
Abstract
Background:
Iodine is essential for thyroid hormone synthesis and is an important regulator of thyroid function. Chronic iodine deficiency leads to hypothyroidism, but iodine excess also impairs thyroid function causing hyperthyroidism, hypothyroidism, and/or thyroiditis. This study aimed to investigate the underlying mechanisms by which exposure to chronic iodine excess impairs pituitary–thyroid axis function.
Methods:
Male Wistar rats were treated for two months with NaI (0.05% and 0.005%) or NaI+NaClO4 (0.05%) dissolved in drinking water. Hormone levels, gene expression, and thyroid morphology were analyzed later.
Results:
NaI-treated rats presented high levels of iodine in urine, increased serum thyrotropin levels, slightly decreased serum thyroxine/triiodothyronine levels, and a decreased expression of the sodium–iodide symporter, thyrotropin receptor, and thyroperoxidase mRNA and protein, suggesting a primary thyroid dysfunction. In contrast, thyroglobulin and pendrin mRNA and protein content were increased. Kidney and liver deiodinase type 1 mRNA expression was decreased in iodine-treated rats. Morphological studies showed larger thyroid follicles with higher amounts of colloid and increased amounts of connective tissue in the thyroid of iodine-treated animals. All these effects were prevented when perchlorate treatment was combined with iodine excess.
Conclusions:
The present data reinforce and add novel findings about the disruption of thyroid gland function and the compensatory action of increased thyrotropin levels in iodine-exposed animals. Moreover, they draw attention to the fact that iodine intake should be carefully monitored, since both deficient and excessive ingestion of this trace element may induce pituitary–thyroid axis dysfunction.
Introduction
T
Iodide uptake into thyrocytes is a limiting step for TH synthesis and is dependent on the activity of the sodium–iodide symporter (NIS), a glycoprotein expressed in the basolateral membrane of thyrocytes (2). Iodide is then transported across the apical membrane into the follicle colloid, presumably mainly by pendrin and anoctamin-1 (3,4). In the follicular lumen, iodide is oxidized and incorporated into the tyrosyl residues of thyroglobulin (Tg) by thyroid peroxidase (TPO). This enzyme is also responsible for iodotyrosine coupling and generation of TH within Tg (5). T4 and T3 are then released from Tg in the cytosol by lysosomal enzymes and subsequently secreted into the bloodstream. All these steps are regulated by thyrotropin (TSH) through stimulation of the TSH receptor (TSHR). TSHR is coupled to G-proteins and activates, among others, the protein kinase A (adenylyl-cyclase) and protein kinase C pathways; the cAMP-PKA signaling pathway appears to be the predominant pathway (6).
Iodine is a scarce element in the environment (7), and its consumption in natural (e.g., seafood) and processed foods as well as in iodinated salt is essential for TH synthesis. However, iodine deficiency is still a public health issue in many countries and corresponds to the primary cause of goiter and hypothyroidism. Iodine deficiency leads to adverse effects such as endemic goiter, cretinism with retardation of growth and abnormal development of the nervous system, including cognitive and neuromotor disabilities, and increased fetal and infant mortality (8).
In contrast, some areas are now exposed to high amounts of iodine, and studies have shown that iodine excess can disrupt thyroid function (9). For example, iodine excess has been associated with thyroid dysfunctions such as thyroiditis, hyperthyroidism, hypothyroidism, and thyroid cancer, especially papillary thyroid carcinoma in postmenopausal women (10,11).
According to the World Health Organization, urinary iodine concentration in children and adults should be 100–199 μg/L. However, in some Asian countries such as Japan and Korea, where seaweed ingestion is common, the concentration of urinary iodine is frequently >300 μg/L (12,13).
An acute and autoregulatory mechanism, known as the Wolff–Chaikoff effect, protects the thyroid gland against large amounts of iodine by inhibiting iodide uptake and organification (14). This effect is transitory, and after few hours of inhibition, synthesis and secretion of TH are restored (15). Despite these protective effects, a chronic iodine overload is related to the occurrence of thyroid lesions and impairment of thyrocyte function, which may be extremely harmful depending on the developmental stage at the time of exposure (16 –18).
Previous studies have shown that the pituitary–thyroid axis is impaired in iodine-excess treated mice/rats (19,20). Nevertheless, the molecular mechanisms that underlie thyroid dysfunction induced by exposure to iodine excess are not completely known. Therefore, this study aimed to characterize the underlying molecular mechanisms further by evaluating the effects of chronic iodine excess treatment on thyroid gene expression, thyroid cell morphology, and the function of the pituitary–thyroid axis in male Wistar rats.
Materials and Methods
Animals and treatments
Male Wistar rats weighing 200–250 g were obtained from the authors’ breeding colony and maintained on rat chow and tap water ad libitum. The animals were housed in a room kept at constant temperature (23 ± 1°C) and on a 12:12-h light–dark cycle schedule (lights on at 07:00 a.m.). The animals were divided in the following groups: • Control (C): animals supplied with normal tap water for 60 days. • I0.005%: animals treated with tap water supplemented with 0.005% NaI for 60 days (21). • I0.05%: animals treated with tap water supplemented with 0.05% NaI for 60 days (19,22,23). • I + P: Animals treated with tap water supplemented with 0.05% NaI and 0.05% NaClO4 for 60 days.
