
Abstract
Background:
Both perinatal hypothyroxinemia and perinatal iron deficiency (ID) are associated with poor neurodevelopment in offspring. Iron is an important component of thyroid peroxidase, a key enzyme in the synthesis of thyroid hormone. The authors’ previous study demonstrated that perinatal ID can lead to maternal hypothyroxinemia during pregnancy. The goal of this study was to determine whether perinatal ID-associated hypothyroxinemia can cause brain defects prior to neonatal brain iron depletion.
Methods:
Two rat models were established to imitate the two common types of maternal ID (mild ID with anemia [ID + A] and ID without anemia [ID – A]), and iron limitation was initiated two weeks before pregnancy. Maternal and neonatal thyroid hormones in serum were analyzed at postnatal day (P) 0 and P10. Neonatal thyroid hormone, as well as mRNA expression of some thyroid hormone–responsive genes in the cerebral cortex and hippocampus, were measured at P10. Serum iron and brain iron concentrations were analyzed by inductively coupled plasma mass spectrometry. Liver iron concentration was determined using graphite furnace atomic absorption spectroscopy. Hemoglobin was analyzed with an automated blood coagulation analyzer. Surface righting reflex and vibrissae-evoked forelimb placing were measured to assess the sensorimotor behaviors.
Results:
It was found that pre-pregnant mild ID resulted in maternal hypothyroxinemia, which lasted from gestation day 13 to P10. Pre-pregnant mild ID decreased the neonatal brain total triiodothyronine level at P10. Consistent with a low total triiodothyronine level, the mRNA expression of some thyroid hormone–responsive genes (Mbp, RC3, and Srg1) were significantly reduced in the neonatal cerebral cortex and hippocampus in both ID rat models at P10. Furthermore, ID rat pups at P10 showed retarded sensorimotor skills. No significant difference was found between the control and the ID pups in terms of iron concentrations in the neonatal brain at P10.
Conclusions:
This study demonstrates that perinatal ID-associated hypothyroxinemia is sufficient to impair early brain development, regardless of whether the neonatal brain iron level is normal, and monitoring thyroid hormone level is indicated in ID pregnant women.
Introduction
B
ID is the single most common micronutrient deficiency worldwide, estimated to affect one to two billion people. Pregnant women and children are most vulnerable to developing ID (11). Perinatal ID can result in cognitive and behavioral function deficits in offspring (3). Its mechanisms are currently considered to be associated with the impairment of energy metabolism, neurotransmitter synthesis, and myelination, which are highly dependent on iron-containing enzymes and hemoproteins (12 –17). A previous study by Beard et al. reported that neurodevelopmental delays occurred prior to brain iron depletion in postnatal day (P) 10 rat pups exposed to moderate perinatal ID (18), suggesting that an unknown mechanism contributed to the behavioral changes prior to brain ID. Thyroid peroxidase (TPO) is an iron-containing enzyme that is essential for the synthesis of thyroid hormones (19). The authors’ previous study demonstrated that perinatal ID can cause a decrease in TPO activity, which leads to maternal hypothyroxinemia during pregnancy (20). Several recent studies have reported that perinatal ID can induce low neonatal brain thyroid hormone levels and thereby alter the expression of thyroid hormone responsive genes (21 –23). Based on these reports, it was hypothesized that perinatal ID-induced maternal and neonatal hypothyroxinemia could be an early mechanism of impairing brain development prior to neonatal brain iron depletion.
The present study established two rat models with different degrees of ID. Neonatal brain iron levels, iron-responsive genes, thyroid hormone levels, thyroid hormone–responsive brain genes, and cognitive and neurobehavioral performance were analyzed. This is the first report to show that perinatal ID-induced hypothyroxinemia is sufficient to impair early brain development, regardless of whether the neonatal brain iron level is normal. It is speculated that hypothyroxinemia might be an early event and reduced brain iron levels might be a later event in the impairment of brain development caused by perinatal ID.
