
Abstract
Background:
Thyroid hormone and leptin are essential regulators of energy homeostasis. Both hormones stimulate energy expenditure but have opposite effects on appetite. The mechanisms behind food intake regulation in thyroid dysfunctions are poorly understood. It has been shown that hypothyroid rats exhibited impaired leptin anorexigenic effect and signaling in total hypothalamus, even though they were hypophagic. It was hypothesized that hypothyroidism modulates the expression of neuropeptides: orexigenic neuropeptide Y (NPY) and anorexigenic proopiomelanocortin (POMC), independently of inducing nuclei-specific changes in hypothalamic leptin signaling.
Methods:
Adult male rats were rendered hypothyroid by administration of 0.03% methimazole in the drinking water for 21 days. Protein content of NPY, POMC, and leptin signaling (the signal transducer and activator of transcription 3 [STAT3] pathway) were evaluated by Western blot, and mRNA levels by real time reverse transcription polymerase chain reaction in arcuate (ARC), ventromedial (VMN), and paraventricular (PVN) hypothalamic nuclei isolated from euthyroid (eu) and hypothyroid (hypo) rats. Leptin anorexigenic effect was tested by recording food intake for two hours after intracerebroventricular (i.c.v.) administration of leptin. Statistical differences were considered significant at p ≤ 0.05.
Results:
Hypothyroidism was confirmed by decreased serum triiodothyronine, thyroxine, and increased thyrotropin, in addition to increased levels of pro-TRH mRNA in PVN and Dio2 mRNA in the ARC of hypo rats. Hypothyroidism decreased body weight and food intake associated with decreased protein content of NPY and increased content of POMC in the ARC. Conversely, hypothyroidism induced central resistance to the acute anorexigenic effect of leptin, since while euthyroid rats displayed reduced food intake after leptin i.c.v. injection, hypothyroid rats showed no response. Hypothyroid rats exhibited decreased leptin receptor (ObRb) protein content in ARC and VMN but not in PVN nucleus. ObRb protein changes were concomitant with decreased phosphorylated STAT3 in the ARC, and decreased total STAT3 in VMN and PVN. However, hypothyroidism did not affect mRNA levels of Lepr or Stat3 in the hypothalamic nuclei.
Conclusions:
Experimental hypothyroidism induced a negative energy balance accompanied by decreased NPY and increased POMC protein content in the ARC, resulting in predominance of anorexigenic pathways, despite central leptin resistance and impairment of the leptin signaling cascade in a nuclei-specific manner.
Introduction
T
The hypothalamus plays an essential role in the regulation of energy homeostasis by receiving and integrating multiple neural and peripheral signals, such as leptin, that reflect the nutritional and energetic state of the body. Three hypothalamic nuclei are essential for energy balance regulation: the arcuate nucleus (ARC), the ventromedial hypothalamic nucleus (VMN), and the paraventricular nucleus (PVN). In the ARC, the activation of proopiomelanocortin (POMC)-synthesizing neurons decreases food intake and stimulates energy expenditure, while activation of neuropeptide Y (NPY)/agouti-related protein (AgRP)-synthesizing neurons have opposite effects. These neuronal populations project to the PVN, targeting melanocortin receptor 4 and NPY receptors to produce part of their effects on energy metabolism (9). Additionally, there is substantial evidence that VMN neurons project to the ARC, regulating the activity of POMC and NPY/AgRP neurons, which in turn also regulate VMN neurons activity (9 –11).
In vivo administration of triiodothyronine (T3) directly into the ARC or VMN increases food intake (6,8). T3-induced hyperphagia has been associated with increased mRNA levels of Npy and Agrp and decreased Pomc mRNA levels in the ARC (3,5,8). Data regarding the effect of hypothyroidism on the regulation of these neuropeptides are scarce and inconclusive. One study reported decreased levels of Agrp mRNA but no alterations in Npy and Pomc mRNA expression in the ARC of hypothyroid rats (4). Another study demonstrated reduced levels of Pomc mRNA but no alteration in Npy mRNA expression in the ARC of hypothyroid mice (5). However, all of these studies are based on mRNA expression analyses, and, to the best of the authors' knowledge, there are no studies demonstrating the regulation of the protein content of these neuropeptides in thyroid dysfunctions.
The white adipose tissue–derived hormone leptin plays a major role in the regulation of energy homeostasis, acting at the hypothalamus to induce satiety and energy expenditure. The long form of the leptin receptor (ObRb) is abundantly expressed in the ARC and VMN, and to a lesser extent in the PVN. In the ARC, leptin stimulates the expression of POMC and inhibits the expression of NPY and AgRP (12 –15). Leptin effects are mediated by binding to ObRb and subsequent activation of intracellular cascades, mainly the Janus Kinase/Signal Transducer and Activator of Transcription 3 (JAK/STAT3) signaling pathway, which is crucial to leptin hypothalamic effects on energy homeostasis. Phosphorylated STAT3 (pSTAT3) translocates to the nucleus and regulates target genes, among them the Suppressor of Cytokine Signaling 3 (SOCS3), which in turn inhibits the JAK-STAT pathway constituting a negative feedback loop (16,17).
It has previously been demonstrated that methimazole-induced hypothyroidism in adult rats impairs hypothalamic leptin signaling and the anorectic response to the peripheral administration of leptin (18), suggesting that thyroid hormone is essential for the leptin effect on food intake. However, one of the limitations of this previous study was that an analysis of the leptin signaling pathway in the isolated appetite-controlling hypothalamic nuclei was not performed.
Therefore, the aims of the present study were to investigate whether the hypophagia associated with hypothyroidism could be related to alterations in protein expression of the anorexigenic and orexigenic ARC neuropeptides, and to explore further the alterations in leptin signaling specifically in the ARC, VMN and PVN in order to understand better the interaction of thyroid hormone and leptin in the regulation of food intake and body weight balance.