Iodine treatment doses were chosen based on several previous studies that evaluated the effect of either acute or chronic iodine excess exposure on thyroid function, both in mice and rats. Moreover, Wistar rats have no tendency for autoimmunity. Therefore, this animal model allowed the effects of iodine excess to be explored in a model that is resilient to developing autoimmune thyroid disease.
After treatment, urine was collected to determine the urinary concentration of iodine. Subsequently, the rats were anesthetized and euthanized by decapitation. The thyroid gland, pituitary gland, kidney, and liver were rapidly excised for total RNA and/or protein extraction. Moreover, thyroid gland lobes were weighed and used for morphological and histological analysis, as described below. Blood samples were collected from the trunk to evaluate serum TSH, T4, and T3. Hearts were also collected to determine the ratio between dry heart weight and body weight. The experimental protocol was approved by the Institute of Biomedical Sciences/University of São Paulo-Ethical Committee for Animal Research (no. 89/2012), and the protocols are in accordance with the ethics principles in animal research adopted by the National Council for the Control of Animal Experimentation.
Determination of TH and TSH serum levels and urinary iodine concentration
TSH, T4, and T3 rat serum levels were assessed using the Milliplex Luminex® kit #RTHY-30K (EMD Millipore, Billerica, MA). The modified Sandell–Kolthoff method was used to assess the concentration of iodine in the urine samples (10).
Real-time polymerase chain reaction analysis
Thyroid gland, pituitary gland, kidney, and liver total RNA extraction was performed through the thiocyanate–phenol–chloroform extraction method, according to a standard protocol (24). The expression of Tsha, Tshb, Dio1, Slc5a5/Nis, Tpo, Slc26a4/Pds, Tg, Tshr, and Tgfb1 mRNAs was evaluated by real-time polymerase chain reaction, according to the manufacturer's instructions (Invitrogen, Carlsbad, CA). Gene-specific primer sequences are described in Supplementary Table S1 (Supplementary Data are available online at
Protein expression
Thyroid and pituitary protein extraction was performed as previously described (26). Briefly, the glands were homogenized in RIPA buffer (20 mM of Tris pH 7.5, 150 mM of NaCl, 1% Nonidet P-40, 0.5% sodium deoxycholate, 1 mM of EDTA, 0.1% SDS and protease inhibitor cocktail). Proteins were quantified by a Bradford assay (Bio-Rad, Hercules, CA) and resolved by 6% or 10% SDS-PAGE and transferred to nitrocellulose membranes. Membranes were blocked and incubated with primary antibodies against Tsha, Tshb, Nis, Tshr, Tpo, Pds, Tg, and Tgfβ (Supplementary Table S2). Equal loading was evaluated by Ponceau staining or by incubating the membranes with a primary antibody against Gapdh or α-Actinin. Blots were developed using the enhanced chemiluminescence kit (Amersham Biosciences, Little Chalfont, United Kingdom). Blot densitometry was analyzed by using Image J software (National Institutes of Health).
Thyroid histological analysis
Thyroid glands were excised and incubated in 10% buffered formalin. Samples were fixed at 4°C for 3 h in Methacarn (absolute methanol, chloroform, and glacial acetic acid; 6:3:1), rinsed with absolute ethanol, and embedded in Paraplast (Merck, St. Louis, MO) at 60°C. Thyroids were cut into 5 μm sections, adhered to glass slides using 0.1% poly-L-lysine (Sigma, St. Louis, MO), dried at 37°C, and stored at room temperature. The sections were stained with hematoxylin and eosin, and then examined by using a Nikon Eclipse E600 microscope. Images were captured using a digital camera (Cool SNAP-Procf color; Roper Scientific, Trenton, NJ), and the morphometric characteristics were analyzed by the Image Pro Plus software (Media Cybernetics, Silver Spring, MD).
Statistical analysis
All data are reported as means ± standard error of the mean. Statistical analysis was performed using GraphPad Prism (GraphPad Software, San Diego, CA). Outliers were removed using Grubbs’ test. Data were subjected to unpaired one-way analysis of variance (ANOVA) followed by a Student–Newman–Keuls post hoc test or unpaired two-tailed Student's t-test. Differences were considered statistically significant at p < 0.05. Data were also subjected to two-way ANOVA in order to analyze inter-experiment variance.
Results
Evaluation of iodine excess treatment effectiveness
The effectiveness of the iodine treatment was confirmed by the increased urine concentration of iodine in the animals chronically exposed to NaI compared with the control group (Table 1). As expected, animals treated with 0.05% NaI presented higher concentration of iodine in the urine comparison with the animals treated with the lower dose (0.005%).
Results are expressed as means ± SEM, n = 10 per group.
Values multiplied by 1000.
p < 0.05; ** p < 0.01; *** p < 0.001 vs. control.
UIC, urinary iodine concentration; BW, body weight; DHW, dry heart weight; DHW/BW, dry heart weight to body weight ratio; SEM, standard error of the mean.
Effects of chronic iodine excess on body weight and dry heart weight
Body weight was not altered in rats exposed to iodine excess treatment compared with the control group (Table 1). In contrast, dry heart weight and the ratio between this parameter and body weight were decreased in the animals treated with the higher dose of NaI (0.05%). No significant changes in these parameters were observed in the animals treated with the lower dose of NaI (0.005%; Table 1).
