
Abstract
Background:
The diaphragm is the main respiratory muscle, and its function is compromised during severe illness. Altered local thyroid hormone (TH) metabolism may be a determinant of impaired muscle function during illness.
Methods:
This study investigates the effects of bacterial sepsis and chronic inflammation on muscle fiber type, local TH metabolism, and mitochondrial function in the diaphragm. Two mouse models were used: sepsis induced by S. pneumoniae infection or chronic inflammation induced by subcutaneous turpentine injection. In vitro, the effect of bacterial endotoxin (LPS) on mitochondrial function in C2C12 myotubes was studied.
Results:
Sepsis induced a transient increase in the fiber type I profile and increased Dio3 expression while decreasing Dio2, Thra1, and Slc16a2 expression. Triiodothyronine positively regulated genes Tnni2 and Myog were decreased, indicating reduced TH signaling in the diaphragm. In contrast, chronic inflammation increased the fiber type II profile in the diaphragm as well as Thra1, Thrb1, and Myog expression while decreasing Dio3 expression, suggesting increased TH responsiveness during chronic inflammation. LPS-stimulated C2C12 myotubes showed decreased Dio2 expression and reduced basal oxygen consumption as well as non-mitochondrial respiration. The same respiratory profile was induced by Dio2 knockdown in myotubes.
Conclusions:
The in vivo results show differential effects of sepsis and chronic inflammation on diaphragm muscle fiber type, TH metabolism, and mitochondrial function, while the in vitro results point to a causal role for altered TH metabolism in functional muscle impairment. These findings may be relevant for the pathogenesis of impaired respiratory function in critical illness.
Introduction
I
Recent studies indicate that in addition to altered TH plasma levels, illness also changes local tissue TH metabolism (4). TH metabolism at the cellular level is regulated mainly by the activity of enzymes that control the production and degradation of TH by deiodination. There are three types of deiodinating enzymes: types 1, 2, and 3 (D1, D2, and D3, respectively) (5). D2 converts T4 into T3, thereby increasing the intracellular level of the active hormone, while D3 decreases intracellular T3 by converting T4 into reverse T3, or T3 into T2. The interplay between D2 and D3 controls the intracellular bioavailability of T3 (6).
Skeletal muscle is an important T3 target tissue, and expresses it both D2 and D3. Muscle Dio2 mRNA and activity are increased in critically ill patients admitted to the intensive care unit compared with healthy controls (7). In contrast, muscle Dio2 expression is decreased, while muscle Dio3 expression is increased in patients with septic shock (8). In mice, severe Streptococcus pneumoniae infection decreases muscle Dio2 expression while Dio3 remains unchanged. Moreover, chronic inflammation induces both Dio2 and Dio3 in skeletal muscle (9). Thus, alterations in muscle deiodinase expression depend on the severity of illness and the inflammatory response (3).
Respiratory muscles of critically ill patients are susceptible to functional impairment, often leading to prolonged dependence on mechanical ventilation in order to maintain appropriate respiratory gas exchange (10,11). The diaphragm is the main respiratory muscle comprising type I slow-twitch fibers and type II fast-twitch fibers. Although the mechanism involved in respiratory muscle failure during sepsis is not well understood, it probably includes decreased mitochondrial function (10,12). Mitochondria are the main sources of cellular energy and, therefore, decisive for muscle performance. Of note, TH plays a central role in the control of mitochondrial biogenesis and activity (13). Moreover, T3 is an important regulator of muscle fiber type and contractility (14). As the diaphragm expresses Dio2 and Dio3 as well as T3 receptors, changes in diaphragm TH metabolism might be an important determinant in the pathogenesis of critical illness myopathy.
The aim of the present study was to evaluate the effect of illness on diaphragm fiber-type expression, local TH metabolism, and mitochondrial activity. To address this question, two in vivo NTIS mouse models were used: a model for lethal sepsis induced by intranasal inoculation of S. pneumoniae, and a model for chronic inflammation induced by a subcutaneous injection of turpentine in the hind limb. In addition, an in vitro model was used for acute inflammation, stimulating C2C12 myotubes with bacterial endotoxin (lipopolysaccharide, LPS) to assess its effect on mitochondrial activity with and without silencing Dio2.