Materials and Methods
Diets and animals
Female Sprague–Dawley rats (n = 120), weighing approximately 200 g, were obtained from the Laboratory Animal Facility (Shanghai, China). They were randomly assigned to one of three groups according to the different iron concentrations in the diet, as previously described (20): control, ID with anemia (ID + A), and ID without anemia (ID – A). Iron concentrations of 10, 30, and 70 ppm were given to rats in the ID + A, ID – A, and control groups, respectively. They were fed diets with different iron content for two weeks before mating until they were euthanized at gestational days (G) 0 and G13, and at P0 and P10. The diets were all purified diets that are a modification of the AIN-93G diet by adding ferrous sulfate heptahydrate in place of ferric citrate (24). Considering the effects of other known micronutrients on thyroid function, iodine, selenium, copper, and zinc content were measured by inductively coupled plasma mass spectrometry (ICP-MS; Series 7500A, Agilent Technologies Inc., Santa Clara, CA), and no difference in the content of any of these elements was found between the sets of diets, as previously described (20). All litters were culled to 10 pups per litter by P2. Animals were housed in a temperature-controlled animal facility with a reversed 12/12 h light/dark cycle. The room temperature was maintained at 24°C, and relative humidity was approximately 45%. Diets and demineralized water were administered ad libitum for all groups. All procedures were approved by the ethics committee of the China Medical University.
Tissue collection
The day on which a vaginal plug is noted is defined as G0. Dams were weighed and then euthanized with intraperitoneal chloral hydrate on G0, G13, P0, and P10. Blood samples (approximately 14 mL) were obtained from the heart for the measurement of hemoglobin, serum ferritin, and thyroid hormone levels. P0 pups were decapitated, and the blood was collected. Each blood sample was from seven to eight hearts from neonatal rats from each dam, and the blood sample was from six dams in each group for the measurement of hemoglobin and thyroid hormones. For P10 pups, each blood sample was from five to six hearts from neonatal rats from each dam, and the blood sample was from six dams in each group for the measurement of hemoglobin and thyroid hormones. The pups were perfused with physiological saline through the left ventricle. A small portion of liver (0.1–0.2 g) was removed, washed, weighed, and digested in 5 mL nitric acid for 24 h. At P10, neonatal brains were perfused with physiological saline through the left ventricle and removed. Next, some of the hippocampus and cerebral cortex were transferred to a 4% paraformaldehyde solution for 24–48 h for histochemical iron detection and Nissl staining. Some of the hippocampus and cerebral cortex were dissected on ice, and were fresh frozen in liquid nitrogen for tissue thyroid hormone measurement and polymerase chain reaction (PCR) analysis. The others were rinsed, weighed, and digested in 5 mL nitric acid for iron analysis.
Analyses of iron parameters
Whole blood was collected into anticoagulation and coagulation tubes. Hemoglobin was immediately analyzed with an automated blood coagulation analyzer (K-4500; Sysmex, Kobe, Japan). Blood samples in coagulation tubes were centrifuged at 2580 g for 10 min at room temperature, and the serum was stored at −70°C.
Serum ferritin concentration was determined in duplicate using a rat ferritin enzyme-linked immunosorbent assay kit (RSCPRA011R; BioVendor Laboratory Medicine, Inc., Heidelberg, Germany). The assay was performed as per the kit instructions. Serum was diluted to 1:50 with sample diluents included in the kit. Sera were analyzed in duplicate. The absorbance was measured at 450 nm using a microtiter plate reader (Infinite F200 PRO; Tecan, Männedorf, Switzerland).
Serum iron was analyzed using an internally standardized method by ICP-MS (Series 7700; Agilent Technologies, Inc.). An aliquot of 200 μL of serum was added to 1.8 mL of diluents containing Triton X-100 (0.05%) and nitric acid (0.5%). Next, the aliquot was vortex-mixed with 20 μL of diluents (5%) of an internal standard mix (no. 5188-6525; Agilent Technologies, Inc.) for detection.
The wet digestion method was used to digest the iron solution containing liver and brain tissues, as described by Prohaska (25). Liver iron concentration was determined using graphite furnace atomic absorption spectroscopy (GF-AAS; 180-80; Hitachi High-Technologies Co., Tokyo, Japan), and brain iron concentration was determined using ICP-MS (Series 7700; Agilent Technologies, Inc.). When wet digestion of liver tissue was complete, the remaining liquid was transferred into a 20 mL volumetric flask and diluted with deionized water to 15 mL. It was then mixed and analyzed using GF-AAS. When wet digestion of brain tissue was complete, the remaining liquid was transferred into a 15 mL volumetric flask and diluted with deionized water to 5 mL. It was then mixed and analyzed with ICP-MS.
Serum and brain thyroid hormone measurements
A chemiluminescent immunoassay (Immulite 1000; Diagnostic Products Co., Los Angeles, CA) was used for analyzing serum total thyroxine (TT4) and total triiodothyronine (TT3). The thyrotropin (TSH) level was measured with a rat kit (cat no. CEA463Ra; Cloud-Clone Corp., Houston, TX), which is based on a competitive inhibition enzyme immunoassay technique.