Materials and Methods
Animals and experimental design
All experimental protocols were approved by the Committee of Ethical Use of Animals of the Health Sciences Centre of the Federal University of Rio de Janeiro (protocol number 048/14). Three-month-old male Wistar rats (300–350 g) were kept under controlled environment conditions with a 12-hour light–dark cycle (lights on at 07:00am and off at 07:00pm) and room temperature at 23 ± 2°C, with rat chow and water ad libitum. Pharmacological hypothyroidism was induced by the antithyroidal drug methimazole (Sigma-Aldrich) that was administered at 0.03% in the drinking water for 21 days. The body weight of euthyroid and hypothyroid rats was evaluated once a week throughout the treatment. To analyze the acute anorexigenic response to central leptin administration in vivo, a subset of euthyroid and hypothyroid rats underwent stereotaxic surgery and food intake was recorded two hours after an intracerebroventricular (i.c.v.) leptin injection. Another subset of rats was used for protein and mRNA analysis using isolated hypothalamic nuclei. After each experimental protocol, rats were euthanized by decapitation, and trunk blood and brain were collected. Serum was obtained by centrifugation and stored at –20°C for hormonal measurements, and brain tissue was frozen and kept at –80°C.
Stereotaxic surgery and leptin sensitivity test
Euthyroid and hypothyroid rats (300–350 g) were submitted to stereotaxic surgery under anesthesia by intramuscular administration of ketamin (90 mg/kg b.w.) and xylazin (15 mg/kg b.w.), as previously described (19). A stainless steel guide cannula (0.6 mm o.d.; 11.5 mm in length; Acerionx) was implanted into the right lateral cerebral ventricle at 0.8 mm posterior and 1.5 mm lateral from the bregma and 3.4 mm ventral to the skull. The coordinates for cannula implantation were based on a stereotaxic atlas (20), and the correct placement of the cannula was made by observing the displacement in a fluid-filled column (attached to the guide cannula) and further confirmed at the time of slicing the brains on cryostat. A single intramuscular prophylactic dose (48,000 IU) of pentabiotic (Zoetis-Fort Dodge) was given immediately before the surgery. Rats were allowed to recover for five days after surgery until the i.c.v. injections.
After 12 hours of fasting during the dark cycle, leptin (10 μg diluted in 2 μL of sterile saline; Peprotech) was administrated into the right lateral cerebral ventricle of euthyroid and hypothyroid rats. In parallel, another group of euthyroid and hypothyroid rats received 2 μL of saline as controls. The microinjections were carried out using a 2 μL Hamilton syringe (Hamilton Company) connected to the guide cannula through a polyethylene tubing (PE-10). Food intake was evaluated two hours after i.c.v. injections.
Serum hormone measurements
Serum concentrations of T3, thyroxine (T4), thyrotropin (TSH), and leptin were quantified by radioimmunoassay (RIA). TSH serum concentration was quantified using a specific RIA, employing reagents from the National Pituitary Hormone Program (NPHP) as previously described (21), with a limit of detection of 0.18 ng/mL. Total T3 and T4 serum concentration was measured using a commercial kit (MP Biomedicals), according to the manufacturer's recommendations. The limit of detection for the T4 assay was 1 μg/dL, and for the T3 assay it was 25 ng/dL. Serum leptin concentration was measured using a commercial kit (Linco Research), following the manufacturer's recommendation. The limit of detection of the assay was 0.5 ng/mL. For each hormone, all of the measurements were performed in a single run.
Microdissection of hypothalamic nuclei
The ARC, VMN, and PVN nuclei were obtained by the punch technique, as previously described (22 –25), with some adaptations and using the stereotaxic atlas (20). Two consecutive coronal slices of 1.5 mm were made in a cryostat using the bregma as reference. The first slice was taken from 0.6 mm to 2.1 mm posterior to the bregma from which the PVN was dissected in one punch using a round needle 1.5 mm in diameter with the top centered on the third ventricle. The ARC and VMN nuclei were dissected from the second slice using a square needle measuring 2 mm on triangular orientation to isolate the ARC from the base of the brain, and a subsequent punch was performed using the square needle just above the ARC region to dissect the VMN. To control the specificity of the punch technique, the mRNA expression of the steroidogenic factor 1 (Nr5a1) was used as a positive control for microdissection of the VMN. The mRNA expression of Npy was used as a positive control for microdissection of the ARC, and the mRNA expression of thyrotropin-releasing hormone (Trh) was used as a positive control for microdissection of the PVN.
Western blotting
Total protein homogenates of the ARC, VMH, and PVN nuclei were obtained in ice-cold lysis buffer (50 mM HEPES, 1 mM MgCl2, 10 mM EDTA, and 1% Triton X-100, pH 6.4) containing a cocktail of protease inhibitor (Complete, Amersham Biosciences). The total protein content of the homogenates was quantified using the BCA kit (BCA Protein Assay Kit, Thermo Scientific Pierce). Denatured samples were submitted to SDS-PAGE electrophoresis and then transferred to a polyvinylidene membrane (PVDF Hybond-P, Amersham Biosciences). The membranes were then blocked with 5% nonfat milk in Tris-buffered saline (TBS; 20 mM Tris-HCl, 500 mM NaCl, pH 7.6) and incubated overnight with primary antibodies diluted in Tween-TBS (TBS with 0.1% Tween 20) with 2% nonfat milk: anti-ObRb (1:1000), anti-JAK2 (1:500), anti-STAT3 (1:200), anti-pSTAT3 (1:1000), anti-SOCS3 (1:500), anti-POMC (1:1000), and anti-NPY (1:1000). The antibody used for pSTAT3, which recognizes the phosphorylated tyrosine residue Tyr 705, as well as the other antibodies for the proteins of the leptin signaling pathway, were purchased from Santa Cruz Biotechnology; antibodies used for POMC and NPY were purchased from Cell Signaling Technology. Glyceraldehyde 3-phosphate dehydrogenase (GAPDH 1:1000; Cell Signaling Technology) and cyclophilin (1:1000; Applied Biosystems, Thermo Fisher Scientific) were used as loading control. After incubation with the primary antibodies, the membranes were incubated for three hours with the appropriate secondary antibody (1:10,000): anti-mouse Ig-G conjugated to biotin (Santa Cruz Biotechnology) or HRP antirabbit Ig-G (Amersham Biosciences). The membranes incubated with antimouse Ig-G antibodies were then incubated with HRP streptavidin (Invitrogen, Thermo Fisher Scientific) for one hour. The immune reactive proteins were visualized by chemiluminescence by using the ECL kit (ECL Western Blotting System, Amersham Biosciences), and the densitometry analysis of the bands were performed with the Image Quant Las4000 software (GE HealthCare Life Sciences).