Effect of chronic iodine excess on T3, T4, and TSH serum concentrations
As shown in Table 2, chronic treatment of rats with 0.05% NaI significantly decreased T4 and T3 serum concentrations. Moreover, this treatment increased TSH levels compared with the control group. These effects were prevented when the animals were concomitantly exposed to NaI and NaClO4. Additionally, the lower dose of iodine treatment (0.005% NaI) was able to decrease T3 and increase TSH serum concentrations, even though no significant changes in T4 serum concentration were observed.
Results are expressed by means ± SEM, n = 6 per group.
p < 0.05; ** p < 0.01 vs. control.
T3, triiodothyronine; T4, thyroxine; TSH, thyrotropin.
Effects of chronic iodine excess on Tsh gene expression
Considering that both 0.005% and 0.05% NaI doses significantly increased serum TSH concentrations, the pituitary expression of Tsha and Tshb was investigated. While mRNA and protein expression of alpha Tsh subunit was not altered by chronic iodine exposure (Fig. 1A and B), pituitary Tshb mRNA expression was increased in rats exposed to 0.05% NaI, even though the Tshb protein content was not altered by excessive iodine treatment (Fig. 1C and D). The inter-experiment variance analysis showed no significant differences between the independent experiments (Supplementary Figs. S1 and 2).
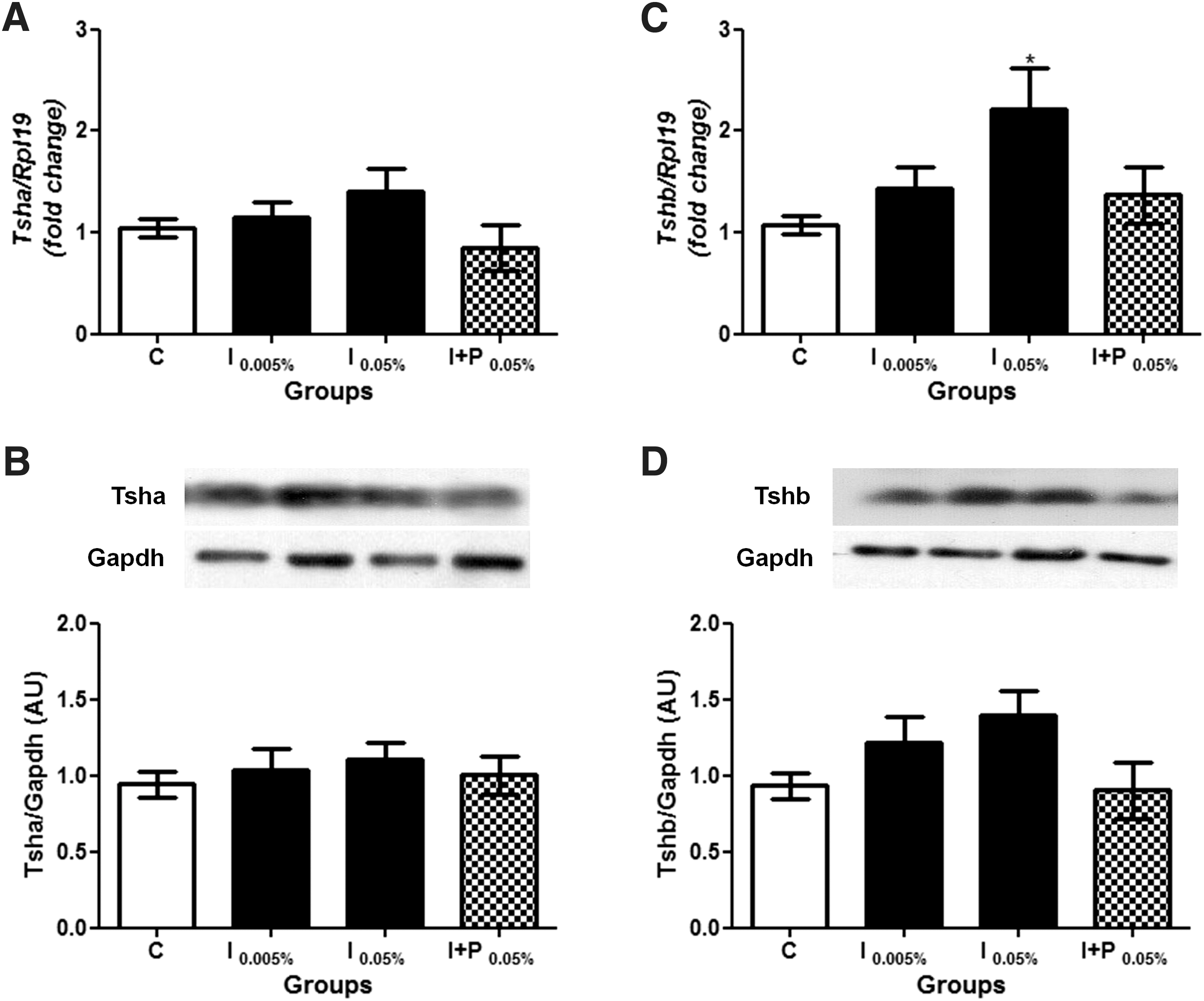
Effect of chronic iodine excess on Tsha and Tshb pituitary expression. (
Effects of chronic iodine excess on thyroid gland morphology
The appearance and the weight of the thyroid lobes were not significantly changed after eight weeks of iodine excess treatment in the two groups exposed to iodine treatment (Fig. 2A and B), even though the histological analysis showed altered thyroid follicle structures compared with the controls. The morphometric analysis indicated that 80–90% of the thyroid follicles of the C and I + P (NaI + NaClO4) groups and 50–60% of the follicles of the iodine-treated groups measured between 10 and 30 × 103 μm2 (Fig. 2C). However, 40–50% of the thyroid follicles of the iodine-exposed animals presented a larger area than the control and I + P groups.