Material and Methods
Animal experiment
Female C57Bl6 mice (Harlan Sprague-Dawley, Horst, The Netherlands) were used at 8–12 weeks of age. The mice were kept in 12-hour light/dark cycles in a temperature-controlled room. Bacterial sepsis was induced by intranasal (i.n.) inoculation of S. pneumoniae serotype 3 (American Type Culture Collection, Manassas, VA). Bacteria were grown in Todd–Hewitt broth (Difco, Detroit, MI) at 37°C, harvested at the mid-logarithmic phase, washed twice in sterile saline, and resuspended in sterile saline at a concentration of 5 × 104 colony forming units (CFU)/50 μL. Lightly anesthetized mice (n = 5–8/group) by inhalation of isoflurane were inoculated i.n. with 50 μL of bacterial solution containing 5 × 104 CFU. Control mice received 50 μL of sterile saline i.n. Local chronic inflammation was induced by subcutaneous (s.c.) injection of 100 μL of steam-distilled turpentine in each hind limb under isoflurane, as described previously (15). Control mice received 100 μL of sterile saline in each hind limb. At time points 24 and 40 hours after inoculation of S. pneumoniae or 48 and 120 hours after turpentine injection, either five to eight or six mice per group, respectively, were anaesthetized with isoflurane and subsequently killed by cervical dislocation. Diaphragm muscle tissue was obtained and immediately stored in liquid nitrogen. The gastrocnemius, a fast-twitch muscle, and the soleus, a slow-twitch muscle, was also obtained from the mice infected with S. pneumoniae and the matched control animals. Both studies were approved by the local animal welfare committee.
Cell culture experiments and transfection
The mouse muscle cell line C2C12 (myoblasts) was cultured in Dulbecco's modified Eagle's medium (DMEM; with 4.5 g/L glucose and L-glutamine, supplemented with 10 IU/mL of penicillin, streptomycin, and fungizone, all from Lonza, Basel, Switzerland) plus 10% fetal bovine serum (Lonza). On day 2, the medium was refreshed with differentiation medium in order to differentiate myoblasts into myotubes (DMEM basic medium plus 2% horse serum; Life Technologies, Carlsbad, CA). After another 24 hours, the medium was replaced by differentiation medium plus 4 μg/mL arabinoside C (proliferation inhibitor; Sigma, St. Louis, MO) during three consecutive days. The myotubes were transfected before analyzing the cellular bio-energetic profile. At day 4 after seeding, Dio2 expression in myotubes was silenced using a siRNA approach using Dio2 specific Stealth™-siRNAs, as described before (16). siRNAs resulted in 80–90% knockdown after 16–48 hours, which was validated in all experiments performed. Scrambled siRNA with matching low GC content was used as control (#12935-300; Life Technologies). Using the HiPerfect kit (Quiagen, Venlo, NL), 5 nM of siRNA were transfected, and 24 hours after transfection, 10 ng/mL of LPS was added to the medium for 18 hours, and the cellular bio-energetic profile was obtained.
For RNA isolation, 1 × 105 C2C12 myoblast/well were seeded in a 0.1% gelatine-coated (Merck, Kenilworth, NJ) six-well plate (NUNC, Rochester, NY). The differentiation into myotubes was performed as described above. Six hours after stimulation with LPS or 10 ng/mL of IL-1β (Sigma), the myotubes were washed twice with cold phosphate-buffered saline, scraped, and centrifuged for 2 min at 13,225 g at 4°C. The pellet was resuspended in RNA lysis buffer for RNA extraction and stored at −80°C until use.