Thyroid hormones were extracted from half brains using a modification of the method described by Morreale de Escobar et al. (21,26). Frozen hippocampus and cerebral cortex were homogenized in 4 mL of methanol (containing 1 mM of PTU)/g tissue with steel balls using a homogenizer (Retsch TissueLyser II; Qiagen, Hilden, Germany) at 25 Hz for 8 min, as described previously. The homogenate was centrifuged at 2000 g at 4°C for 15 min, and the supernatants were analyzed via a chemiluminescent immunoassay, as described above.
TPO activity
TPO activity analysis was conducted with a modified mini-assay method, as previously described by Hess et al. (20).
Nissl staining
After deparaffinization in xylene and rehydration, sections were incubated in toluidine blue (1%) for 10 min. Then sections were washed with distilled water, incubated in ethanol (95% for 30 sec and 100% for 1 min) and xylene for 2 min, and mounted with neutral gum.
Quantitative real-time PCR analysis of gene expression in hippocampus and cerebral cortex
Total RNA was extracted from the hippocampus and cerebral cortex using Trizol reagent (15596-026; Life Technologies, Carlsbad, CA) following the manufacturer's instructions. cDNA was synthesized from 2 μg of total RNA using a PrimeScript RT reagent kit (DRR037A; TaKaRa Bio, Inc., Otsu, Japan). Quantitative real-time PCR (qPCR) was performed with the SYBR Premix Ex Taq kit (RR420A; TaKaRa) and a LightCycler 480 Instrument (Roche Applied Science, Penzberg, Germany). PCR reactions were performed with cDNA in a final volume of 20 μL, according to the manufacturer's protocol. The data were normalized to the HPRT mRNA level and expressed as arbitrary units. The primers used in real-time PCR reactions are listed in Supplementary Table S1 (Supplementary Data are available online at
Behavioral assessments
Thyroid hormone controls a wide range of developmental processes, including cognitive and sensory-motor behaviors, such as learning and memory, auditory and visual attention, fine and gross motor skills, as well as the integration between the hemisphere. Two tests were selected from the battery described by Altman and Sudarshan (27) in order to assess the sensorimotor behaviors: (i) surface-righting reflex and (ii) vibrissae-evoked forelimb placing.
Surface-righting reflex
Pups were individually placed in a supine position on a flat surface, and the time elapsed to righting (12,28) was measured. The time that pups returned to prone posture with all four paws on the surface was recorded at P5, P7, and P10. Animals were tested for a maximum of 2 sec.
Vibrissae-evoked forelimb placing
The rat was held by its torso, allowing one forelimb to hang free. The vibrissae on the ipsilateral side were brushed against the table edge. A successful response was determined by the immediate placing of the ipsilateral forelimb on the table surface (29,30). The number of ipsilateral forelimb placements out of 10 trials per side was averaged for each animal. At P5, P7, and P10, the outcome measure was percent vibrissae-evoked forelimb placing by litter (30).
Statistical analysis
All data were statistically analyzed using SPSS Statistics for Windows v17.0 (SPSS, Inc., Chicago, IL). Tests of normality and homogeneity of variance were carried out for data regarding liver iron, serum ferritin, serum iron, hemoglobin, serum TSH, thyroid hormones in serum and tissue, and brain iron concentration. In order to assess for significant differences between two groups, a t-test was performed. To assess for significant differences between three groups, one-way analysis of variance with Tukey's post hoc test was performed. Values are expressed as means ± standard error of the mean. Differences were considered significant at p < 0.05.
Results
Evaluating iron status of rat dams and pups
The authors’ previous study established two types of iron-deficient rat dam models in imitation of ID with and without anemia in pregnant women. The first was the mild ID + A model, which mirrors pregnant women who do not have adequate iron status before pregnancy and go on to develop ID + A during pregnancy. The other is an ID – A model, which mirrors pregnant women who have less than adequate iron status before and during pregnancy but do not develop ID + A during pregnancy (20). That study found that both mild ID with anemia and ID without anemia resulted in maternal hypothyroxinemia from mid-gestation (G13) to the end of the pregnancy (P0; Supplementary Table S2). To investigate whether perinatal ID-induced maternal and neonatal hypothyroxinemia is an early mechanism of impairing brain development prior to neonatal brain iron depletion, the neonatal brain iron levels need to be evaluated. Due to the interference of circulating iron concentrations on brain iron levels and the difficulty to perfuse pups with physiological saline through the left ventricle in P0 rat pups, this study focused on P10 rat dams and pups. Consistent with the previous study, the present study also found that different degrees of pre-pregnant diet iron limitation induced ID with anemia and without anemia in P10 rat dams (Table 1).