mRNA expression analysis
Total RNA of the hypothalamic nuclei ARC, VMN, and PVN was obtained employing the commercial NucleoSpin RNA XS kit (Macherey-Nagel, Inc.), in accordance with the manufacturer's recommendations. This kit is provided with columns containing rDNAse to remove DNA contamination. The concentration of the extracted RNA was quantified in a nanophotometer, and the RNA integrity was confirmed in an agarose gel. RNA samples were stored at −70°C until cDNA synthesis. The reverse transcription was performed employing the Superscript III enzyme (Invitrogen, Thermo Fisher Scientific) and oligo(dT) (Promega) in a final volume of 20 μL. The cDNA was stored at −20°C for future analysis. The mRNA expression was analyzed by real-time polymerase chain reaction (PCR). The cDNA was added to the reaction plate with the mix containing the specific primers, deionized water, and SYBR Green PCR Master Mix buffer (Applied Biosystems, Thermo Fisher Scientific). We performed a minus reverse transcription (RT) reaction (RT–) in all real-time PCR assays, and no amplification product (no Cq value) was detected in any RT– control reaction. The quantification of gene expression levels was performed in the Eppendorf Realplex system. Gapdh was used as reference gene because all samples exhibited similar Cq mean values for this message, without statistical differences between the Cq mean of the control and the hypothyroid group. The quantification of gene expression was performed by the standard curve analysis; each sample was measured in duplicate, and the data are expressed in relation to the euthyroid group. The efficiency of the reactions was calculated by the software Realplex 2 (included in the Eppendorf Realplex 2, MasterCycler), using a cDNA serial dilution, and it varied from 90% to 110%. The same cycling parameters were used for all analyzed messages. Briefly, after an initial denaturation at 50°C for 2 min and at 95°C for 10 min, reactions were cycled 40 times using the following parameters: 95°C for 15 sec, 60°C for 30 sec, and 72°C for 45 sec. The purity of the qPCR was checked by the melting curve added at the end of the cycling reactions, which exhibited a single peak. The PrimeTime Predesigned primers used in this study were intron spanning, and they were obtained from Integrated DNA Technologies (Síntese Biotecnologia):
Lepr—F: 5′-AGG CCC AGA CAT TTT TCC TT-3′/R: 5′-TTC CAA AAG CTC ATC CAA CC-3′; Trh—F: 5′-CAG AAC GCT GAT TCT TGT GG-3/R: 5′-TCC CCT CTC TTC AGC TTC AA-3′; Dio2—F: 5′-TGC TTC AGG ATT GGA CAC G-3′/R: 5′-CTC CTA GAC GCC TAC AAA CAG-3′; Pomc—F: 5′-CCC CGG TTT GAC TGT GTA CT-3′/R: 5′-GTT CCG TCG GTG GTA AAG AA-3′; Npy—F: 5′-AGA GAT CCA GCC CTG AGA CA-3′/R: 5′-TCA CCA CAT GGA AGG GTC TT-3′; Stat3—F: 5′-CTG ACA GTT GGA GAT CAC C-3′/R: 5′-TGT GAT GCC TCC TTG ATT GTC-3′; Nr5a1(SF-1)—F: 5′-CTG TCT TCC TTG CCG TAC TG-3′/R: 5′-CGA AGG TGC ATG GTA TTT AAG G-3′; Gapdh—F: 5′-GTA ACC AGG CGT CCG ATA C-3′/R: 5′-TCT CTG CTC CTC CCT GTT C-3′.
Statistical analysis
Results are expressed as mean ± standard error of the mean (SEM). Statistical differences were determined using one-way analysis of variance followed by the post-test Student Newman Keuls for leptin sensitivity test, or unpaired Student's t-test for protein content and mRNA analysis. A p-value of <0.05 was considered to be statistically significant.
Results
Hormonal profile and Dio2 and Trh mRNA levels
After 21 days of treatment with methimazole (MMI), the total serum concentration of T3 and T4 of hypothyroid rats was reduced by 70% and 50% (p < 0.0001), respectively, compared with euthyroid rats (Fig. 1A and B). The serum TSH of hypothyroid rats was fivefold higher compared with euthyroid rats (p < 0.0001; Fig. 1C). Additionally, to confirm hypothyroidism, the mRNA expression of the type 2 deiodinase (Dio2) and Trh genes were evaluated in the ARC and PVN, respectively. The expression of these two genes is classically increased in hypothyroidism, and hypothyroid rats showed a twofold increase in the expression of Dio2 (p < 0.01) and Trh (p < 0.01) compared with euthyroid rats (Fig. 1D and 1E). These results confirm the efficacy of the protocol in inducing hypothyroidism.