Cellular bio-energetic profile
Mitochondrial function in C2C12 myotubes was assessed using the Seahorse XFe96 (Seahorse Bioscience, North Billerica, MA). A total of 5 × 103 C2C12 myoblasts were seeded in 0.1% gelatine-coated XF96 cell culture microplates (Seahorse Bioscience) in DMEM medium and differentiated into myotubes, as described above. On the day of measurement, the medium was replaced by Seahorse assay medium (DMEM with 25 mM glucose, 1 mM pyruvate, 2 mM glutamine without sodium bicarbonate; Sigma), and the cells were placed in an incubator without CO2 at 37°C for 1 h. The oxygen consumption ratio (OCR) was measured three times at the basal condition, and twice after 1.5 μM oligomycin (OM, ATP synthase inhibitor), another twice after 10 μM carbonyl cyanide-p-trifluoromethoxyphenyl-hydrazon (FCCP, uncoupling agent) injection, and subsequently twice after adding 2.5 μM of antimycin-A (AM) +1.25 μM rotenone (Ro; both inhibitors of the electron transport chain) in each well. These measurements at the XFe96 Extracellular Flux Analyzer (Seahorse Bioscience) consisted of three-minute cycles of mixing and two-minutes cycles of measuring during the basal condition, after the injection of OM, and after AM + Ro injections, or five-minute cycles of mixing and two-minute cycles of measurement after FCCP injection. All measurements were performed in each well of the XF96 cell culture microplates (Seahorse Bioscience) at the same time, and each condition was measured independently in at least 10 replicates per plate. All mitochondrial stress drugs were purchased from Seahorse Bioscience. The OCR/min ratio was corrected by the DNA content of each well using the CyQuant reagent (Life Technologies), according to the manufacturer's protocol. This approach provided the following information regarding mitochondrial function: basal respiration (first three measurements points), ATP production related to mitochondrial phosphorylation capacity (first two measurements after OM), the maximal respiratory capacity (first two measurements after FCCP), and the non-mitochondrial respiration (first two measurements after AM/Ro).
RNA isolation and quantitative polymerase chain reaction
Diaphragm RNA from S. pneumoniae–infected mice and matching controls was isolated using the MagNA Pure LC RNA Isolation Kit III–Tissue (Roche Molecular Biochemicals, Mannheim, Germany), according to manufacturer's protocols. The diaphragm RNA from the turpentine-injected mice and matching controls was isolated using the TRIzol® Reagent (Invitrogen, Carlsbad, CA) and Macherey Nagel kit (Macherey Nagel, Düren, Germany), according to each manufacturer's protocol. RNA yield was measured using a Nanodrop (Nanodrop, Wilmington, DE) in order to perform the cDNA synthesis with equal RNA-input. cDNA synthesis was performed using the First Strand cDNA Synthesis Kit for reverse transcription polymerase chain reaction with oligo d(T) primers (Roche), according to the manufacturer's protocols.
Quantitative polymerase chain reaction (qPCR) was performed using a Lightcycler 480 (Roche) with SensiFAST master mix (Biolane, London, United Kingdom) and 50 ng primer (Biolegio, Nijmegen, The Netherlands). Primer sequences and annealing temperatures are given in Table 1. Primers were typically intron spanning. If this was not the case, genomic DNA contamination was tested using a cDNA synthesis reaction without the addition of reverse transcriptase. The qPCR programs was: denaturation 5 min at 95°C, 45–55 cycles of 5 sec at 95°C, 10 sec annealing temperature, 15 sec at 72°C. Quantification was performed using the LinReg software (17). The mean efficiency was calculated for each assay, and samples that had a deviation of >5% were excluded. Calculated values were normalized by the geometric mean of the reference values (Rplp0, Phactr4, and Hprt1 for diaphragm samples; Hprt1 and Ppib for soleus and gastrocnemius samples; Hprt1 and G6pd2 for C2C12 samples). The best reference genes were chosen using the GeNORM qBase+ software (Biogazelle, Ghent, Belgium).
Statistics
Statistical analyses were carried out using Graph Pad Prism v5 (Prism Software, La Jolla, CA). The normal distribution of the data was evaluated using the Shapiro–Wilk normality test. Differences between groups were analyzed using one way analysis of variance followed by either the Mann–Whitney U-test if not normally distributed or by Student's t-test in normally distributed data. p-Values of <0.05 were considered statistically significant. Results are shown as mean values ± standard error of the mean (SEM) of at least four animals per group, or, in case of the in vitro experiments, as four independent experiments.
Results
IL-6 increases in diaphragm during sepsis
In order to evaluate the diaphragm inflammatory response during illness, Il6 mRNA expression was analyzed, as well as macrophage infiltration by the expression of Emr1 (F4/80), a macrophage marker. Il6 mRNA expression increased in the diaphragm 24 and 40 hours after the induction of bacterial sepsis compared with controls (Fig. 1A; p = 0.02 and p < 0.001). Sepsis also increased Emr1 mRNA expression in the diaphragm, but only after 24 hours (Fig. 1B; p = 0.009). Socs3 (a suppressor of cytokine signaling—SOCS3) mRNA expression increased 40 hours after infection (Fig. 1C; p = 0.04).
