
Abstract
Background:
Distant metastases uncommonly occur in differentiated thyroid carcinoma (DTC), but they are a frequent cause of thyroid cancer–related death. Genomic alterations in metastatic tumors, and the relationship with their corresponding primary tumors in DTC, are poorly understood. The objective of this study was to investigate whether genetic alterations in primary tumors are concordant with distant metastases in DTC patients.
Methods:
Surgical samples from primary and matched distant metastatic tumor pairs from 17 DTC patients, and three additional unpaired metastatic tumors from two patients, were analyzed using targeted next-generation sequencing (Ion Torrent Ampliseq cancer panel) with a focus on known recurrent somatic mutations in thyroid cancer. Additionally, TERT promoter mutations were assessed by direct sequencing.
Results:
BRAF mutations were found in 8/10 patients with papillary thyroid carcinoma (PTC). A NRAS mutation was detected in one patient with follicular variant PTC. TERT promoter mutations were detected in 8/10 patients with PTC, and most were coexistent with a BRAF mutation (7/8 BRAF-positive PTC patients, and one BRAF-negative PTC patient). In follicular thyroid carcinoma, NRAS was the most frequently observed mutation (4/9 patients), followed by HRAS (two patients) and KRAS (one patient). TERT promoter mutations were found in 6/7 RAS-positive follicular thyroid carcinoma patients. Key somatic alterations such as BRAF and RAS mutations were highly concordant between primary and matched metastatic tumors without discrepancies. The BRAF or RAS mutant allelic frequency was higher in matched metastatic tumors than in the corresponding primary tumors (35% vs. 25% for BRAF mutation, p = 0.04; and 40% vs. 34% for RAS mutation, p = 0.002). TERT promoter mutations were also mostly concordant in matched tumors (concordance rate 93%).
Conclusions:
BRAF, RAS, and TERT mutations are highly prevalent in metastatic DTC, and are concordant between primary and metastatic DTC. This high concordance suggests that primary tumors may reflect the key somatic alterations in matched metastatic DTC. Frequent coexistent TERT promoter and BRAF or RAS mutations in metastatic DTC also suggests its important role in the progression of DTC.
Introduction
D
Radioactive iodine (RAI) remains the mainstay of treatment for patients with metastatic DTC, but clinical response is variable and influenced by multiple factors. Recent progress in understanding genetic alterations in thyroid cancer has opened the possibility of individualized treatment by targeting relevant mutated genes. In thyroid cancer, this has mainly resulted from the identification of implicated molecular pathways such as the mitogen-activated protein kinase (MAPK) pathway (4,5) and the phosphatidylinositol-3 kinase (PIK3CA)/Akt pathway (6,7). Beyond common mutations such as BRAF or RAS mutations, which are elements of the MAPK pathway, promoter mutations in the telomerase reverse transcriptase (TERT) gene have recently been identified in follicular cell–derived thyroid cancer (8,9). Recent studies have demonstrated that TERT promoter mutations were strongly correlated with aggressive clinicopathologic characteristics of thyroid cancer (9 –13).
In patients with metastatic cancer, genetic alterations are often assessed in archived primary tumor specimens. However, metastatic thyroid tumors may differ from primary tumors at the molecular level, and clonal relationships between primary tumors and distant metastases are still not understood in DTC.
The advent of next-generation sequencing (NGS) technologies has revolutionized cancer genomics research (14). NGS consists of high-throughput DNA sequencing technologies that are capable of sequencing large numbers of different DNA templates in parallel. NGS has clear advantages over conventional Sanger sequencing, by providing a simultaneous analysis of large regions of the genome with increased sensitivity and speed (15).
The aim of this study was to conduct an NGS-based molecular analysis of DTC to evaluate whether genetic alterations in primary DTC are concordant with those in metastatic DTC. The study also aimed to investigate the frequency of TERT promoter mutations in metastatic DTC.
Methods
Identification of patient samples
Paraffin blocks from formalin-fixed primary DTC and metastatic tumors were obtained at Samsung Medical Center (Seoul, Korea). All histologic diagnoses were evaluated by two pathologists (Y.Y. Jung and Y.L. Oh) at the authors' institution. Patients were selected on the basis of sample availability. Nineteen DTC patients (10 with PTC, 9 with FTC) with pathologically proven distant metastases or recurrence after surgery from January 2003 to December 2012 were retrospectively identified. Clinicopathologic data were retrospectively reviewed. The formalin-fixed paraffin-embedded (FFPE) tumor tissue blocks included 17 primary thyroid tumors and 23 distant metastatic tumors or recurrent tumors at the following sites: lung (n = 11), bone (n = 6), brain (n = 3), kidney (n = 1), and cervical lymph nodes (n = 2). This study was approved by the Institutional Review Board committee of Samsung Medical Center.