Effects of chronic iodine excess treatment on thyroid morphology. (
There was a clear increase of the follicular lumen with higher Tg accumulation in the iodine-treated groups (Fig. 2E and F) compared with the control group (Fig. 2D). However, thyrocytes flattened only in the group exposed to the higher dose of iodine (Fig. 2F). Moreover, there was a significant increase in the extracellular matrix around the thyroid follicles, without immune cell infiltration in iodine-treated animals. Finally, the concomitant treatment of the rats with iodine and perchlorate prevented the effects triggered by chronic iodine excess on thyroid morphology (Fig. 2G), and no differences were detected between the I + P (Fig. 2G) and the control (Fig. 2D) groups.
Since Tgfβ expression is implicated in the induction of fibrosis in several tissues (27), and considering the increased extracellular matrix around the thyroid follicles in iodine-treated rats and its relationship with fibrosis, Tgfβ1 mRNA and protein content were determined in the experimental groups. Figure 3 shows that both Tgfβ1 mRNA and protein expression were increased in the iodine-treated animals comparison with the C and I + P groups. No significant difference was observed between the independent experiments (Supplementary Fig. S3).
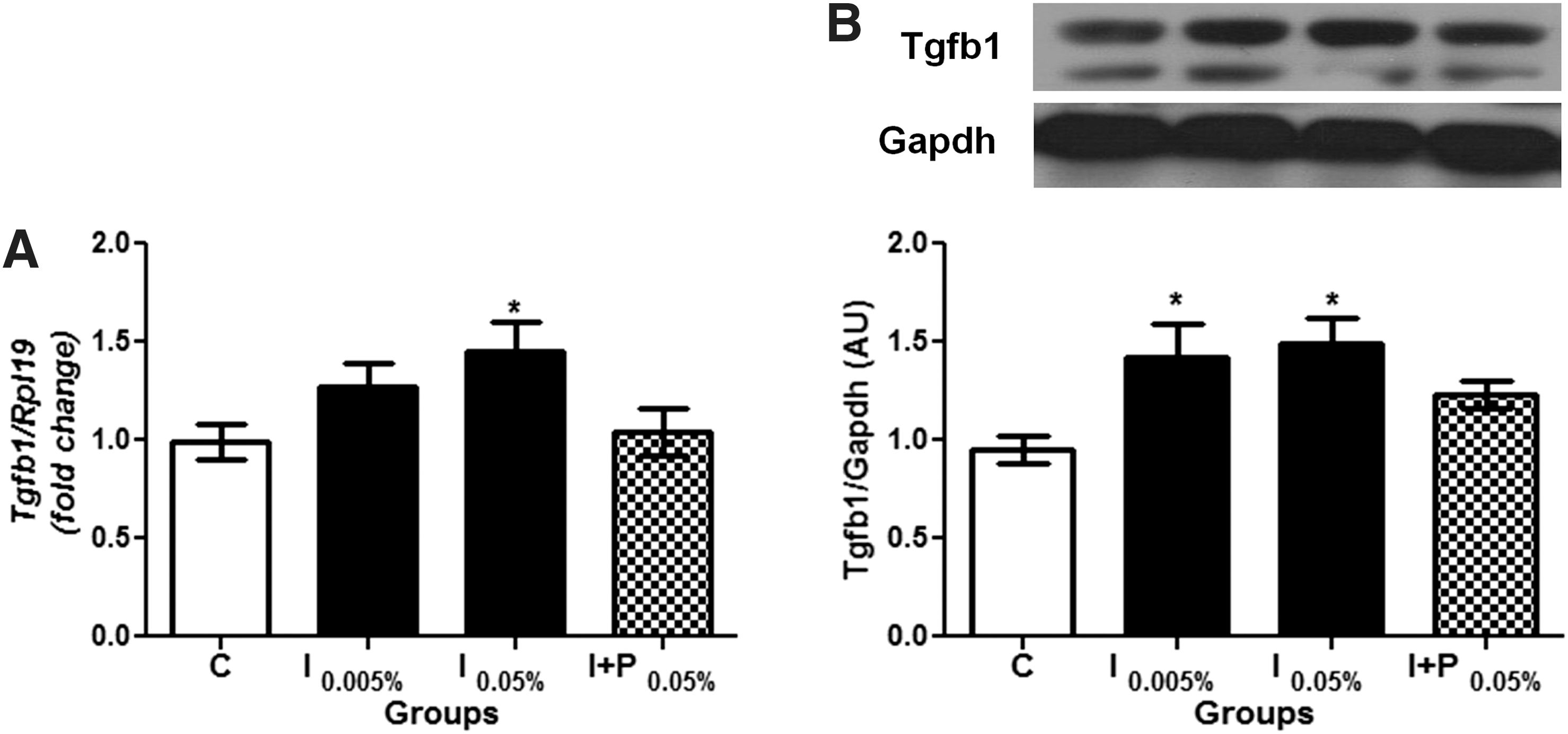
Chronic iodine excess augments thyroid Tgfb1 mRNA and Tgfb1 protein expression. (
Effects of chronic iodine excess on thyroid gene expression
TH synthesis depends on the expression and activity of different proteins in the thyroid gland. Therefore, this study evaluated whether treatment with excessive amounts of iodide could impair their expression. First, Slc5a5 mRNA and Nis protein expression were evaluated, and it was observed that both were decreased by iodine treatment, as shown in Figure 4A and B. Regarding Tshr mRNA and protein expression (Fig. 4C and D, respectively), the results also indicate that both are downregulated in animals exposed to 0.05% and 0.005% NaI treatment, even though the change in Tshr mRNA expression failed to reach statistical significance at the lower dose of iodine treatment. Furthermore, iodine treatment induced a downregulation of Tpo mRNA and protein expression, which reached significant values at the higher dose of iodine treatment (Fig. 4E and F).