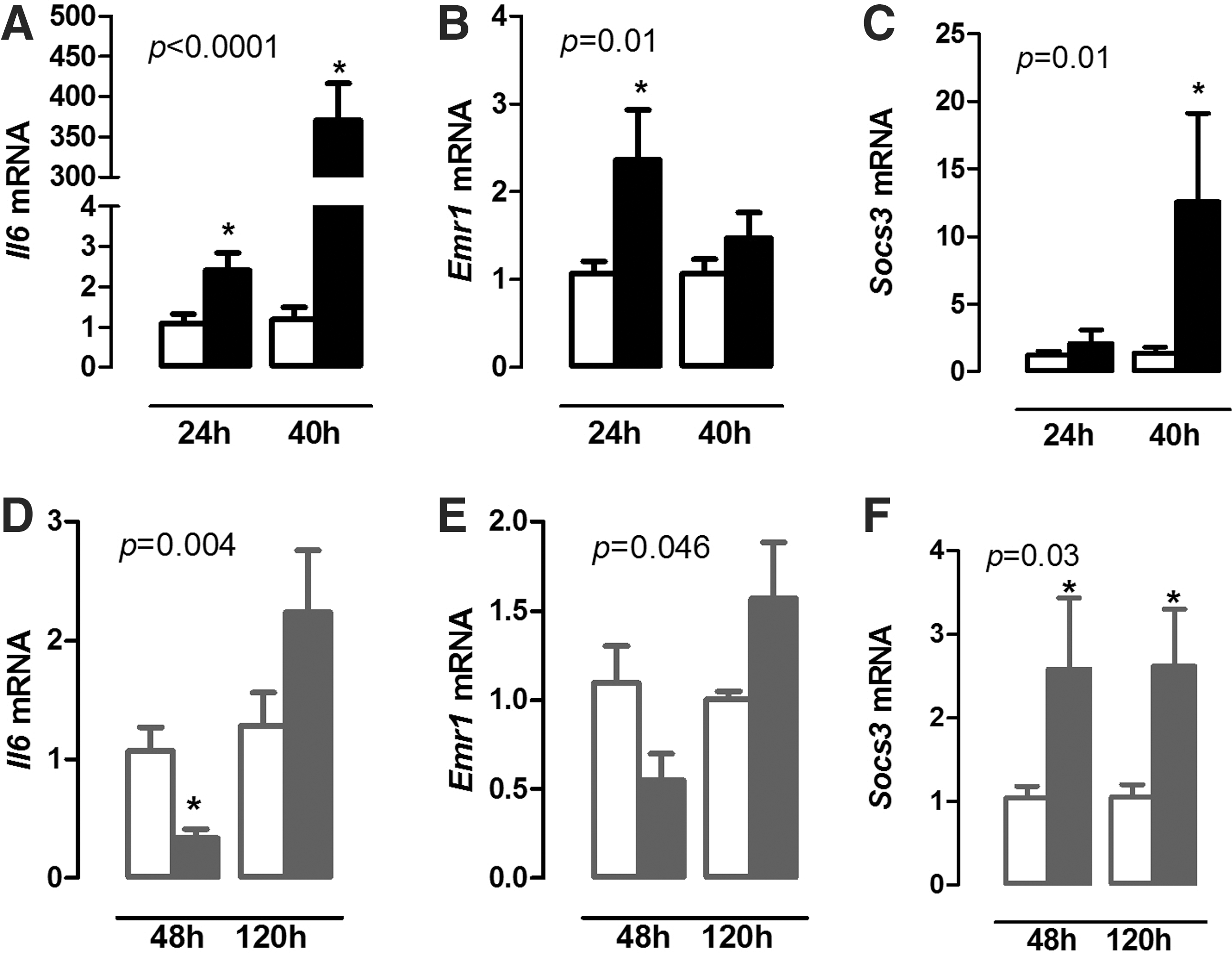
Relative expression of Il6 (
Chronic inflammation resulted in decreased Il6 mRNA in the diaphragm 48 hours after the onset of inflammation, and returned to control levels at 120 hours (Fig. 1D; p = 0.004). No significant difference was observed in the Emr1 expression (Fig. 1E), while Socs3 mRNA expression increased at 48 and 120 hours in the mice with chronic inflammation compared with control mice (Fig. 1F; p = 0.03).
