in orthotopic thyroids (p=0.12). Expression of hTERT was quiescent in both ectopic and orthotopic thyroids.
Conclusions:
Compared with orthotopic thyroids, TL shortening is not observed in ectopic thyroid tissues and, consequently, no compensatory hTERT expression was measured. This makes premature differentiation an unlikely cause of arrested migration and it suggests, indirectly, that ectopic thyroids are not at higher risk of cancer than orthotopic thyroids.
Introduction
C
The molecular causes of CH due to TD (CHTD) remain unclear. Germline mutation(s) in transcription factors implicated in the morphogenesis of the thyroid gland (NKX2.1, PAX8, FOXE1, and GLIS3) have been found in a small percentage of patients with CHTD and are mostly associated with other malformations (5 –9). Furthermore, linkage studies have excluded these genes in some multiplex families with CHTD (10). This low prevalence of germline mutations, together with the high discordance rate (92%) between monozygotic (MZ) twins (11), underline the importance of looking for somatic alterations in the dysgenetic thyroid tissue itself.
Telomeres are specialized functional complexes that protect the ends of eukaryotic chromosomes from chromosomal fusion; hence, telomeres maintain genomic stability and cell replicative capacity (12). In somatic cells, telomere length is primarily maintained by telomerase, a ribonucleoprotein consisting of telomerase reverse transcriptase (TERT) and telomerase RNA (13). Immature undifferentiated cells show telomerase activity, which is lost upon differentiation into a mature specialized cell type (14). TERT overexpression extends the proliferative life span of normal human cells (15). Conversely, telomerase inhibition promotes an initial differentiation step of primate embryonic stem cells (16). Mice genetically deficient for the telomerase RNA (mTR−/−) present with telomeres shortening and neural tube defects (17). Cells derived from these mice show increased apoptosis and decreased viability (17). These observations support the concept that decreased telomere length (TL) below a given threshold might impair cell viability during organogenesis. TERT remains active in proliferating spermatocytes, reflecting the importance of maintaining telomere length in the germline (18). In normally differentiating tissues, it decreases rapidly after a peak at the blastocyst stage (19) to become quiescent in most healthy noncancerous somatic tissues. Consequently, TL is at its maximum early in life and shows a progressive decrease with ageing in human somatic tissues (20). Only cancerous cells express TERT to reestablish a TL sufficient to escape senescence and apoptosis (21).
Ectopic thyroids are well-differentiated tissues with normal follicular architecture (4,22) and express all key enzymes required for thyroid hormonogenesis (23). Therefore, the hypothyroidism of patients with ectopic thyroids is likely due to limited thyrotropin-stimulated growth of the smaller amount of tissue present (4). We hypothesized that accelerated differentiation could induce a premature arrest in migration and that this would be reflected in shorter telomere length (TL). We further investigated whether a compensatory increase of TERT expression is observed in ectopic thyroids, as in cancerous tissues which thereby escape senescence (21).
Patients and Methods
Patients and tissue samples
We obtained flash-frozen samples of ectopic thyroid tissue removed from four girls (aged 8, 10, 14, and 15 years) to alleviate local symptoms (i.e., dysphagia) or for cosmetic reasons. For controls, we used healthy thyroid tissue from three girls (aged 8, 12, and 16 years) and three boys (aged 5, 5, and 9 years) who were either operated on prophylactically (four RET-mutated individuals of known MEN2 families) or for papillary thyroid cancer (Table 1). This study was approved by the Sainte Justine Ethics committee.
CH, congenital hypothyroidism due to thyroid ectopy; MTC, medullary thyroid cancer: healthy thyroid tissue obtained from prophylactic thyroidectomy in RET–positive patients; PTC, papillary thyroid cancer: healthy contralateral thyroid tissue obtained from near total thyroidectomy.
F, female; M, male.
TL (kb), telomere length in kilobases per diploid genome.
RQ, relative expression of human telomerase reverse transcriptase (hTERT). The hTERT expression in HCT116 colon cells serves as reference (set point of 1.0).