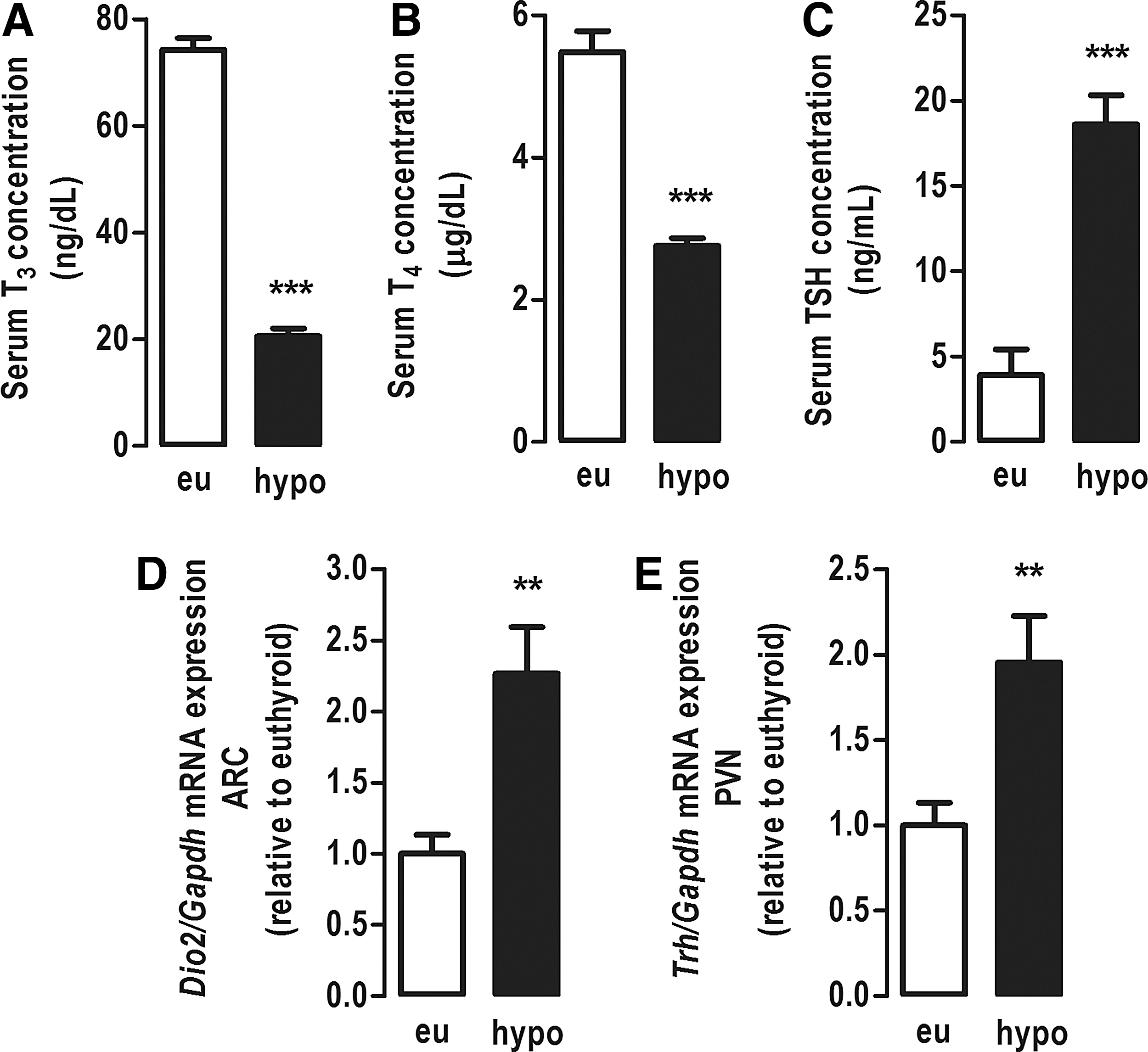
Hormonal profile and Dio2 and Trh mRNA levels. (
Effect of hypothyroidism on body weight, food intake, and ARC neuropeptides
Over the period of treatment with MMI, hypothyroid rats lost weight, and at the end of 21 days of treatment, the mean body weight of the hypothyroid group was significantly lower compared with the euthyroid group (p < 0.0001; Fig. 2A). Hypothyroid rats presented hypophagia with a 50% decrease in food intake compared with euthyroid rats (p < 0.0001; Fig. 2B). Consistent with this hypophagia, the protein content of POMC (Fig. 2C and E) was increased by 70% (p < 0.01), and the protein content of NPY (Fig. 2D and F) was reduced by 50% (p < 0.05) in the ARC of hypothyroid rats compared with euthyroid rats. However, the mRNA expression of Pomc (Fig. 2G) and Npy (Fig. 2H) in the ARC was not affected by the hypothyroidism. Therefore, these data show that the lower body weight and the hypophagia in hypothyroid rats are associated with alteration in the content of appetite-regulating neuropeptides in the arcuate nucleus, without changes in neuropeptide mRNA levels.
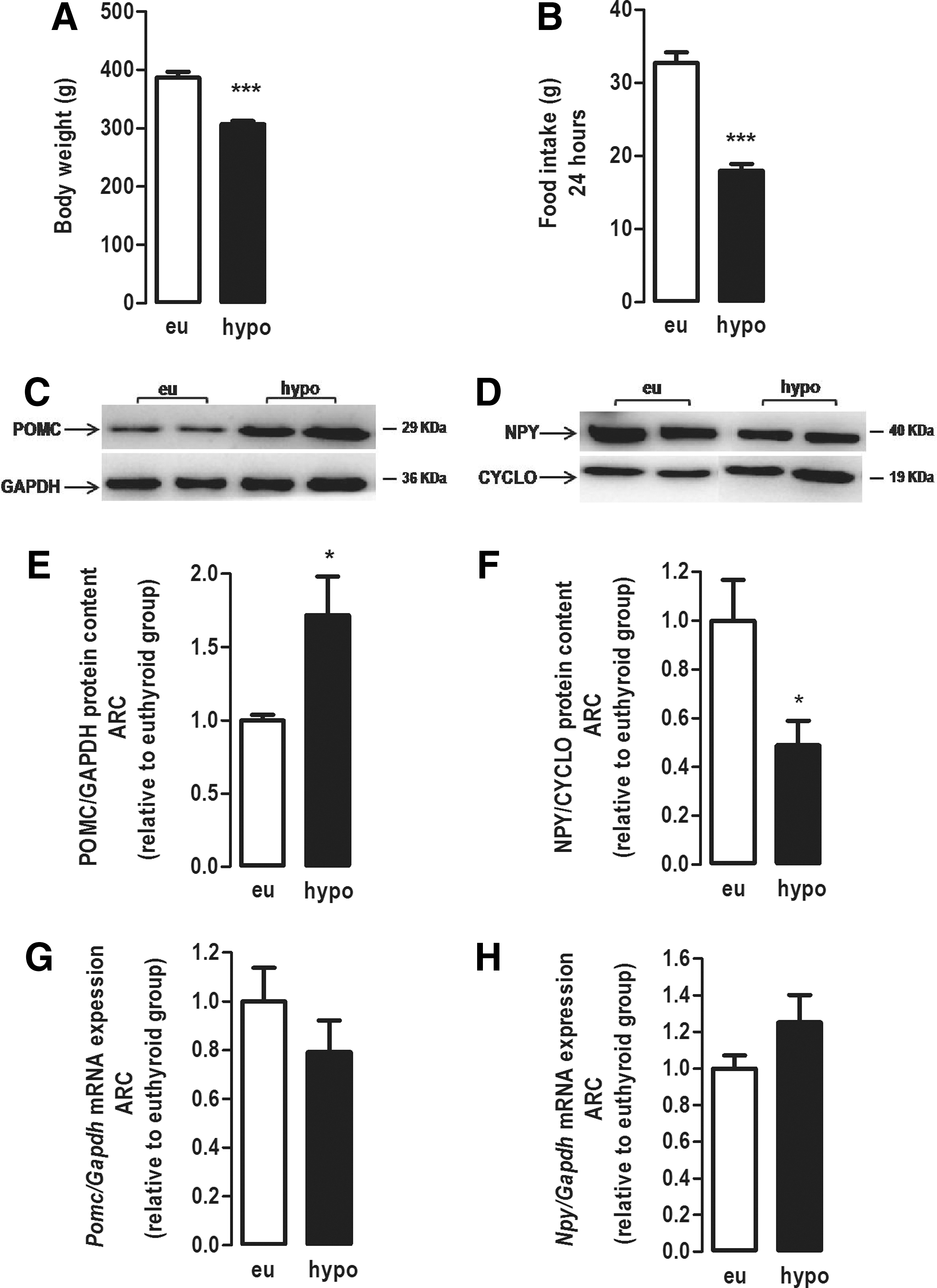
Effect of hypothyroidism on body weight, food intake, and protein expression of POMC and NPY neuropeptides in the ARC of adult rats. (
Effect of hypothyroidism on serum leptin and central leptin sensitivity