Data are presented as means ± standard error of the mean (SEM; n = 10 for hemoglobin and liver iron; n = 6 for thyroid function, serum iron, and serum ferritin). One-way analysis of variance (ANOVA) and Tukey's post hoc test were used to establish significant differences between more than two groups. Differences were considered significant at p < 0.05.
p < 0.05 compared with the control; $ p < 0.05 compared with ID – A.
TT4, total thyroxine; TT3, total triiodothyronine; TSH, thyrotropin; P, postnatal day; ID – A, iron deficiency without anemia; ID + A, iron deficiency with anemia.
To evaluate the iron status of postnatal pups, liver iron and blood hemoglobin levels were measured. The blood hemoglobin level was reduced in the ID + A group, but it was not altered in the ID – A group, compared with the control group at P0 (Table 2). The liver iron level was reduced and the blood hemoglobin level was not altered in the ID – A group compared with the control group at P10. However, both liver iron and blood hemoglobin levels were reduced in the ID + A group compared with the control group at P10 (Table 3). The body weights of the pups were lower in the ID + A groups at P0 (Table 2) and in both the ID + A and ID – A groups at P10 (Table 3) compared with the control group.
Data are presented as means ± SEM (n = 42–48 in each group). One-way ANOVA and Tukey's post hoc test were used to establish significant differences between more than two groups. Differences were considered significant at p < 0.05.
p < 0.05 compared with the control; $ p < 0.05 compared with ID – A.
Data are presented as means ± SEM (cerebral cortex iron and hippocampus iron: n = 14 in each group; weight, hemoglobin, liver iron, serum TT4, serum TT3, serum TSH, brain tissue TT4,TT3: n = 30–36 in each group; TPO activity in the thyroid: n = 36 in each group; iron in the thyroid: n = 30 in each group). One-way ANOVA and Tukey's post hoc test were used to establish significant differences between more than two groups. Differences were considered significant at p < 0.05.
p < 0.05 compared with the control; $ p < 0.05 compared with ID – A.
TPO, thyroid peroxidase.
Perinatal ID induces hypothyroxinemia in both dams and pups
As described above, the authors’ previous studies documented that both mild ID with anemia and ID without anemia resulted in maternal hypothyroxinemia (low serum TT4) beginning at mid-gestation (G13) and continuing to the end of the pregnancy (P0; Supplementary Table S2). In the present study, maternal hypothyroxinemia was also found in ID + A and ID – A dams on P10 compared with the controls. A summary of thyroid hormone levels is included in Table 1.
To evaluate the thyroid hormone status of postnatal pups, serum TT4, TT3, and TSH concentrations were measured at P0 and P10. Serum TT4, TT3, and TSH concentrations did not differ between the groups in P0 pups (Table 2). However, serum TT4 showed a decreasing trend in ID pups, and it was significantly reduced in ID + A P10 pups (Table 3). Serum TT3 also showed a decreasing trend in ID pups, but no significant differences were found between the three groups (Table 3). These results indicate that ID dominantly affects serum TT4 levels but not TT3 levels in both dams and pups, and that it is dependent on the degree of ID. It is known that low maternal T4 levels, but not low maternal T3 levels, are associated with adverse effects on brain development in offspring (31 –33), since >80% of the T3 in rodent brain tissue may be produced by local deiodination of T4. It is speculated that this is the reason why TT3 levels were reduced in the cerebral cortex and hippocampus of both the ID – A and ID + A pups.