DNA and RNA extraction from tissue samples
DNA was extracted using a pureLink genomic DNA Mini kit (Life Technologies). RNA was extracted using a miniRNeasy kit (Qiagen). RNA from HCT116 cells (human colon cancer cells) was used as positive control for human TERT (hTERT) transcript quantification.
Measurement of TL by quantitative polymerase chain reaction
Quantitative polymerase chain reaction (qPCR) was performed with 30 ng/μL genomic DNA using the ViiA 7 Real-Time PCR System (Life Technologies) as previously described (24 –26). Briefly, telomere repeat copy number is determined relative to a single-copy number in all samples using standard curves. This ratio is proportional to the telomere length. The 36B4 gene, encoding acidic ribosomal phosphoprotein P0, has been used as a reference single-copy gene. Primers, conditions, and standard curves are provided in references (24 –26).
Measurement of hTERT mRNA by reverse transcription quantitative PCR
Total RNA was treated with DNAse then reverse transcribed with random primers using the Maxima First Strand cDNA synthesis (Thermo Fisher), as described by the manufacturer. Before use, RT samples were diluted 1:5. Gene expression was determined using assays designed with the Universal Probe Library from Roche (
Statistical analysis
The data were first analyzed for normal distribution (Kolmogorov-Smirnov test) and homogeneity of variances (Bartlett's test), and group means were then analyzed using bilateral two sample t-test with Welch correction. Statistical analyses and graphs were performed using the software package R 3.1.2 (R Core Team) and the GraphPad Prism version 6.00 (GraphPad Software). Post hoc power analysis was performed with the G power software (27). To allow for more detection of differences with a reasonable power, differences were considered significant at p<0.1.
Results
TL is not shorter in ectopic thyroids
The mean TL (mean kilobases per diploid genome±standard deviation) of ectopic thyroids was 140.45±40.07, actually even longer than that of orthotopic thyroids (97.50±30.48), although the difference did not reach significance (p=0.12; [95%CI −16.7 to 102.6]; two-tailed Welch's t-test) (Table 1 and Fig. 1). Given the hypothesis that TL is shorter in ectopic thyroids when compared with orthotopic thyroid, the analysis using one-tailed two sample t-test with Welch correction showed a consistent nonsignificant result (p=0.98; [95%CI −89 to∞]). A linear model showed no significant effect of age on TL for all samples combined, and no significant interactions of age, sex, and diagnosis (ectopic thyroids vs. orthotopic controls) were observed.
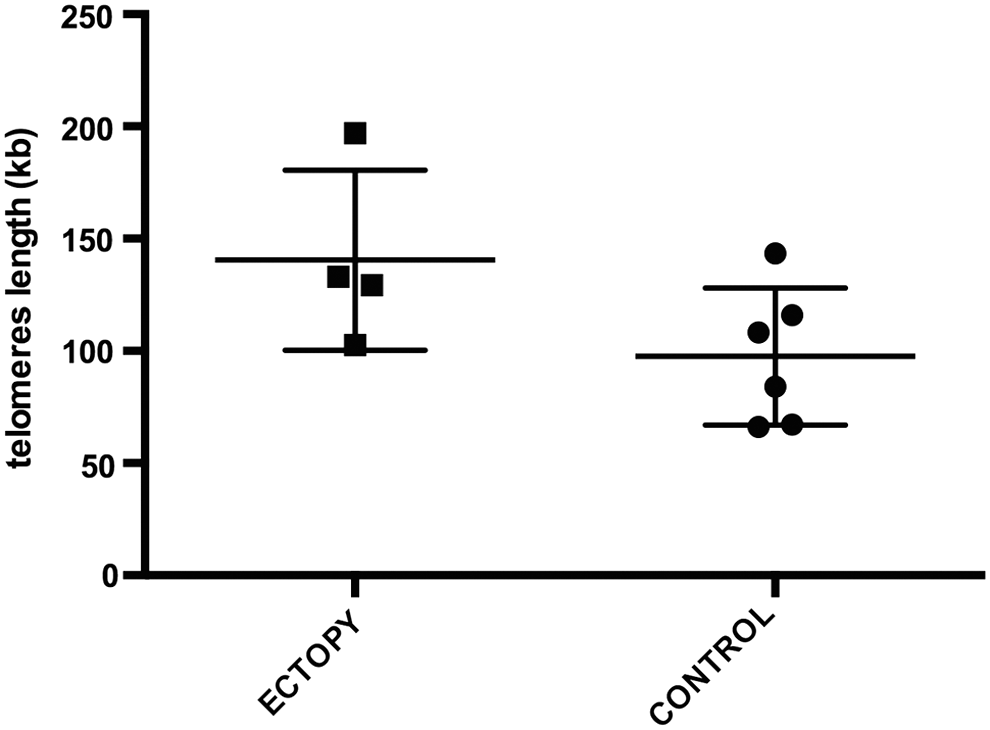
Telomere length (TL) in orthotopic and ectopic thyroid tissues. TL (reported in kilobases per diploid genome on the y-axis) in four ectopic thyroids (left) represented by a full square and in six orthotopic thyroids as control (right) represented by a full circle. The bar in the middle represents the mean, and the two adjacent upper and lower bars depict the standard deviations. The mean TL (mean±standard deviation) of ectopic thyroids was 140.45±40.07, actually longer than that of orthotopic thyroids (97.50±30.48), although the difference did not reach significance (p=0.12 [95%CI −16.7 to 102.6]; two-tailed Welch's t-test).