Inflammation modulates diaphragm fiber type
In order to understand the changes in diaphragm physiology during illness better, muscle-fiber composition was studied by evaluating mRNA expression of type 1 fiber markers (slow twitch) Myh7 (MHC I) and Atp2a2 (SERCA2a), and type 2 fiber markers (fast twitch) Myh4 (Mhc IIb), Myh1 (MHC IIx), Myh2 (MHC IIa), and Atp2a1 (SERCA1).
Bacterial sepsis increased Myh7 and Atp2a2 mRNA expression after 24 hours (Fig. 2A and B; p = 0.03 and p = 0.004, respectively), while there was no significant difference at 40 hours. No significant changes were observed in the fiber II markers Myh4, Myh1, Myh2, and Atp2a1 (Fig. 2C–F) mRNA at any time point. As the diaphragm consists of both fast and slow fibers, muscle-fiber composition was analyzed in the gastrocnemius, a fast-twitch muscle, and the soleus, a slow-twitch muscle. Sepsis induced an increase in Myh7 (MHC I) and Atp2a2 (SERCA2a) expression in the gastrocnemius, while only an increase in ATp2a1 (SERCA1) was observed in the soleus (Supplementary Fig. S1; Supplementary Data are available online at
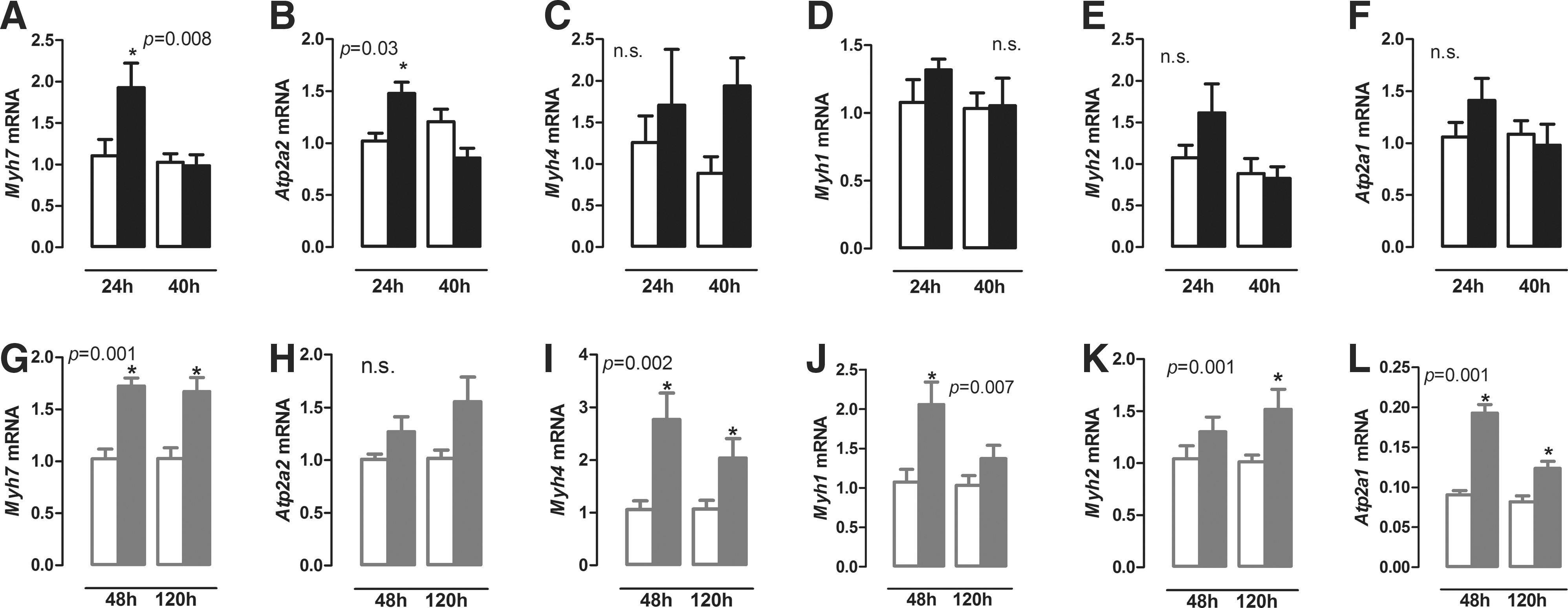
Relative expression of Myh7 (
Chronic inflammation resulted in increased Myh7, Myh4, Myh1, and Atp2a1 mRNA expression in the diaphragm after 48 hours (Fig. 2G, I, J, and L; p = 0.004, p = 0.009, p = 0.002, and p = 0.004, respectively), whereas Atp2a2 mRNA expression did not change (Fig. 2H). Increased expression of Myh7, Myh4, and Atp2a1 mRNA was also observed after 120 hours (Fig. 2G, I, and L; p = 0.004, p = 0.015, and p = 0.009, respectively), while Myh1 mRNA returned to control levels, and Myh2 mRNA expression increased only after 120 hours (Fig. 2J and K; p = 0.009 and p = 0.015, respectively). In summary, bacterial sepsis and chronic inflammation induce marked and differential changes in mRNA profiles defining distinct fiber types.