Perinatal ID decreases cerebral cortical and hippocampal thyroid hormone levels and associated thyroid hormone–responsive gene mRNA expression in P10 rat pups prior to neonatal brain iron depletion
It has been previously reported that maternal thyroid hormone deficiency impairs brain development, which suggests that the main effects of thyroid hormones are mediated by their receptors (TRs), which directly regulate gene expression (34,35). This study assessed the thyroid hormone levels in the cerebral cortex and hippocampus, as well as brain mRNA expression of several known thyroid hormone–responsive genes. Considering the interference of circulating thyroid hormone concentrations on brain thyroid hormone levels, P10 pups were perfused with physiological saline through the left ventricle. Table 3 shows that both ID protocols decreased TT3 levels in the cerebral cortex and the hippocampus in P10 pups, and the TT3 levels are almost 20% lower than those in the controls. Next, several thyroid hormone–responsive genes involved in development of the cerebral cortex and the hippocampus were selected: Rc3 (neurogranin); Camkiv (ca2+/calmodulin dependent protein kinase IV); Hairless; Mbp (myelin basic protein); Srg1 (synaptotagmin related gene 1); and Reelin; Dio2 (type 2 deiodinase). Figure 1A shows the effect of the ID protocols on gene expression at P10 in the cerebral cortex and the hippocampus. Mbp mRNA expression levels were significantly lower at P10 in both ID protocols in the hippocampus compared with controls, In contrast, Dio2 levels were higher in hippocampus at P10 in the two ID conditions. RC3, Mbp, and Srg1 mRNA expression levels were significantly lower at P10 in both ID protocols in the cerebral cortex compared with controls. These data are consistent with the decreased T3 levels observed in the cerebral cortex and the hippocampus in the ID pups exposed to the two ID protocols (Table 3).
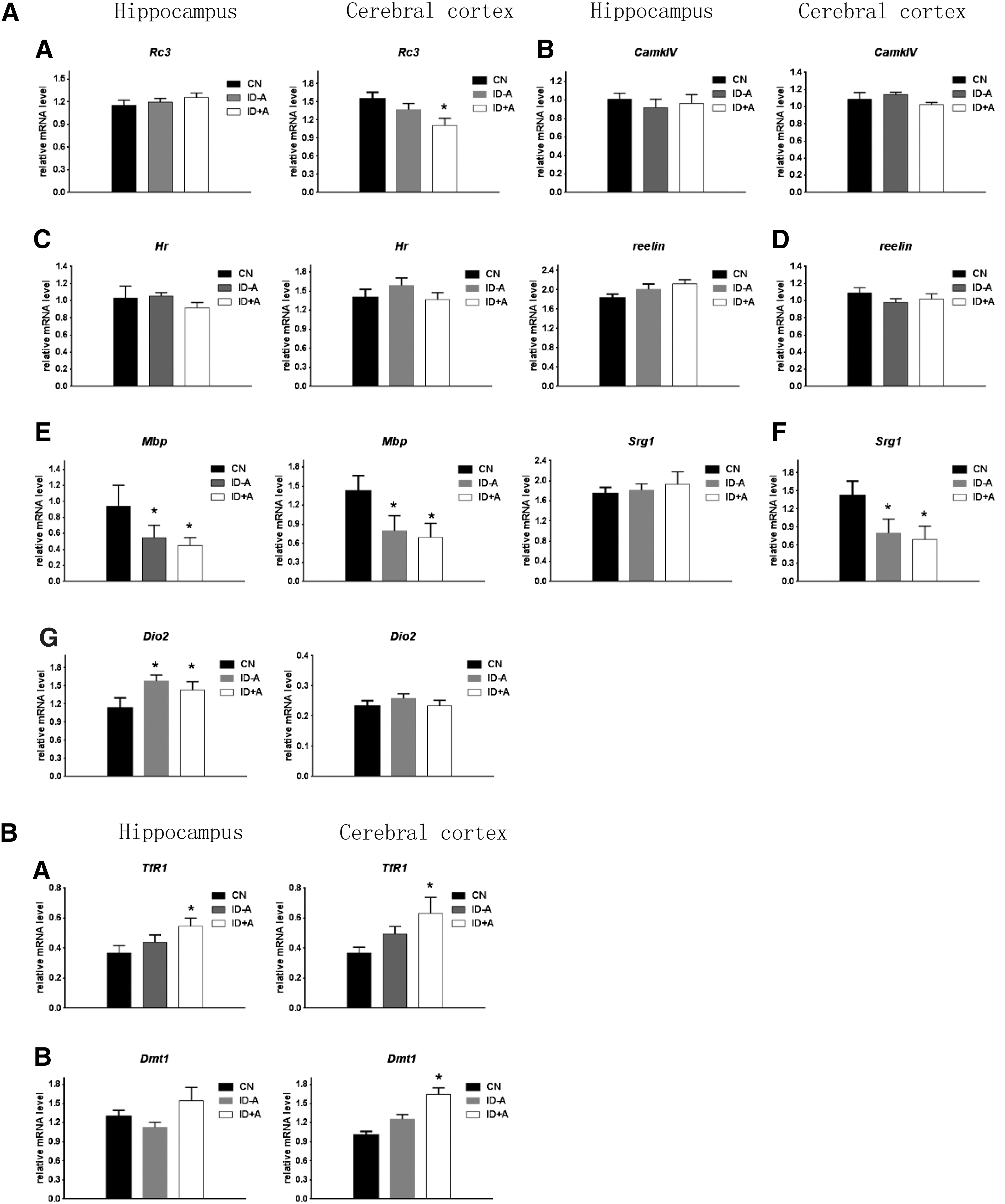
(
Iron concentrations in the cerebral cortex and hippocampus were not reduced in ID groups in P10 rat pups