DNA isolation
For FFPE tissues, tumor-rich areas (>80% of tumor cells) were dissected from unstained sections of 1–4 μm thickness under microscopy using a hematoxylin and eosin–stained slide for guidance. DNA was extracted using the QIAamp DNA FFPE Tissue Kits (Qiagen, Valencia, CA).
NGS
The Ion Ampliseq Cancer Panel v2 (Life Technologies, Foster City, CA) was used to generate target amplicon libraries. DNA (10 ng) derived from FFPE tissue was amplified by polymerase chain reaction (PCR) using the Ion AmpliSeq Cancer Panel Primer Pool and Ion AmpliSeq HiFi Master Mix according to the manufacturer's protocols.
The multiplexed 154-amplicon library targets 50 cancer genes (Table 1), including 2800 COSMIC mutations. For library construction, an average of 154 multiplexed amplicons were treated with a FuPa reagent to digest primer sequences partially and phosphorylate the amplicons. The amplicons were then ligated to adapters from the Ion Xpress barcoded adapters 1–16 kit according to the manufacturer's instructions (Ion AmpliSeq Library Kit 2.0). After ligation, the amplicons underwent nick translation and additional library amplification by PCR to complete the linkage between adapters and amplicons. Library concentration and amplicon size were determined using an Agilent BioAnalyzer high sensitivity DNA kit (Agilent Technologies, Santa Clara, CA). Next, multiplexed barcoded libraries were enriched by clonal amplification using emulsion PCR on Ion Sphere particles (IonPGMTemplate OT2 200 kit) and loaded on an Ion 318 Chip. Massive parallel sequencing was performed on Ion 316 Chips using the Ion PGM Sequencing 200 Kit version 2 according to the manufacturer's instructions.
Detection of TERT promoter mutations
The TERT promoter mutations were analyzed by PCR amplification and direct Sanger sequencing of the hot spots, as previously described (chr5:1,295,228C>T and chr5:1,295,250C>T, commonly termed C228T and C250T) (9). Briefly, a 235 bp region of the TERT promoter containing the hot spots of C228T and C250T mutations was amplified by PCR. After gel electrophoresis to confirm the quality of PCR products, sequencing of the PCR-generated templates was performed using a GeneAmp PCR system 9700 thermal cycler (Applied Biosystems, Foster City, CA), and the DNA sequence was analyzed on an ABI 3730xl Genetic Analyzer (Applied Biosystems).
Screening for RET/PTC and PAX8/PPARγ rearrangements
RNA was extracted from FFPE tissue from the corresponding areas from which DNA was extracted, using a High Pure RNA Paraffin Kit (Roche, Mannheim, Germany). Real-time PCR assays (RT-rPCR) were performed to detect RET/PTC1, RET/PTC3, and PAX8/PPARγ rearrangements. RT-rPCR thermal cycling for RNA amplification and data acquisition were performed with an Applied Biosystems 7500 Real-Time PCR system (Life Technologies). FAM fluorescence of KRT7 was employed as an internal control to confirm appropriate nucleic acid isolation from thyroid follicular cells and exclude possible PCR inhibition.
Bioinformatics (NGS) analysis
Data analysis was performed using the Torrent Suite v3.6.0 and Ion Torrent Variant Caller v3.6 software. The pipeline includes signaling processing, base calling, quality score assignment, adapter trimming, read alignment to 19 human genome reference points, mapping quality QC, coverage analysis, and variant calling. Variant calls were further analyzed using ANNOVAR to provide variant filtering and annotation using the COSMIC database and dbSNP build 137.
In order to eliminate erroneous base calling, several filtering steps were used to generate the final variant calling. First, known non-pathogenic polymorphisms or common germline variants were filtered by searching the dbSNP database. The second filter consisted of visually examining mutations using Integrative Genomics Viewer (IGV) software (
Among the detected mutations, the focus was on those mutations annotated in the COSMIC database or in the literature. Known recurrent somatic alterations were defined as genetic alterations in genes reported to be mutated with a frequency ≥5% in the COSMIC database or in the literature (9,16 –18). Other mutations were considered as likely passenger mutations.