TH metabolism in the diaphragm is differentially modulated during illness
Since TH are important determinants of muscle fiber type, Dio2, Dio3, Thra1, and Thrb1 mRNA expression was investigated in order to establish whether changes in fiber-type profile are associated with putative changes in T3 availability.
Dio3 mRNA expression increased in the diaphragm 24 hours after the induction of sepsis (Fig. 3A; p = 0.04). This change was transient, as Dio3 mRNA restored to control levels 40 hours after infection (Fig. 3A). Dio2 mRNA expression decreased 40 hours after infection compared with controls (Fig. 3B; p = 0.005).
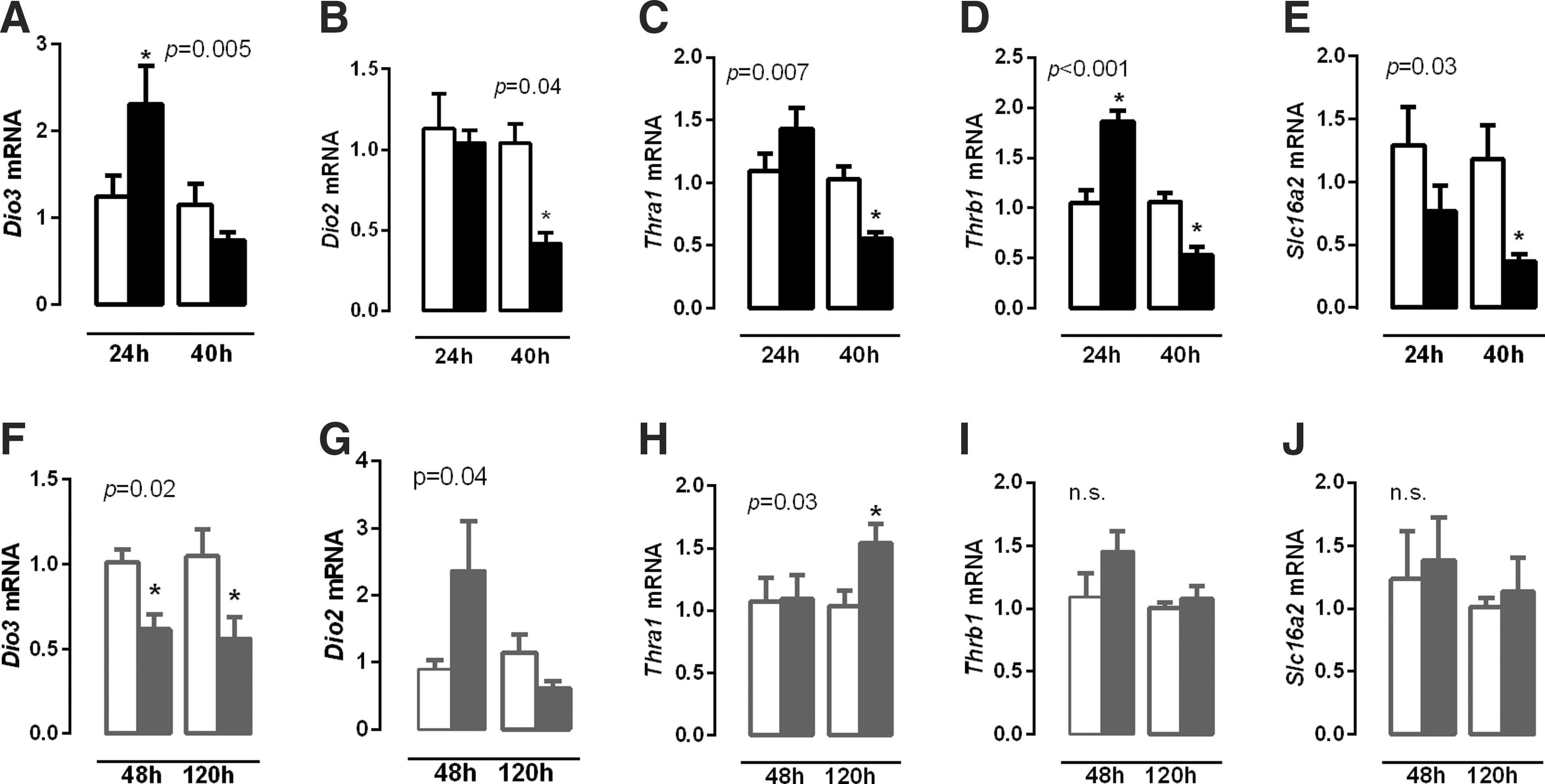
Relative expression of Dio3 (
Thra1 mRNA expression did not differ within 24 hours during sepsis, but Thrb1 mRNA expression increased compared with controls (Fig. 3C and D; p = 0.006). After 40 hours of bacterial sepsis, Thra1 and Thrb1 mRNA expression both decreased (Fig. 3C and D; p = 0.007 and p = 0.002, respectively) compared with control mice. Slc16a2 (Mct8) mRNA expression decreased in the diaphragm 40 hours after the induction of sepsis (Fig. 3E; p = 0.005).