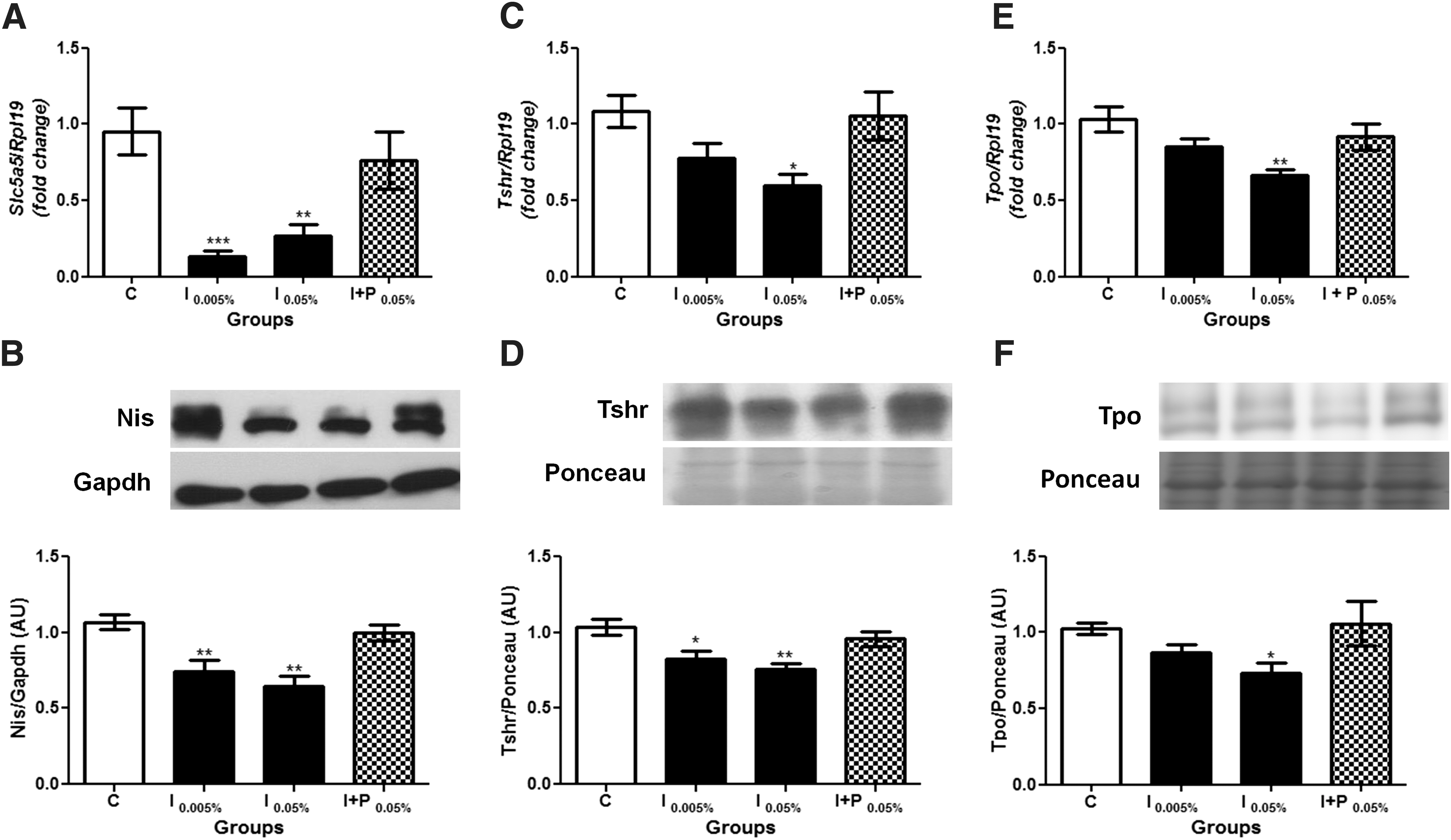
Chronic iodine excess treatment reduces thyroid Nis, Tshr and Tpo expression. (
In contrast, both doses of iodine treatment increased Slc26a4 mRNA and Pds protein expression, as shown in Figure 5A and B, respectively. Finally, while both doses of iodine exposure did not alter Tg mRNA expression (Fig. 5C), they significantly increased Tg protein content (Fig. 5D). It is worth noting that the concomitant treatment of rats with NaI and NaClO4 prevented all the effects induced by excessive iodine treatment, as shown in Figures 4 and 5. The inter-experiment variance analysis of all above-mentioned genes showed no significant differences between the independent experiments (Supplementary Figs. S4–S8).