Statistical analysis
A paired t-test was used to compare allelic frequency of primary tumors and metastatic tumors. Because only 10 PTC and 9 FTC patients were included in this study, it was not possible to perform statistical analysis between clinical outcome and mutation status. All statistical analyses were performed using SPSS Statistics for Windows v21.0 (IBM Corp., Armonk, NY). p-Values of <0.05 were considered significant.
Results
Patients and sample characteristics
The clinicopathologic characteristics of the 19 patients are presented in Table 2. The mean age at the time of diagnosis was 59.8 ± 13.8 years, and females predominated (70%). The median stimulated serum thyroglobulin (Tg) level at initial ablation was 227 ng/mL (interquartile range 33–3246 ng/mL). All patients received RAI therapy, with a mean frequency of three treatments and a median cumulative dose of 450 mCi (interquartile range 290–680 mCi). Distant metastases were detected at the time of diagnosis in 10 patients. None of the patients received targeted therapy before genomic analysis. Median follow-up was 64 months (range 20–132 months).
cPTC, conventional papillary thyroid carcinoma; FV-PTC, follicular variant papillary thyroid carcinoma; FTC, follicular thyroid carcinoma; sTg, stimulated thyroglobulin; RAI, radioactive iodine.
Forty thyroid tumors from the 19 patients were analyzed for genetic analysis, which included 17 primary thyroid tumors and 23 distant metastatic lesions or metastatic lymph nodes (lung, n = 11; bone, n = 6; brain, n = 3; kidney, n = 1; and lymph node, n = 2). Primary tumor samples and metastatic or recurrent samples were available from 17/19 patients; of these, eight patients yielded samples of both synchronous primary tumors and metastases (including one patient with both synchronous and metachronous metastases), while the other nine patients yielded samples of primary tumors and of subsequent metastatic tumors. In addition, three metastatic samples (two of which were from one patient) without matched primary tumors were analyzed, because little is known about the spectrum of genomic alterations in patients with metastatic thyroid cancer.
Genetic alterations of PTC and comparison between primary and metastatic tumors
A total of nine primary tumors, 10 distant metastatic tumors, and two lymph nodes were analyzed for genotyping. Among 10 patients with PTC, nine were diagnosed with classic PTC and one with follicular variant PTC (FV-PTC). Oncogenic mutations were found in all nine primary tumors and 11/12 metastatic tumors (Table 3 and Supplementary Table S1; Supplementary Data are available online at
NA, not applicable due to absence of tumor; PTC, papillary thyroid carcinoma.
To investigate molecular evolution, genomic alterations in matched primary and metastatic tumors from nine PTC patients were compared (Table 3). Two metastatic tumor samples were available from two patients (patients 8 and 9). Known recurrent somatic alterations in thyroid cancer, such as BRAF and RAS mutations, were highly concordant between primary and metastatic tumors. No discrepancies in BRAF or NRAS mutations were observed between primary tumors and corresponding metastases. Among the eight patients harboring TERT promoter mutations, one PTC patient showed a discordant TERT mutation between the primary and the metastatic tumor; the TERT promoter mutation was detected only in the primary tumor. Two patients had the same BRAF mutations in their primary and two subsequent metastatic tumors (Fig. 1A). They harbored coexisting TERT promoter C228T mutations in the primary tumor and two corresponding metastases (Fig. 1B). In contrast, likely somatic mutations were discordant between primary and metastatic tumors, and these were coexistent with BRAF or RAS mutations.