To define whether the iron concentrations were altered in P10 pup brain tissue, iron concentrations were determined in the cerebral cortex and hippocampus using ICP-MS at P10 (Table 3). No significant difference was found between the controls and the ID pups. This result was consistent with the previous findings reported by Beard et al., which also found that moderate perinatal ID did not decrease the iron concentrations in the cerebral cortex and hippocampus in P10 rat pups (18). It has been reported that when brain cells are iron deficient, neurons may upregulate mRNA expression of iron-responsive genes, such as the transferrin receptor (TfR) and divalent metal transporter (DMT1), to ensure the cells acquire adequate iron levels (36 –38). In response to ID, TfR and DMT1 gene expression increases in pup brain tissue (36 –38). In the present study, TfR and DMT1 mRNA expression was also tested. Figure 1B shows that the mRNA expression of TfR and DMT1 did not significantly change in the cerebral cortex and the hippocampus in the ID – A group compared with the control group, which is consistent with the normal iron levels in the P10 neonatal rat brains. In the ID + A group, the iron concentrations and iron staining were unchanged in P10 brain tissues. However, TfR mRNA expression in the cerebral cortex and the hippocampus were higher compared with the control group (Table 3 and Fig. 1B). This is consistent with the concept that the brain attempts to maintain adequate iron levels by upregulating the expression of TfR and DMT-1 under ID conditions. The rat models developed in this study demonstrate that perinatal ID does not necessarily lead to early deficits in regional brain iron concentrations in P10 pups.
Perinatal ID results in delay of sensorimotor behaviors prior to neonatal brain iron depletion
To determine whether ID-induced hypothyroxinemia can induce mental retardation and neurobehavioral dysfunction prior to the neonatal brain iron depletion, sensorimotor skills were tested. Three peripheral systems, including the vestibular, exteroceptive, and proprioceptive systems, may be involved in regulation of postural adjustments, and the cerebral cortex is more involved in forelimb co-ordination (where placing is tested) (27). Upper extremity sensorimotor function assessed by the vibrissae-evoked forelimb placing test was chronically impaired by the damage to the forelimb region of the brain sensorimotor cortex (28). The pattern of sensorimotor skill achievement was obviously changed in ID postnatal pups. Surface righting was significantly delayed for the ID + A pups (Fig. 2A), and vibrissae-evoked forelimb placing was significantly reduced for both ID pups at P10 (Fig. 2B).