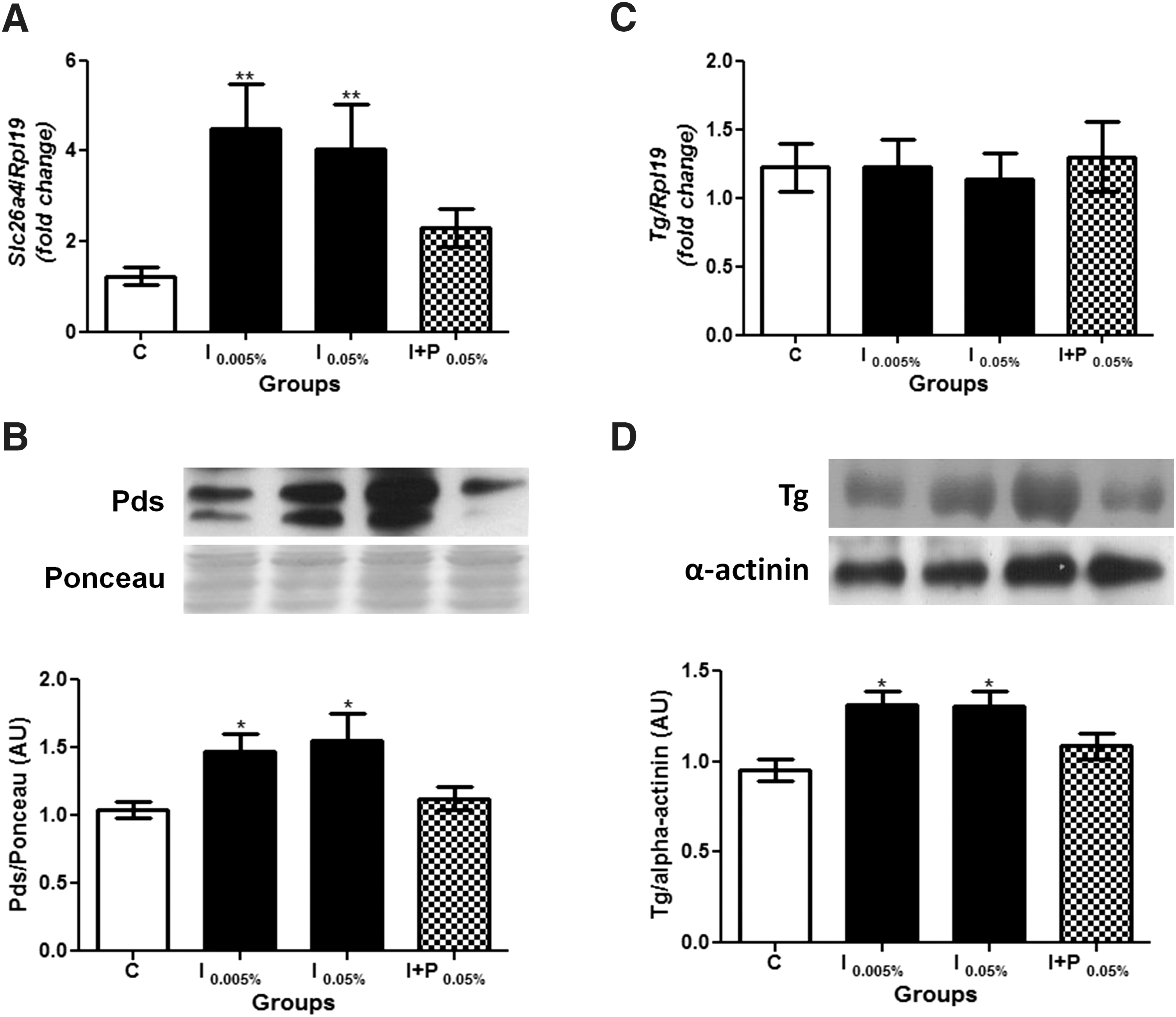
Chronic iodine excess treatment increases thyroid Pds and Tg expression. (
Effects of chronic iodine excess on Dio1 mRNA expression
Since both doses of iodine treatment were able to reduce T3 serum concentrations and that, in rats, approximately 45% of the circulating T3 comes from the peripheral conversion of T4 to T3 by D1 (28), this study evaluated Dio1 mRNA expression under these conditions. The results demonstrate that both doses of iodine treatment significantly reduce Dio1 mRNA expression in the kidney and liver (Fig. 6A and B). These effects were prevented in the presence of perchlorate. No difference was observed between the independent experiments (Supplementary Fig. S9).

Effects of chronic iodine excess treatment on kidney and liver type 1 iodothyronine deiodinase (Dio1) mRNA expression. Dio1 mRNA expression was analyzed by real-time PCR in the kidney (
Discussion
The current study supports previous findings that reported alterations of thyroid function in response to high iodine ingestion. Moreover, the data provide new insights into the molecular mechanisms underlying pituitary–thyroid axis impairment and the repercussions of chronic iodine excess (0.05% and 0.005%) on TSH synthesis and secretion, TH synthesis, thyroid morphology, and the peripheral conversion of T4 to T3. The data indicate that iodine excess disrupts the pituitary–thyroid axis, even in the animals treated with a low dose of iodine.