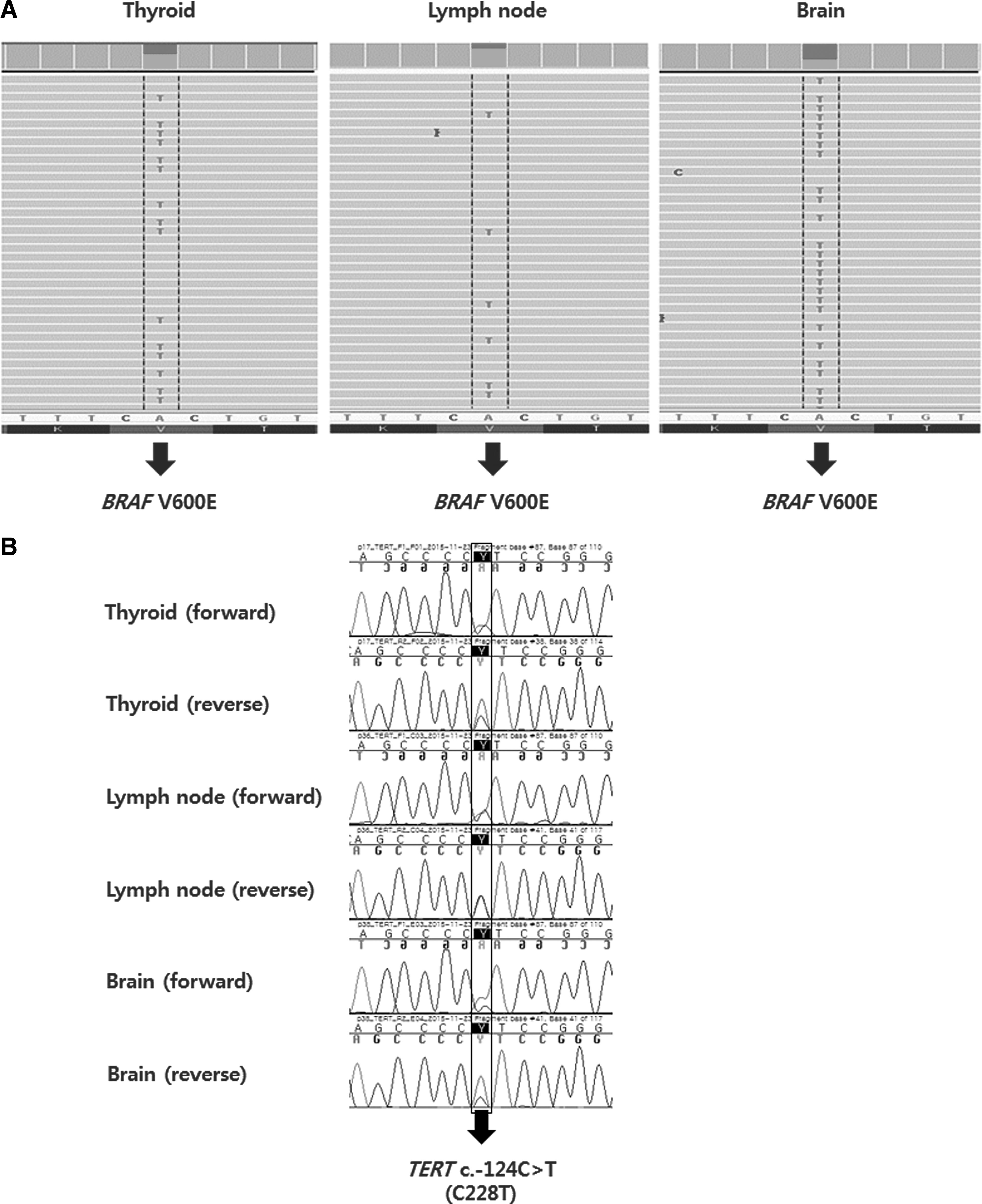
(
Genetic alterations of FTC and comparison between primary and metastatic tumors
Eight primary tumors and 11 metastatic tumors were analyzed. Seven of the eight primary tumors and 9/11 metastatic tumors harbored at least one alteration (Table 4 and Supplementary Table S1). The most common alterations were RAS mutations in both primary and metastatic FTC. These included six RAS mutations (three NRAS, two HRAS, and one KRAS) in primary tumors, and eight RAS mutations (five NRAS, two HRAS, and one KRAS) in metastatic tumors. All of these RAS mutations were found in codon 61. A TERT promoter mutation was frequently detected in tumors harboring a RAS mutation, that is, in 5/6 RAS-positive primary tumors and 7/8 RAS-positive metastatic tumors, respectively. All of the TERT promoter mutations were C228T. Three primary FTC and six metastatic FTC exhibited genetic alterations other than RAS mutations. These included ALK (n = 2), APC (n = 1), CDH1 (n = 2), RB1 (n = 3), RET (n = 4), PTEN (n = 1), and SMAD4 (n = 2). The known recurrent PTEN p.V166I mutation was coexistent with NRAS mutations. PAX8/PPARγ rearrangements were not detected in primary or metastatic FTC.