The serum concentration of leptin was decreased in hypothyroid rats compared with euthyroid rats (p < 0.05; Fig. 3A). However, serum leptin normalized per body weight was similar between hypothyroid and euthyroid rats (Fig. 3B). The leptin sensitivity test showed that hypothyroidism resulted in impairment of the central anorectic effect of leptin (Fig. 3C). The i.c.v. administration of leptin to euthyroid rats reduced food intake in approximately 50% compared with euthyroid rats treated with saline (p < 0.05; Fig. 3C). Hypothyroid rats treated with saline had a 50% lower food intake than euthyroid rats, confirming their hypophagia (p < 0.05; Fig. 3C), and the i.c.v. administration of leptin was not able to reduce the food intake of hypothyroid rats further (Fig. 3C). These results show that hypothyroid rats are resistant to the central anorectic effect of leptin.
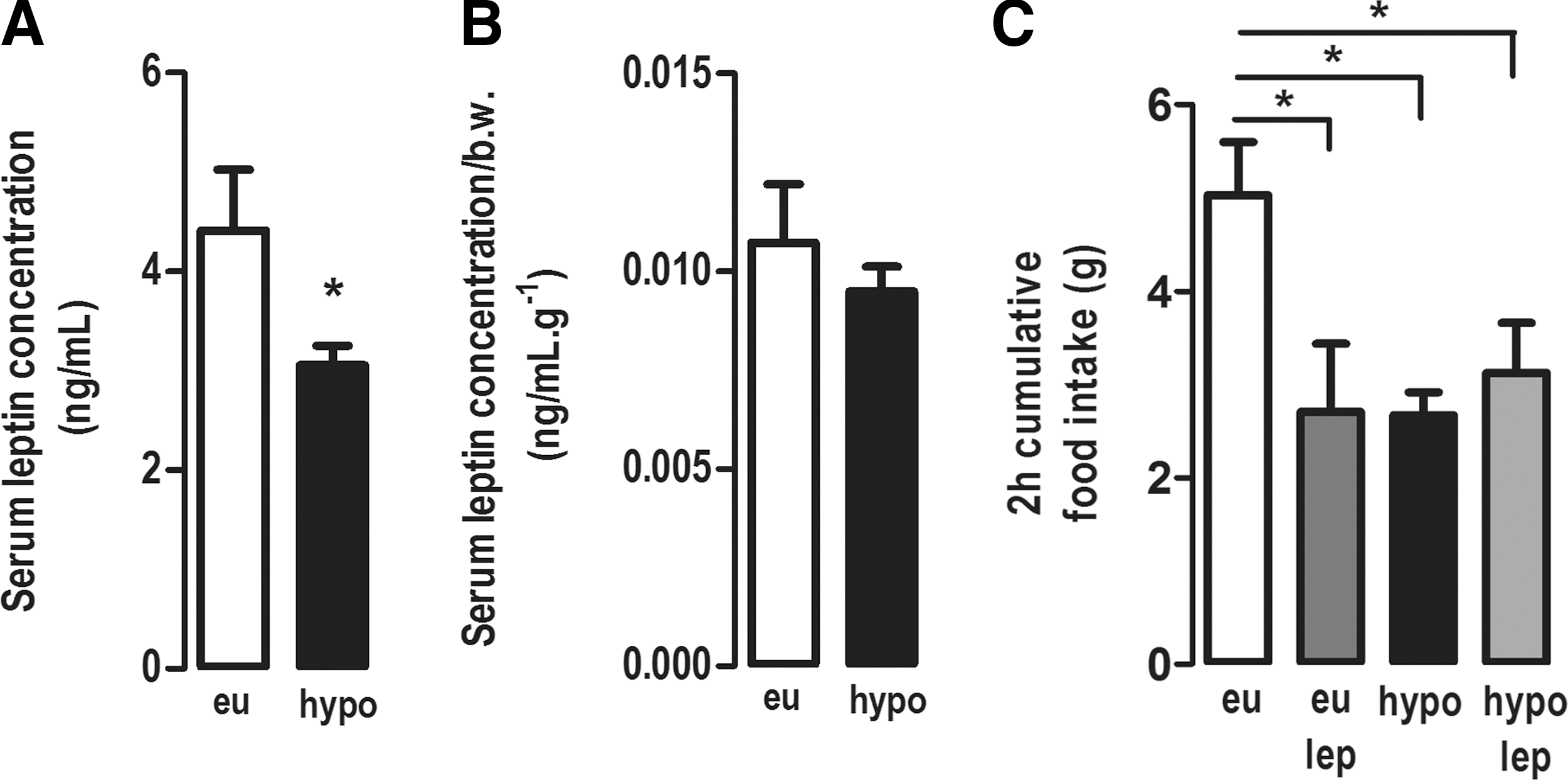
Effect of hypothyroidism on serum leptin concentration and leptin sensitivity in adult rats. (
Effect of hypothyroidism on the leptin signaling in hypothalamic nuclei
In the ARC, hypothyroidism reduced the protein content of ObRb by 40% (p < 0.05; Fig. 4B) and the protein content of pSTAT3 by 30% (p < 0.01; Fig. 4E), with no alterations in the protein content of JAK2, STAT3, and SOCS3 compared with euthyroid rats (Fig. 4C, D, and F, respectively).