Transcription of hTERT is absent in ectopic and orthotopic thyroids
Human TERT transcripts were not detectable in ectopic and orthotopic thyroids using standard (Fig. 2) and quantitative RT-PCR methods (Table 1; all RQs are less than 0.025), but hTERT transcripts were clearly detectable in HCT116 colon cancer cells (positive controls, Fig. 2) and served as reference (value of 1) for the RQ calculation (Table 1).
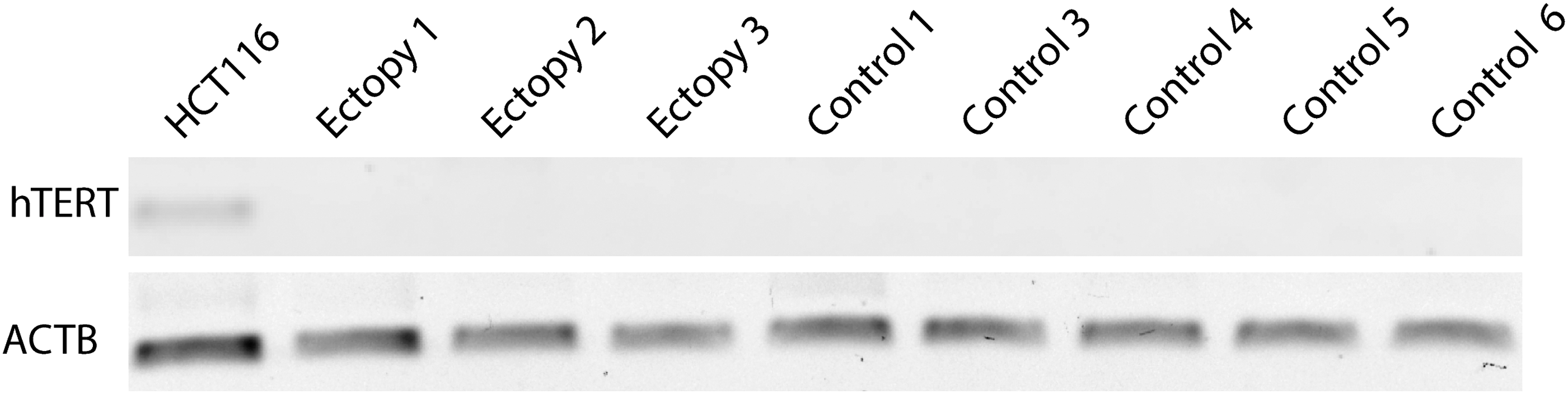
Lack of human telomerase reverse transcriptase (hTERT) transcripts in orthotopic and ectopic thyroid tissues. Absent signal of hTERT transcripts with the standard semiquantitative RT-PCR method (1.5% agarose gel, upper panel) are consistent with the results of the quantitative RT-PCR method, which are expressed in relative expression (RQ) to the control (in HCT116 colon cells); RQ values are listed in the last column of Table 1. ACTB, actin beta.