NA, not applicable due to absence of tumor.
Genomic alterations in matched primary and metastatic tumors from nine patients, and two matched metastatic tumors from one patient, were compared (Table 4). RAS and TERT promoter mutations were highly concordant between primary and metastatic tumors, without discrepancy. The concordance rate for five known recurrent mutations in FTC (HRAS, KRAS, NRAS, PTEN, and TERT promoter) was 93%, whereas for likely passenger mutations, it was 17%.
Mutation allelic frequency
The NGS technique allows quantitative assessment of mutation frequency in addition to the detection of mutations. The allelic frequency of BRAF and RAS mutations is summarized in Table 5. In PTC, the allele frequency of BRAF mutation was 9–2% (M = 25%) in primary tumors, corresponding to 18–84% of cells with heterozygous mutations. The allele frequency of BRAF mutations was higher in matched metastatic tumors, with a frequency of 22–49% (M = 37%, p = 0.04). In FV-PTC and FTC, the allele frequency of RAS mutation was 14–42% in primary tumors (M = 34%), and 21–48% in matched metastatic tumors (M = 40%, p = 0.002). Likely passenger mutations had relatively lower allele frequencies than did BRAF or RAS mutations (data shown in Supplementary Table S1).
DTC, differentiated thyroid carcinoma; NA, not applicable due to absence of tumor.
Clinical outcome
The clinical outcome of enrolled patients is summarized in Table 6. No patient achieved complete remission. Of the 19 enrolled patients, one PTC patient with BRAF and TERT promoter mutations and two FTC patients with NRAS and TERT promoter mutations died from disease progression. Median progression-free survival after metastasectomy was 36 months. FTC patients had clinical outcomes indicating more aggressive disease than that in PTC patients. Among the nine FTC patients, six presented progressive disease or death at last follow-up. Four of 10 PTC patients showed progressive disease or death during follow-up, and all had both a BRAF and a TERT promoter mutation in the metastatic tumors.
Progression-free survival after metasectomy.
Detected only in primary tumor, not in metastatic tumor.
PD, progressive disease; SD, stable disease.
Discussion
An NGS-based molecular analysis of metastatic DTC was conducted and alterations therein were compared with those in primary tumors. To the best of the authors' knowledge, this is the largest study of targeted NGS analysis of primary tumors and matched distant metastases in DTC patients. Key somatic mutations such as BRAF or RAS mutations were highly concordant between primary and metastatic tumors, with higher allelic frequency than likely passenger mutations. Coexistent TERT promoter mutations were frequently found in tumors harboring BRAF or RAS mutations and were mostly concordant in matched tumor samples.
Distant metastasis is generally related to cancer-related death. Therefore, identifying genetic alterations in metastases may be of great importance in therapeutic decision making. Metastatic progression is considered a multistep process involving a succession of acquired genetic and epigenetic events in tumor cells (19). These acquired genetic changes are regarded as a result of selection during the metastatic process (20). Thus, theoretically, it can be assumed that there is molecular discrepancy between primary and metastatic tumors. A recent study of a single kidney cancer highlighted the heterogeneity not only between primary and metastatic tumors, but also between different metastatic sites and even in different regions of the same tumor (20). Despite this seeming divergence, the present study showed a high degree of concordance of key somatic alterations in DTC. The results are in line with previous reports that found close clonal relationships between primary tumors and secondary metastatic sites in cancers of other sites (21 –23). A recent pooled literature analysis of colorectal cancer reported high concordance rates of 92% for KRAS and 96.8% for BRAF mutations (21). Other NGS-based studies also found a high concordance of key somatic alterations between primary and metastatic tumors in patients with non-small cell lung cancer (23) and in breast cancer (22). In thyroid cancer, a study by Julio et al. that included 12 patients revealed a high concordance of BRAF (7/8 patients) or RAS (only in one patient) mutations between matched tumor samples (24). Other studies also showed close clonal relationships between primary and metastatic tumors in thyroid cancer. However, the majority of matched analyses were evaluated in lymph nodes, and the number of matched cases in distant metastasis was limited. (25 –27). In the 17 patients in the present study, nine patients with classic PTC had BRAF mutations, and eight patients (one with FV-PTC and seven FTC patients) had RAS mutations, and the BRAF or RAS mutation found in the primary tumor was also present in all distant metastases. Taken together, mutations activating the MAPK pathway, such as BRAF and RAS, were highly concordant between matched thyroid tumors, suggesting that primary tumors may reflect the key somatic mutations in matched metastatic DTC.