Effect of hypothyroidism on leptin signaling pathway in the ARC. (
In the VMN, hypothyroidism reduced the protein content of ObRb by 35% (p < 0.001; Fig. 5B) and the protein content of JAK2 (Fig. 5C) and STAT3 by 30% (Fig. 5D; p < 0.01) compared with euthyroid rats. However, hypothyroidism did not affect the protein content of pSTAT3 (Fig. 5E) and SOCS3 (Fig. 5F).

Effect of hypothyroidism on leptin signaling pathway in the ventromedial nucleus (VMN). (
In the PVN, hypothyroidism reduced the protein content of STAT3 by 35% (Fig. 6D) compared with euthyroid rats (p < 0.01), with no changes in the content of other leptin signaling proteins (Fig. 6B, C, E, and F).
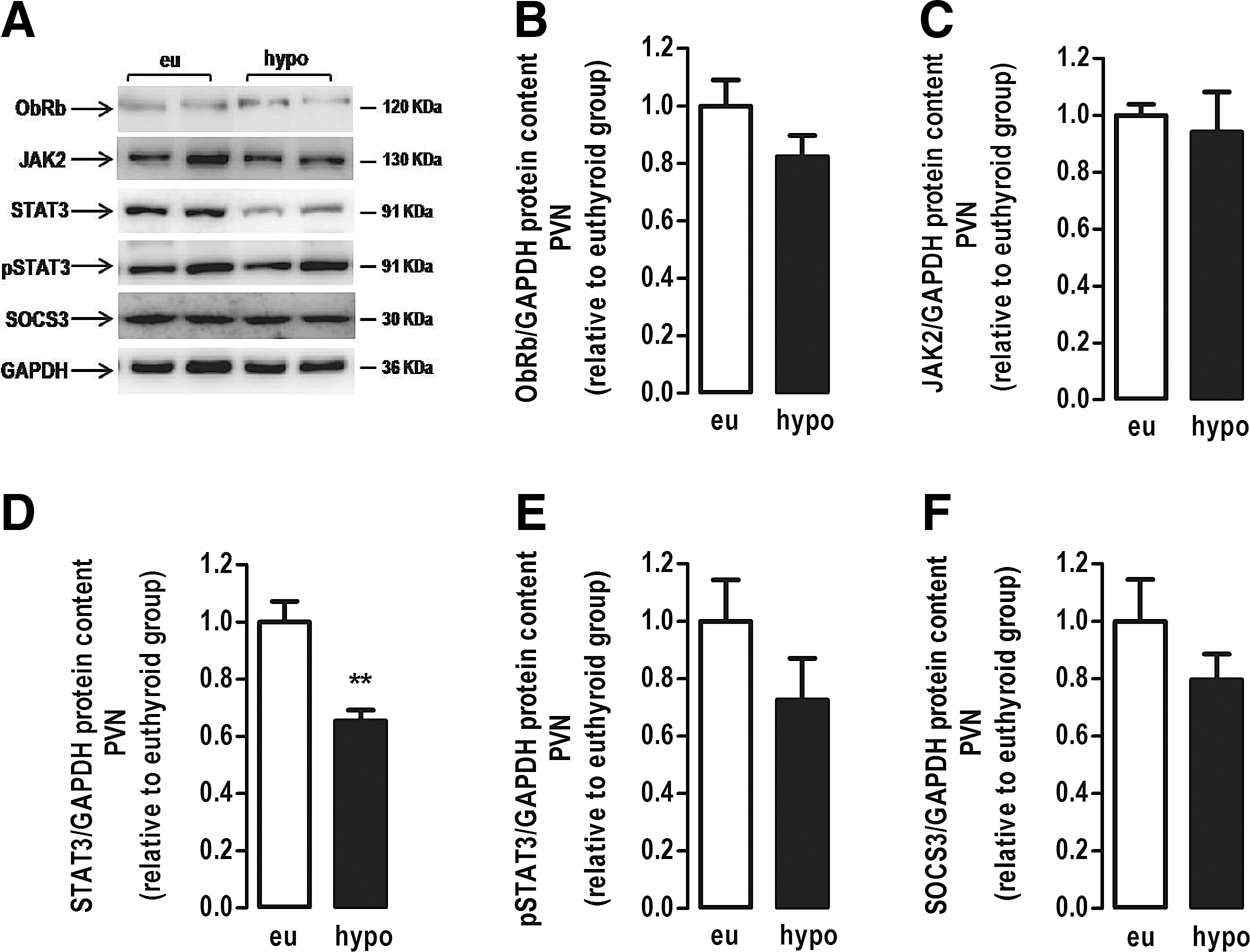
Effect of hypothyroidism on the leptin signaling pathway in the paraventricular nucleus (PVN). (
Hypothyroidism did not alter the mRNA expression of Lepr or Stat3 in the evaluated hypothalamic nuclei (Fig. 7). These data demonstrate that hypothyroidism differentially modulates the content of proteins of the JAK/STAT signaling pathway in the hypothalamic nuclei independently of changes in gene expression.