Discussion
Telomere shortening can lead to dysfunctional telomeres, chromosomal instability and apoptosis (28). Apoptosis is a physiological process that starts early during development. In animal models, congenital neural tube defects are linked to reduced telomerase activity and decreased TL (17). TL is tissue-specific even if a high correlation exists between tissues of individuals matched for age and shows a continuous decrease with age (20). A marked synchrony exists in average TL between embryonic human tissues, but this synchrony is lost during extrauterine life, probably as a result of different proliferative programs (28,29). Therefore, to assess the biologically plausible association between reduced telomere length (TL) and congenital malformations, we used ectopic and orthotopic thyroids obtained from children of similar age.
Herein, we show that in both ectopic and orthotopic control thyroids (i) TL is similar and even tends to be longer in ectopic thyroids, and (ii) hTERT expression is quiescent. Of note, all control tissues were healthy thyroid tissue (prophylactic thyroidectomy for MTC and healthy contralateral tissue of unilateral PTC). These findings extend those of our previous study (23) and are consistent with the histological aspect of ectopic thyroids, which shows a well-differentiated tissue with normal follicular architecture (4). Telomerase inhibition promotes the initial steps of cell differentiation of primate embryonic stem cells (16) and, conversely, the induced differentiation of immortal cells inhibits telomerase activity (14). Consequently, quiescent hTERT expression is expected in differentiated non-proliferative tissue. Specifically, in papillary thyroid cancer, hTERT is expressed (30) and TL is decreased. These results suggest that ectopic thyroids are not more prone to malignant transformation compared with orthotopic thyroids, but the following points hamper any firm conclusion: (i) there is an obvious report bias of cancers in ectopic thyroids because of their “curiosity value” (with the risk of overestimating the risk) (31); (ii) the majority of reports refers to ectopic thyroid tissues in the thyroglossal duct with the presence of normal thyroid gland in situ (32), which is different from thyroid cancer arising in a congenital ectopic thyroid; and unfortunately, (iii)the recent US epidemiological survey of thyroid cancer (33) does not contain information about the anatomic location of the cancerous glands (Louise Davies, personal communication).
TL is influenced by environmental cues (34,35) and a conserved TL is also an additional argument against environmental causes of CHTD, consistent with the lack of a seasonal impact on the occurrence of ectopic thyroids (1). TL in blood DNA is more similar in MZ than in DZ twins (36), which is a further argument for a somatic event accounting for discordance of MZ twins for CHTD (11).
This study has some limitations. First, TL and hTERT expression profiles in tissue collected and analyzed postnatally may not reflect the dynamics of embryonic expression. Second, the sample number is small, but this is inherent to the exceedingly exceptional indication for surgically removing ectopic and orthotopic thyroid tissue in children. Therefore, our study does not have enough power to reach a firm conclusion, and the fact that zero is contained in the wide 95%CI of the difference between the means indicates that we cannot rule out the possibility that there is no difference (37), but we can at least conclude that it would be unlikely that TL in ectopic thyroids is shorter than in orthotopic thyroids.
Conclusion
TL shortening—the hallmark of accelerated cell senescence and differentiation—is not observed in ectopic compared with orthotopic thyroid, and hTERT is not expressed in either tissue. Ectopic thyroids have a molecular profile comparable to that of normal non-tumoral thyroids (38), which differs from that seen in thyroid cancer. Our findings suggest that TL shortening is unlikely to cause arrested migration and, indirectly, that ectopic thyroids are not at higher risk for thyroid cancer than orthotopic thyroids (30,39).
Footnotes
Acknowledgments
We thank the patients and their parents for their cooperation. We thank Raphaëlle Lambert (Institut de Recherche en Immunologie et Cancérologie, Université de Montréal) for technical assistance.
This work was supported by a grant from the Canadian Institutes of Health Research (CIHR-MOP-130390 to J.D.); Research in pediatric thyroid diseases at Centre Hospitalier Universitaire (CHU) Sainte-Justine is supported by the Girafonds/Fondation du CHU Sainte-Justine (to J.D. and G.V.V.).
Author Disclosure Statement
No competing financial interests exist.