This study also showed mutual exclusivity of key somatic alterations such as BRAF, RAS, and RET/PTC in PTC, findings that are consistent with some previous studies (4). Mutual exclusivity of key somatic mutations was also preserved in metastatic lesions. In contrast, other studies found concomitant mutations in BRAF or RAS and other mutations, and association thereof with poor clinical outcome (19,20). These results illustrated the biological relevance of subclonality of BRAF mutations. However, strong tumorigenicity of BRAF was well demonstrated in a cell line (28,29) and in a transgenic mouse model (30,31). In the transgenic mouse model, development and progression of PTC could be initiated by expression of BRAF alone. The quantitative assessment of BRAF mutant alleles also supports that BRAF is a clonal driver mutation in PTC. The current study found that the mean allelic frequency of BRAF was 25% (50% of cells with heterozygous mutation) in primary PTC, and 38% (76% of cells with heterozygous mutation) in metastatic PTC. In contrast to the current findings, Gandolfi et al. reported that the average mutated allele percentage of BRAF was significantly lower in corresponding lymph node metastases (LNM) than in the primary tumors (18.6% vs. 28.6%) (25). Guerra et al., using a pyrosequencing approach, reported that BRAF mutations were detected only in a small subset of the tumor cells: 66% of PTCs with a BRAF mutation had a mutated allele frequency of only <25% (32). Furthermore, de novo mutations of BRAF in LNM, with BRAF wild type in the primary tumor, was observed, albeit rarely (26). These findings suggest that the occurrence of a BRAF mutation may be a later subclonal event in PTC. However, a pyrosequencing approach might underestimate the frequency of mutated alleles compared with NGS. Using an NGS approach, Nikiforova et al. observed that the allelic frequency of BRAF in a set of 19 PTC samples ranged from 18% to 44% of total alleles, and the majority of them had >25% mutated alleles (i.e., >50% of cells with a heterozygous mutation) (18). Because tumor samples have some degree of contamination with non-tumoral cells, the presence of BRAF mutations in ≥50% of cells within the target tumor regions supports the conclusion that it is a clonal driver mutation. Recently, the Cancer Genome Atlas (TCGA) study of 496 PTCs found that BRAF mutations were present in the majority of tumor cells, strongly suggesting that BRAF mutations occur as a clonal event in thyroid cancer (16). In addition, LNMs might not represent progression to a more aggressive status, as in the case of distant metastasis, in PTC. Therefore, the mutant allele fraction in LNM might be different compared with distant metastasis. The present finding of a higher mutated allele frequency in metastatic tumors suggests a clonal expansion of tumor cells harboring BRAF mutations during tumor progression. Taken together, these results support that a BRAF mutation is a clonal driver mutation in PTC tumorigenesis and can propagate to the metastases developed during tumor progression.
The present analysis of FTC found a high prevalence of RAS mutations, and all of these RAS mutations were located in codon 61. The most frequent RAS mutation was the NRAS codon 61 mutation (44%; 4/9), followed by HRAS codon 61 mutations (22%; 2/9) and KRAS codon 61 mutations (11%; 1/9), which is similar to previous reports (33 –35). Previous studies reported that the presence of any RAS mutation was associated with poor clinical outcomes (33), and that the NRAS codon 61 mutation was particularly related to distant metastases (33,35). RAS mutations seem not only to be involved in early tumorigenesis, but also in FTC progression, according to the results of previous studies (36). The presence of this mutation in benign follicular adenomas (FA), and a higher rate of mutations in follicular-patterned carcinomas (33,37), suggests that RAS mutations are an early event in follicular thyroid carcinogenesis. Furthermore, RAS mutations may predispose well-differentiated cancers to dedifferentiation (36,38). In the present analysis of RAS mutant allele frequency, RAS mutations had an average allelic frequency of 34% (range 14–42%) in primary tumors and 39% (range 21–48%) in metastatic tumors. In RAS mutant FTC tumors, all primary and metastatic tumors had >25% RAS mutant alleles (>50% of cells carrying RAS heterozygous mutations), and matched metastatic tumors had a mean 42% of RAS mutant alleles (>84% of cells harboring RAS heterozygous mutation). Likely, passenger mutations other than RAS had relatively low allelic frequencies. The recent TCGA study revealed that all RAS mutations were present in the majority of cells within tumors like other driving mutations such as BRAF, indicating that mutated RAS is a clonal event in thyroid cancer (16). The present finding of a relatively high-mutated allele frequency in metastatic tumors also suggests a clonal expansion of tumor cells harboring RAS mutations during tumor progression.