Although the iodine doses used in this study are extremely high compared with the usual levels of iodine consumption in humans, they have been extensively used in studies that aimed to evaluate the effects of iodine excess in rats and mice (19,21 –23). Moreover, it is well known that Wistar rats are highly resistant to developing thyroid autoimmune disorders in response to iodine excess (19,29). Therefore, the effects of iodine excess in the absence of an autoimmune background in this experimental model differ from most studies presented in the literature.
The present data indicate that TSH serum concentrations are markedly elevated in the animals exposed to iodine excess, even though serum T4 and T3 levels only showed minor differences in treated animals. In fact, pituitary expression of Tshb mRNA was increased under conditions of chronic iodine excess. However, Tshb protein expression was not significantly altered in the iodine-treated animals, suggesting an increased secretion rate of TSH into the blood, as indicated by the increased serum TSH concentrations. In accordance with these findings, a recent study demonstrated that prolonged high iodine intake inhibits pituitary D2 activity and consequently increases serum TSH levels (20). The regulation of D2 activity by iodine treatment adds a new aspect because high doses of iodine might interfere with thyroid function acting both on the thyroid gland and the pituitary gland, thus affecting TH synthesis and secretion at multiple levels.
Besides the increased levels of TSH, the data also reveal reduced serum levels of T3 in rats subjected to both doses of iodine treatment, and reduced T4 levels in the animals exposed to the highest dose of iodine. Therefore, the data suggest a primary impairment of thyroid gland function under conditions of chronic iodine excess that is compensated by increased levels of TSH. The group of rats treated with the lower dose of iodine presented a similar hormonal pattern as observed in subclinical hypothyroidism, since in this condition, TSH levels are increased but no changes in total T4 are observed (30 –32). Indeed, subclinical hypothyroidism is commonly found in clinical practice, and epidemiological studies have suggested a correlation between excessive iodine consumption and increased levels of TSH in normal populations (31,33).
In contrast, the administration of 0.05% NaI to the rats may be related to the induction of overt hypothyroidism (32). In fact, previous studies have suggested that subclinical hypothyroidism may progress to overt hypothyroidism in human patients (34). Moreover, it is possible that longer periods of treatment could induce more significant alterations in TH serum levels, as demonstrated by Wang et al. (29).
The inhibitory effect of iodine on serum TH concentration has also been demonstrated in several studies in humans and mice (17,35,36). In fact, the primary dysfunction of the thyroid gland, as suggested by the present data, may be associated with alterations in thyroid morphology, decreased expression of proteins involved with TH biosynthesis, and/or to an impairment of peripheral TH metabolism.
Thyroid histological analysis indicated a significant enlargement of thyroid follicles and an increased amount of extracellular matrix around them in response to iodine excess treatment. In fact, thyroid follicle enlargement may be related to the impaired secretion of TH, as demonstrated by the serum TH levels data. Moreover, it is known that thyroid TGFβ expression is increased by iodine treatment and that this cytokine stimulates the production of extracellular matrix proteins and inhibits their degradation (37,38). This effect may be associated with the presence of fibrous tissue in the thyroid gland (39). Therefore, the increased Tgfβ expression induced by both doses of iodine treatment described in the present study might be implicated in the development of fibrous tissue in the thyroid gland of the animals. Since no lymphocyte, macrophage, or neutrophil infiltration was observed in the thyroid tissue, it is speculated that thyrocytes could be responsible for the synthesis and secretion of this cytokine.
Additionally, even though previous studies have demonstrated that female mice exposed to iodine develop goiter and hypothyroidism (19), the present data did not indicate alterations in thyroid weight after iodine treatment. This discrepancy may be explained by the longer period of iodine treatment, as well as by the immune response observed in the animals in the study by Li et al. (19) compared with our experimental model.
Besides the morphological changes in the thyroid gland, significant alterations in thyroid gene expression were observed in iodine-treated rats. It is well known that TSH is the main positive regulator of all steps involved in TH synthesis by means of its binding to the TSHR expressed at the basolateral membrane of thyrocytes (40,41). In the present experimental model, chronic iodine excess treatment reduced Tshr expression, suggesting that thyrocyte responsiveness to TSH is impaired in both treatment conditions.
It is well known that TSH upregulates Slc5a5 gene transcription, NIS synthesis, as well as NIS activity, which is a limiting step in TH synthesis (42,43). The data presented here indicate that total Slc5a5 mRNA and Nis protein contents were reduced by the iodine treatment at both concentrations. Even though the impairment of TSH signaling could explain the decreased Nis expression, the possibility cannot be discarded of a direct inhibitory effect of iodine on Slc5a5 gene expression (44). Additionally, previous studies have demonstrated reduced Slc5a5 mRNA expression in animals chronically treated with iodine excess (9,19). The present results corroborate these results and demonstrate, for the first time, that total Nis content is reduced by treatment of thyroid cells with iodine.
This study also reveals a decreased Tpo mRNA and protein expression after the exposure of rats to the higher dose of iodine. It is well known that TSH is the main positive regulator of Tpo mRNA expression in thyrocytes (45,46). Even though TSH serum levels were increased in iodine-exposed animals, both Tshr mRNA and protein expression were decreased by iodine treatment. Therefore, impaired Tpo expression in iodine-treated animals could be a consequence of an impaired TSH signaling cascade. As mentioned before, a direct effect of iodine on the reduction of Tpo expression cannot be ruled out, since previous studies have reported inhibitory effects of acute iodine administration on the regulation of this gene (26,47). However, as far as it is known, this is the first time that the chronic administration effect of iodine has been shown to reduce both Tpo mRNA and protein content.