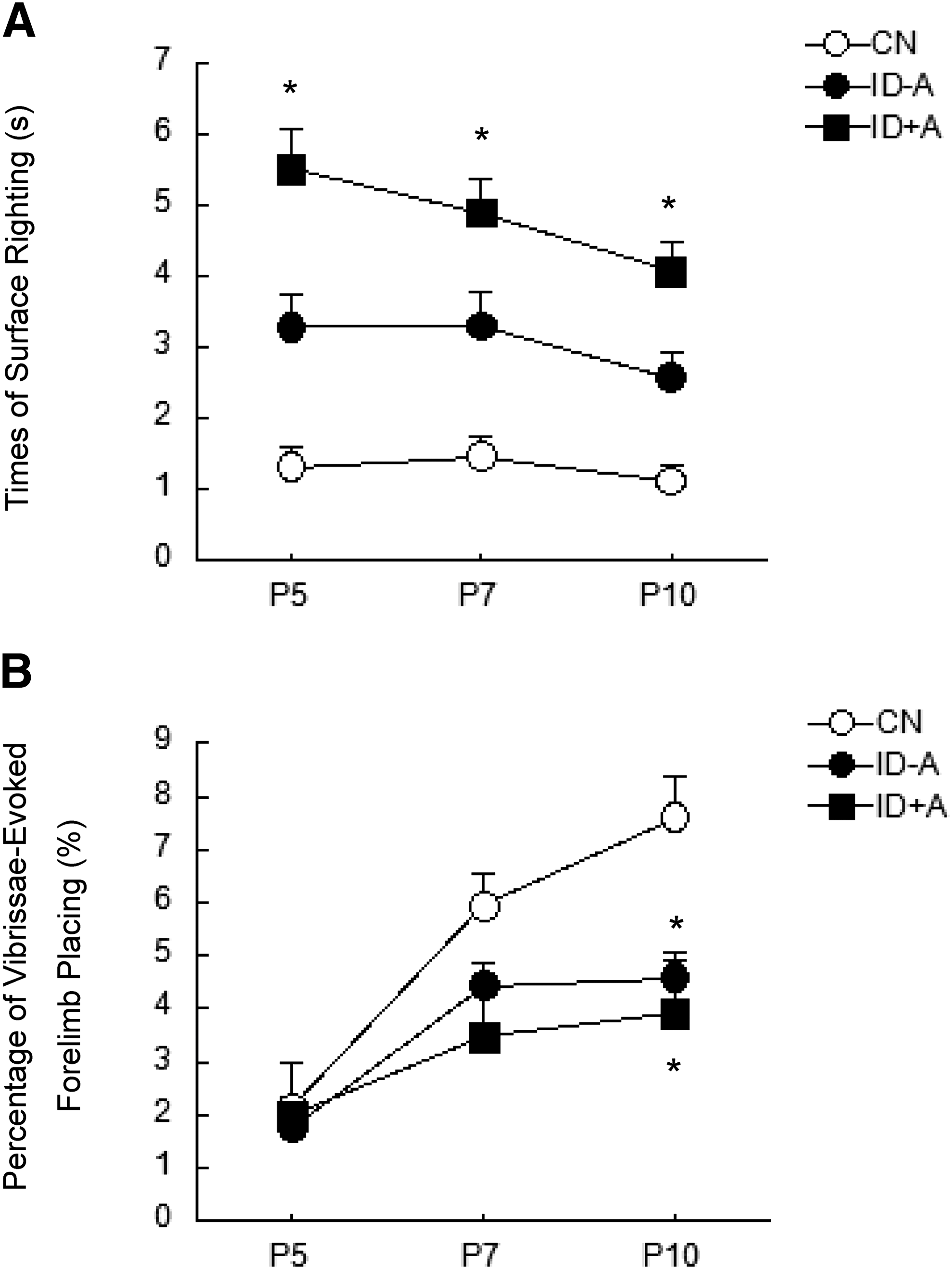
ID impairs surface righting and vibrissae-evoked forelimb placing in postnatal pups. Surface righting was significantly delayed for the ID + A pups at P5, P7, and P10. Vibrissae-evoked forelimb placing was significantly reduced for both ID – A and ID + A pups at P10. (
Discussion
ID is prevalent in pregnant woman. Iron requirements are greater during pregnancy than at other times, especially from the second trimester onward. This is due to the expansion of total red blood cell mass and the requirements of the fetus (39). In 2007, Zimmermann et al. first reported that ID was predictive of lower TT4 during pregnancy in a study of pregnant women living in an area with borderline iodine deficiency (40). The authors’ epidemiological study also found a higher frequency of maternal hypothyroxinemia in pregnant women with ID than those with sufficient iron storage, even in an iodine sufficient area (41). These data suggest that maternal ID may be an indicator for maternal hypothyroxinemia.
Presently, the mechanisms of perinatal ID-induced cognitive and behavioral function deficits in offspring are thought to be primarily caused by iron depletion in in the neonatal brain. In order to evaluate whether hypothyroxinemia associated with perinatal ID plays an important role in impairing brain development, two types of ID rat models were established that mimic ID with and without anemia in pregnant women (20). The present study found that perinatal ID results in poor neurobehavioral outcomes, regardless of normal iron levels in the neonatal brain, but during the same time period, perinatal ID also results in the decline of brain thyroid hormone levels and a decrease in the expression of thyroid hormone–responsive genes such as RC3, Mbp, and Srg1. These genes play important roles in neuronal synaptogenesis, dendritogenesis, and myelinogenesis in the cerebral cortex, as well as in hyppocampal development (42 –46). Myelin is a multi-lamellar protein that insulates mature axons and facilitates conduction of nerve impulses. Mbp is an essential protein involved in myelination (45). RC3 is a 78 amino acid brain-specific substrate for calmodulin binding and protein kinase C, which is mainly expressed in the soma and dendrites of the cerebral cortex and the hippocampus (46). Srg1 is also distributed in the axon and dendrites (44). Both RC3 and Srg1 are reported to regulate synaptic plasticity and dendritic spine formation. It is hypothesized that the altered thyroid status may contribute to neurodevelopmental defects prior to neonatal brain iron depletion.