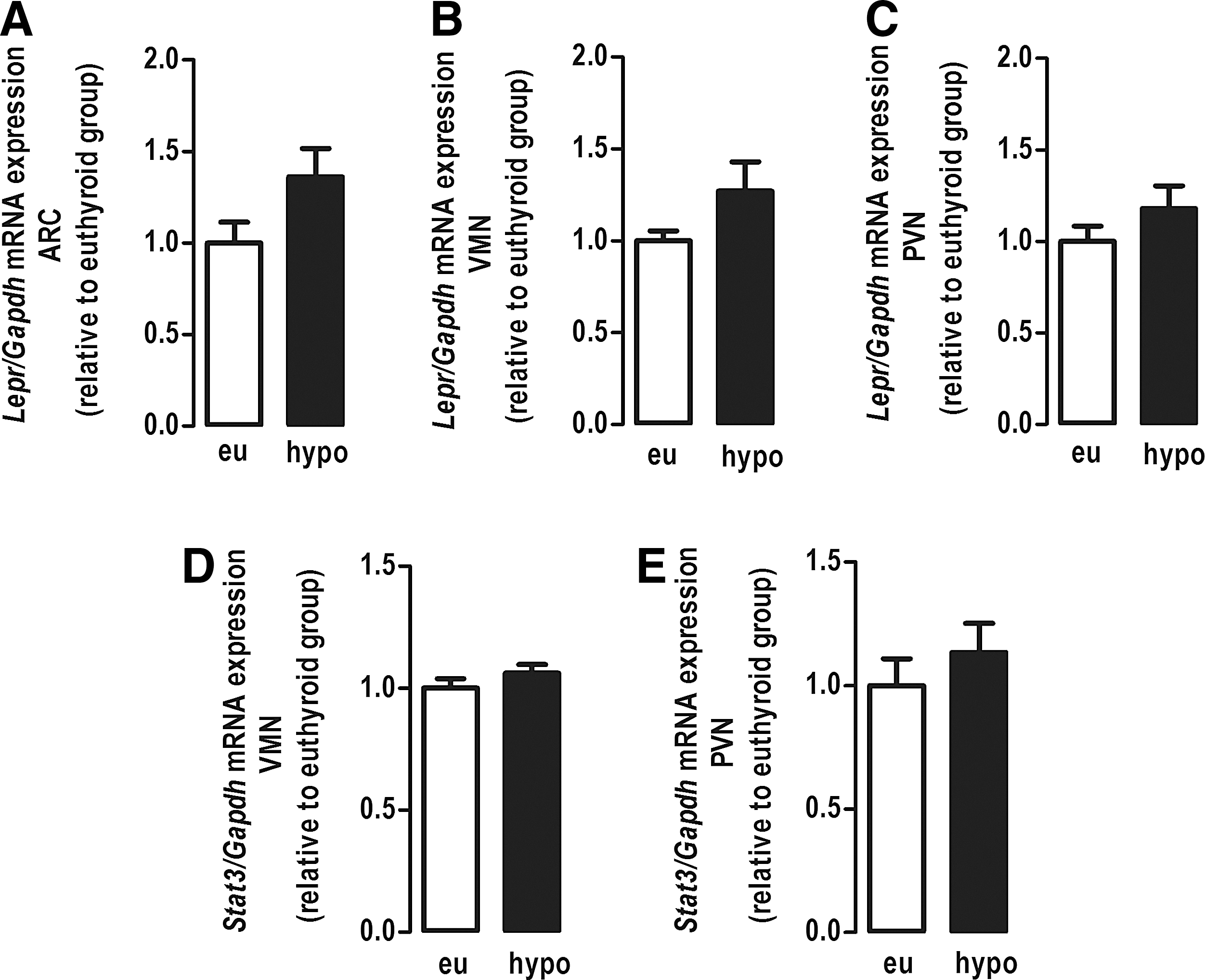
Effect of hypothyroidism on the mRNA expression of the Lepr and the Stat3 genes in hypothalamic nuclei. (
Discussion
In the present study, the primary findings are that hypothyroidism induces hypophagia associated with reduction in the orexigenic peptide NPY, and an increase in the anorexigenic peptide POMC in the ARC, despite central leptin resistance and nuclei-specific changes in leptin signaling. These findings suggest that the effect of thyroid hormone on food intake regulation overrides the leptin effect in hypothyroidism.
Hypothyroidism compromises the body weight gain in rats (3 –5,26 –28), as demonstrated in the present study. Because thyroid hormones have a well-known stimulatory effect on obligatory and adaptive thermogenesis (1), the hypothyroid rats presumably had reduced metabolic rate and energy expenditure. Concomitantly, hypothyroid rats exhibited a pronounced decrease in food intake. Thus, the body weight loss of hypothyroid rats can be explained by the prominent hypophagia, which overrides the decrease in energy expenditure caused by the hypothyroidism. This phenotype may contrast with some reports of weight gain in human hypothyroidism, but the weight gain of hypothyroid patients is mostly due to increasing water retention rather than a consequence of increased adiposity caused by the reduced energy expenditure (29,30).
In accordance with their hypophagia, hypothyroid rats presented increased POMC protein content and reduced NPY protein content in the ARC. Interestingly, the alterations in protein content of these neuropeptides were not accompanied by modifications in their mRNA expressions. To the best of the authors' knowledge, this is the first report to demonstrate that hypophagia of hypothyroid rats may be caused, at least in part, by alterations of the protein content of POMC and NPY in the ARC, independently of changes in their gene expression.
It has been demonstrated that T3 can stimulate food intake by acting directly on the ARC and VMN. However, the data from different studies are controversial. A few studies have shown that hyperthyroidism reduced mRNA expression of Pomc and increased mRNA expression of Npy in the ARC, and these alterations could be the main contributors to the hyperphagia observed in hyperthyroid animals (3,8). However, others have reported that the peripheral and central administration of T3 to euthyroid rats induced hyperphagia concomitant to activation of VMN neurons with no alterations in the expression of POMC and NPY in the ARC (6,7).