Chronic inflammation appeared to affect TH metabolism in the diaphragm differentially; Dio3 mRNA expression decreased 48 and 120 hours after turpentine injection (Fig. 3F; p = 0.02 and p = 0.04, respectively), while Dio2 mRNA expression tended to increase after 48 hours compared with controls (Fig. 3G; p = 0.08). Thra1 mRNA increased at 120 hours (Fig. 3H; p = 0.03), and no significant changes were observed in Thrb1 or Slc16a2 mRNA expression (Fig. 3I and J).
In order to determine alterations in T3 responsiveness during illness, the mRNA expression of two positively T3 regulated genes were investigated: troponin (Tnni2), a motor protein expressed preferentially in type II fibers, and myogenin (Myog), a myogenic transcription factor.
Septic mice did not show significant differences in Tnni2 and Myog mRNA expression after 24 hours, but both Tnni2 and Myog expression decreased after 40 hours, suggesting decreased TH availability (Fig. 4B and C; p = 0.04 and p = 0.004, respectively). During chronic inflammation, Tnni2 tended to increase 48 hours after turpentine injection (Fig. 4D and E; p = 0.065) while Myog mRNA expression increased twofold in the diaphragm 48 hours after turpentine administration, but returned to control levels again at 120 hours (Fig. 4F; p = 0.04). In summary, sepsis resulted in a setting consistent with decreased TH signaling with a reversed pattern in chronic inflammation.