Interestingly, both concentrations of iodine utilized in this study have increased Slc26a4 mRNA and Pds protein expression. A previous study has demonstrated that Pds protein expression is positively regulated by TSH (48). Considering that the Tshr mRNA and protein content were reduced in iodine-treated animals and that previous studies have indicated a stimulatory effect of iodine on PDS gene expression (49), the present data strongly suggest that increased Slc26a4 mRNA and Pds protein expression are mainly mediated by iodine per se.
Additionally, increased Tg protein content was observed in the iodine-treated animals. Tg expression and TH secretion are known to be stimulated by TSH (40,50). However, as described, the Tshr content was significantly decreased in this experimental model. Therefore, it is mostly unlikely that the increased Tg content is related to TSH action on its synthesis. It seems more plausible that this finding could derive from the reduced action of TSH on TH secretion. This hypothesis is strengthened by the histological data, which indicate an increased size and amount of colloid in thyroid follicles of iodine-treated animals, thus suggesting decreased thyroid activity with impaired TH secretion.
Previous studies have shown that the treatment of thyrocytes in vitro with Tg decreased the expression of Nis, Tpo, and Tshr but increased Pds expression (51,52). Furthermore, it has been described that a high degree of iodinated Tg in the follicular lumen inhibits thyroid-specific gene expression (53). These effects may be mediated by the binding of Tg to the asialoglycoprotein receptor (ASGPR) expressed at the apical membrane of thyrocytes (54). Remarkably, it has been suggested that Tg maintains thyroid homeostasis by acting as a negative-feedback regulator of gene expression in thyrocytes (55).Therefore, in addition to the direct effects of iodine on thyroid and on the TSH signaling cascade, chronic iodine excess could also indirectly alter the expression of genes involved in TH synthesis through an augmented content and/or alterations in the levels of glycosylation of Tg in the follicular lumen.
In addition to the impairment of the expression of thyroid proteins related to TH synthesis, the reduction of T3 serum levels may be associated with the effects of iodine excess on peripheral tissues of the exposed rats. D1 is responsible for thyroidal and peripheral conversion of T4 to T3 (56). The present data indicate a reduction in Dio1 mRNA expression both in the kidney and liver after treatment with an excessive dose of iodine. Even though the reduced expression of Dio1 mRNA could suggest a reduced D1 content and consequently impaired peripheral conversion of TH, it is well known that D1 is upregulated by T3 (57). Thus, the reduced content of Dio1 mRNA expression could be a consequence rather than the cause of reduced serum T3 levels. Of note, previous studies have shown decreased D1 activity in the thyroid of Wistar rats chronically treated with iodine (29,58). In accordance with this, the preliminary data in this study suggest that D1 activity is reduced both in the kidney and in the liver, especially at the highest dose of iodine at excessive concentrations (data not shown). Therefore, the reduced serum levels of T3 after chronic iodine treatment may be linked to an impaired T4 deiodination both in the thyroid and in peripheral tissues.
Additionally, the reduced circulating levels of T3 were also confirmed by the decreased dry heart weight in the iodine-exposed animals. The heart is a classical target of T3 action, and the expression of several structural and functional proteins is upregulated by T3 (59). Therefore, the reduction of T3 serum levels is commonly associated with the reduction of the heart weight in animals (60,61).
Interestingly, the effects triggered by iodine excess on thyroid function and morphology were reversed by the concomitant treatment of the animals with NaI and NaClO4. Perchlorate is known to be a competitive inhibitor with iodide for the NIS protein, thus inhibiting thyroid iodide uptake (62). Therefore, since iodide uptake is inhibited by perchlorate, the data suggest that the effects promoted by chronic iodine treatment depend on thyroid iodide uptake and increased intracellular levels of iodide. It is relevant to mention that the concomitant exposure to perchlorate and iodine in this study has not altered serum TH levels in the rats, even though it has been shown that individuals exposed to doses of perchlorate than are higher than usual environmental exposures decreased the serum levels of TH (63,64). Perchlorate exposure has been shown to be particularly detrimental to thyroid function in iodine-deficient populations (65). However, the large amounts of iodine that were administered concomitantly with perchlorate in this study might have compensated the inhibition of NIS-mediated iodide transport, therefore resulting in sufficient uptake of iodine to maintain normal TH synthesis.
In summary, the data further elucidate the mechanisms involved in the disruption of thyroid function induced by chronic iodine excess. The compensatory effect of increased TSH serum levels emphasize the importance of maintaining iodide within appropriate limits, since both iodine deficiency and excess can impair the function of the pituitary–thyroid axis with potential repercussions on health.
Footnotes
Acknowledgments
The authors are grateful to Leonice Lourenço Poyares for excellent technical support, to Dr. Telma Maria Tenório Zorn for contributing to the histological analysis, to Dr. Marcus Vinicius C. Baldo and Rosana Prisco for the statistical analysis support, and to Astrid Freitas Meira for English review.
This work was supported by grants and fellowship from Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) to M.T.N. (473924/2010-2) and R.C.L. C.S.N. and R.B.S. are recipients of fellowships from Fundação de Amparo a Pesquisa do Estado de São Paulo (FAPESP) 2012/24391-6 and 2014/22934-8, respectively. J.C.S. is recipient of a fellowship from Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES; PVE A112/2013).
Author Disclosure Statement
The authors have nothing to disclose.