Hypophagia of hypothyroid animals is not well understood. Herwig et al. reported decreased Agrp mRNA levels in the ARC of hypothyroid rats without alterations in the mRNA expression of Pomc or Npy (4). In contrast, another study (5) reported that there were no significant changes in Npy mRNA expression in the ARC of hypothyroid mice, but a robust decrease in the expression of Pomc mRNA with increase in mRNA expression of the melanocortin receptor 4 (Mc4r), suggesting a higher sensitivity to the anorexigenic peptide alfa-MSH (5). Collectively, these mRNA data suggest that the melanocortin system is activated in hypothyroidism. The present data support this view, adding that hypothyroidism increased POMC and reduced NPY at the protein level, and therefore, argues in favor of a predominant activation of central anorexigenic pathways in hypothyroid rats. The mechanisms by which hypothyroidism induced alterations in the neuropeptide content may not involve direct TR-mediated gene transcription, since Pomc and Npy mRNA levels were unchanged. A previous study suggests that the hyperthyroidism-induced hyperphagia involves the activation of the hypothalamic mammalian target to rapamycin (mTOR) signaling pathway (8) that is an important pathway to regulate protein synthesis. In addition, the hypothalamic AMP-activated protein kinase (AMPK) is associated with the effects of thyroid hormone on energy metabolism (3). However, even though other nonclassical signaling pathways can be modulated by the thyroid status, to the best of the authors' knowledge, there are no studies showing a direct nongenomic effect of thyroid hormone that could explain the mechanism by which hypothyroidism modulates the protein expression of POMC and NPY in the ARC.
Interestingly, hypothyroid animals are resistant to the acute anorexigenic effect of leptin injected directly in the central nervous system, confirming the same profile we reported when leptin was administered intraperitoneally (18). These observations suggest that leptin resistance is mediated by primary alterations at the hypothalamus and not related to impairment in leptin transport through the blood–brain barrier. Leptin resistance is expected to result in hyperphagia, which is the opposite phenotype shown for hypothyroid rats. Therefore, hypothyroidism induces primary central alterations in appetite-controlling neurons, independently of the anorexigenic effect of leptin.
It has been previously reported that hypothyroid rats exhibited decreased ObRb and phosphorylated STAT3 (pSTAT3) protein levels in total hypothalamus (18). The mechanism by which hypothyroidism induces leptin resistance is further detailed in the present study, and involves modifications of the hypothalamic leptin signaling pathway in a nucleus-specific way. The reduction of the ObRb protein content in the ARC and VMN suggests that hypothyroidism impairs leptin sensitivity in these specific hypothalamic nuclei. In the ARC, this is reinforced by the reduced content of pSTAT3, which is usually an indicative of leptin pathway activity. The present study agrees with that of Groba et al. (31) showing that hypothyroid leptin-deficient ob/ob mice exhibited reduced phosphorylation of STAT3 in the ARC after peripheral leptin administration, suggesting leptin resistance. However, Groba et al. attributed this profile to an increased mRNA expression of the short isoform of the leptin receptor (OBRa; Lepr gene), which would increase leptin internalization, and thus decrease leptin binding to ObRb and JAK-STAT pathway activation. The present study supports the concept that hypothyroidism impairs leptin signaling in the ARC and highlights another possible mechanism by which this might occur, notably the reduction of the protein content of the functional isoform of the leptin receptor, ObRb.
Additionally, the present study demonstrates, for the first time, that the effect of hypothyroidism on leptin signaling extends to the VMN, which was found to have a reduced protein content of ObRb and STAT3, and to the PVN, where hypothyroidism reduced the protein content of STAT3. However, the mechanism by which hypothyroidism interferes with hypothalamic leptin signaling seems not to involve the classical thyroid hormone action via transcriptional regulation (32), since neither the Lepr nor the Stat3 mRNA expression were altered in the hypothalamic nuclei of hypothyroid rats. These data suggest that the alterations observed in the protein content of ObRb and STAT3 were probably due to post-transcriptional modulation (33,34). It has been shown that the ObRb isoform is constitutively endocytosed independently of ligand binding and suffers degradation via the ubiquitin-proteasome system, which regulates its cellular lifetime (35,36). Thus, a possible mechanism, yet to be tested, is that hypothyroidism promotes increased proteasomic degradation of the ObRb in the ARC and VMN resulting in lower levels of this receptor isoform in these particular nuclei.
Hypothyroid rats clearly exhibited a negative energy balance with a pronounced loss of weight and hypophagia. Here, it is suggested that the lack of the central orexigenic effect of T3 might possibly be the primary contributor to the profound hypophagia of hypothyroid rats, since they were unresponsive to the anorexigenic hormone leptin. This is reinforced by the fact that the hypothyroid rats had reduced expression of NPY in the ARC, which is in contrast with situations of negative energy balance, such as fasting, which usually is characterized by stimulation of NPY (37,38). Thus, taking into account that the central leptin action stimulates energy expenditure and inhibits appetite, the hypothalamic resistance to leptin could prevent more dramatic alterations in body weight of hypothyroid rats. Therefore, it is suggested that the loss of the anorectic effect of leptin in hypothyroidism may be an adaptive mechanism to protect the organism.
In conclusion, the present study shows that experimental hypothyroidism induces a negative energetic balance by reducing food intake due to the modulation of hypothalamic protein levels of the main ARC neuropeptides, namely an increase of POMC and a decrease of NPY with consequent predominance of the anorexigenic pathway. In addition, hypothyroidism interferes with leptin signaling in a nucleus-specific fashion in the ARC, VMN, and PVN. The reduction of the leptin signaling in these nuclei may contribute, at least in part, to the central resistance to the anorexigenic effect of leptin in hypothyroid rats, and suggests that leptin is not a determinant factor for the negative energy balance associated with the hypothyroid state. Thyroid hormone and leptin act centrally in a coordinated way to regulate the energy balance, and this coordination includes not only the effects of leptin on the hypothalamus–pituitary–thyroid axis, but also the influences of the thyroid status on the hypothalamic leptin action.
Footnotes
Acknowledgments
All authors are grateful to Luela Luan Dias, Juliana Pena, and Ieda Coleto for their technical assistance. The authors are thankful to Dr. Rosana Torrezan from State University of Maringá for her assistance in the development of the stereotaxic surgery method. The authors are thankful for funding from the National Council for Scientific and Technological Development (Conselho Nacional de Desenvolvimento Científico e Tecnológico—CNPq), the Carlos Chagas Filho Research Foundation of the State of Rio de Janeiro (Fundação Carlos Chagas Filho de Amparo à Pesquisa do Estado do Rio de Janeiro—FAPERJ), and Coordination for the Enhancement of Higher Education Personnel (Coordenação de Aperfeiçoamento de Pessoal de Nível Superior—CAPES).
Author Disclosure Statement
The authors declare that there is no conflict of interest.