This conclusion is supported by the two ID animal models used in this study. The rat ID models that were developed differ from previous investigations. Previous studies using severe ID rat models reported a 30–50% reduction in brain iron concentration in adulthood (12,47).
The dynamic changes of iron and thyroid parameters were assessed during the perinatal period in the present study. It was found that maternal hypothyroxinemia occurred first, that is, from mid-gestation throughout the mid-lactation period (Supplementary Table S2). The maternal hypothyroxinemia occurred at the same pace as the decline in maternal serum ferritin levels, and earlier than the decline of serum hemoglobin levels. Neonatal hypothyroxinemia and lower hemoglobin levels were present at P10, thus preceding neonatal brain iron depletion. In aggregate, the results demonstrate that under ID conditions, iron is prioritized to the fetal and neonatal brain and the synthesis of hemoglobin, but at the expense of iron-containing enzymes such as TPO, which then results in both maternal and neonatal hypothyroxinemia. These results are consistent with previous studies that investigated the impact of maternal iron status on neonatal iron stores, and also reported that the fetus can acquire sufficient iron, even under conditions of mild to moderate ID anemia (36,48). The enhanced expression of TfR and DMT1 were reported to be responsible for the functional increase in iron transfer during deficiency (49 –53).
Previous rat studies showed that serum TSH levels associated with ID vary between the different studies. Serum TSH levels were decreased in some studies in ID rats, but unaltered in other studies (54 –56). Pituitary TSH is negatively regulated by thyroid hormone. The pituitary conversion of T4 to T3 is catalyzed by an outer ring 5′-deiodinase enzyme, type 2 deiodinase (Dio2). Dio2 activity is regulated by thyroid function; for example, it increases in case of hypothyroidism, but decreases in hyperthyroidism. At the same time, Dio2 activity is thought to be decreased under ID conditions, since the deiodinases are iron-containing enzymes. Beard et al. measured the activity of the hepatic type 1 deiodinase (Dio1), but not the type 2 pituitary enzyme in iron deficient rats, and found that Dio1 activity was decreased in iron-deficient rats (55). But Kaplan and Utiger (57) earlier reported that the outer ring deiodinase activity was not affected by either ferrous or ferric ions in an in vitro study. Thus, as of yet, the mechanism of how ID regulates serum TSH remains unclear.
There are several limitations in this study. Although the study shows that perinatal ID results in the decline of brain thyroid hormone levels and the expression of thyroid hormone–responsive genes (such as RC3, Mbp, and Srg1), the mechanisms underlying this result were not investigated further. Mechanisms that are speculated to be involved in these alterations associated with brain thyroid hormone deficiency include impaired synaptogenesis and dendritogenesis of cortical neurons. Another limitation is that the protein levels of thyroid hormone–responsive genes and iron-responsive genes were not measured because the brain samples of P10 rat pups were not sufficient to perform Western blot analyses. The third limitation is that a group of ID animals substituted with thyroid hormone was not included. This would have provided more convincing data to characterize the consequences of ID-associated hypothyroxinemia, and would help to determine to what extent treatment with thyroid hormone can rescue the phenotype.
In summary, this study shows that perinatal ID can lead to maternal and (or) neonatal hypothyroxinemia, which impairs early brain development prior to any neonatal brain iron depletion. This indicates that the effect of hypothyroxinemia may be an early event, and reduced neonatal brain iron level may be a later event affecting brain development associated with perinatal ID. It is recommended that thyroid hormone levels be monitored in ID pregnant women. Considering the adverse effects of maternal and neonatal hypothyroxinemia on brain development in this animal study, it is necessary to treat maternal hypothyroxinemia. The question of whether treatment should involve levothyroxine supplementation or iron supplementation or both deserves further investigation in animal studies.
Footnotes
Acknowledgments
This work was supported by the National Science Foundation of China (30800918, 81570711, 81271511) Important Platform of Science and Technology for the universities in Liaoning Province (16010).
Author Disclosure Statement
The authors declare that they have no competing interest.