Beyond more common driver mutations such as BRAF or RAS mutations in thyroid cancer, this study showed that TERT promoter mutations were highly prevalent in metastatic DTC. The prevalence of TERT promoter mutations in PTC was 80% (8/10) and 67% (6/9) in FTC. As only patients with distant metastasis were included, these findings are in line with previous reports of a particularly high prevalence of TERT promoter mutations in follicular cell-derived thyroid cancer with higher tumor stage or aggressive clinical behavior (10 –13). Furthermore, these findings show that TERT promoter mutations were frequently detected in tumors harboring BRAF or RAS mutations. Recent studies have addressed the association between TERT promoter and BRAF mutations (10,13). Coexisting TERT promoter and BRAF mutations were more commonly and significantly associated with clinicopathologic aggressiveness than were the presence of a mutation in a single gene (10,38), suggesting a functional cooperative effect of these mutations. Two somatic TERT promoter mutations, chr5:1,295,228C>T and chr5:1,295,250C>T (termed C228T and C250T, respectively), were found to generate a binding site for ETS transcription factors (39,40). As activation of the MAPK pathway upregulates the ETS system (41), MAPK activation through either BRAF or RAS mutations promotes the upregulation of the TERT gene expression through the interaction of ETS factors with the TERT promoter. A highly concordant TERT promoter mutation status was also found in matched tumors. De novo occurrence of a TERT promoter mutation in metastatic tumors was not found in the present cases. Unexpectedly, a TERT promoter mutation was detected only in the primary tumor in one patient. This finding cannot be explained, but the different cellular composition of the metastatic tumor may influence this observation, despite careful selection of tissues to minimize a bias. An Italian study involving 16 matched LNM originating from 15 primary PTCs also showed a highly concordant TERT mutation status, although de novo occurrence of a TERT mutation in the lymph nodes in a single patient with a primary tumor without TERT mutation has been detected (11). A recent study using NGS analysis found that the mutant allele frequency of TERT mutations was higher in LNM than in primary tumors (42), suggesting that TERT promoter mutations are late events during progression. However, matched analysis was performed in only one patient, and therefore a further large study involving quantitative analysis is warranted.
This study has some limitations. First, patients were selected based on sample availability, and multivariate regression analysis could not be performed to assess associations between prognostic factors, including genetic alterations and clinical outcomes. However, it has been challenging to find matched sets of primary and metastatic thyroid tumors because biopsy of metastatic diseases is not routinely performed in thyroid cancer management. Second, the focus here was on known recurrent somatic mutations, and these were considered as key oncogenic events in DTC. The possibility cannot be excluded that some alterations currently considered as likely passenger mutations because of their unknown significance in thyroid cancer may actually also exert a driver function in some specific molecular subtypes. Third, an analysis of gene expression and some important fusion genes such as ALK, BRAF, and NTRK rearrangements (16) was not performed because of the limitation of the Ampliseq Cancer panel. Fourth, this study was conducted in Korea, which is an iodine-rich area with a higher prevalence of BRAF mutations than most countries (39). However, this should have little impact on the findings of highly concordant key genetic alterations between primary and metastatic DTC.
In conclusion, the present findings from a subset of DTC tumors reveal a high level of concordance in key somatic alterations. Therefore, genetic alterations in primary tumors can reflect the genomic spectrum of the patients' metastatic disease. The relatively high mutant allele frequency of BRAF or RAS mutations in metastatic tumors suggests clonal expansion of tumors harboring these mutations during progression of DTC. Coexistence of a TERT promoter mutation with a BRAF or RAS mutation was commonly found in metastatic DTC, suggesting their co-operative role in the progression of DTC.
Footnotes
Acknowledgments
This study was supported by the Korean Foundation for Cancer Research (#CB-2011-03-02).
Author Disclosure Statement
The authors have no conflicts of interest to declare.