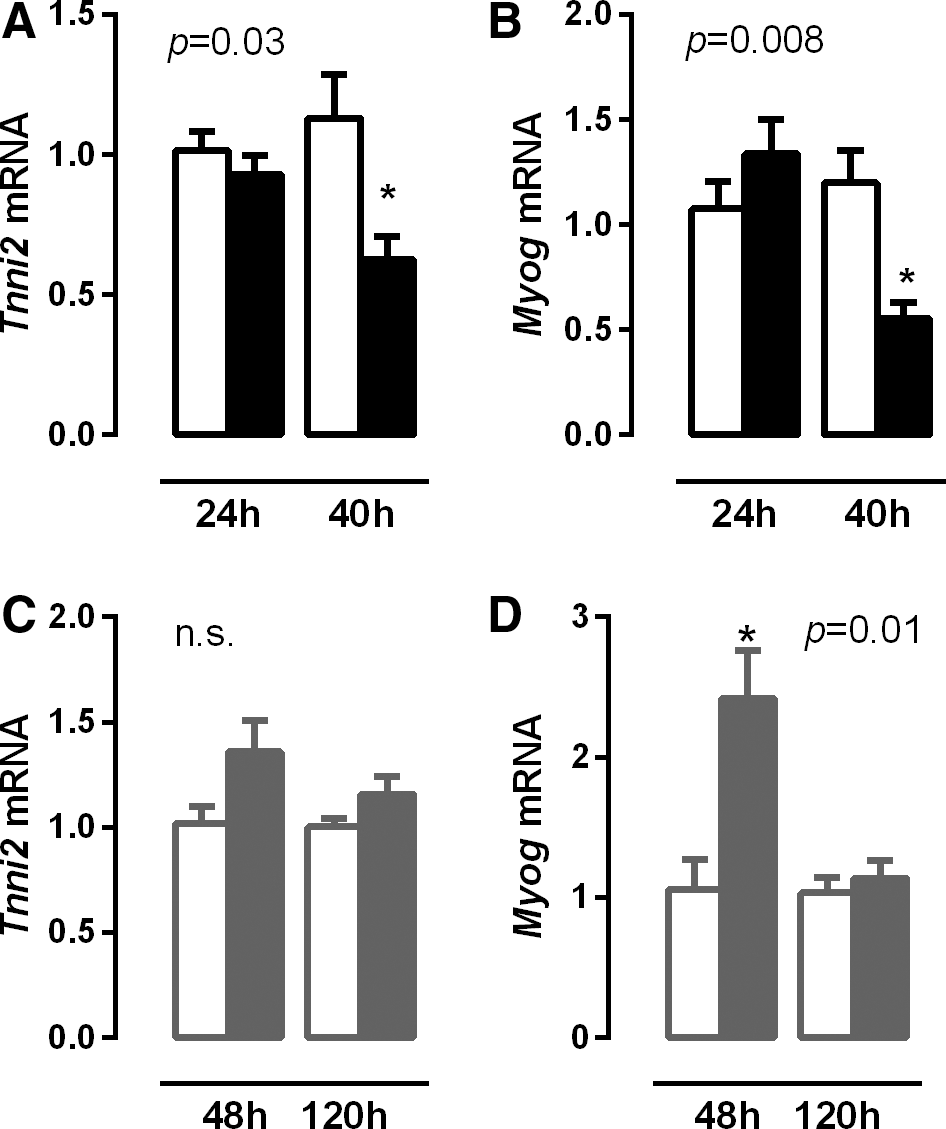
Relative expression of Tnni2 (
Influence of the inflammatory status on mitochondrial metabolism
Intracellular T3 controls cellular metabolism via proteins involved in mitochondrial function. Ppargc1a (Pgc1α), Ucp3, and Slc2a4 (GLUT4) are positively regulated by T3. Since were observed changes in TH metabolism in the diaphragm of infected mice, mRNA expression of Ppargc1a, a main transcriptional coactivator in the control of mitochondrial biogenesis, was investigated. In addition, Ucp3, associated with fatty acid oxidation in muscle, as well as Slc2a4, which codes for the main glucose transporter in skeletal muscle (GLUT4), was assessed in the diaphragm of mice with lethal sepsis and chronic inflammation.
Bacterial sepsis significantly increased Ucp3 mRNA expression after 40 hours (Fig. 5A; p < 0.05). By contrast, Ppargc1a (PGC1α) and Slc2a4 (GLUT4) mRNA expression decreased at the same time point (Fig. 5B and C; p = 0.0008 and p = 0.02, respectively), suggesting a decrease in mitochondrial efficiency and glucose uptake during the later stages of sepsis.

Relative expression of Ucp3 (
During chronic inflammation, Ucp3 mRNA expression increased after 120 hours compared with controls (Fig. 5D; p = 0.015). Ppargc1a mRNA expression did not change (Fig. 5E), while Slc2a4 mRNA expression increased in the diaphragm 120 hours after turpentine administration (Fig. 5F; p = 0.03).
In order to study the role of inflammation-induced alterations in TH metabolism on muscle mitochondrial function in more detail, C2C12 myotubes were stimulated with LPS. LPS stimulation induced an inflammatory response, reflected by increased Il6 mRNA expression (Fig. 6A; p = 0.002) and reduced Dio2 mRNA expression (Fig. 6B; p = 0.0007). IL-1β stimulation induced the same profile in C2C12 myotubes as LPS treatment (Supplementary Fig. S2). No significant differences in ATP-linked oxygen consumption (which correlates with the mitochondrial phosphorylating capacity) or in the maximal oxygen consumption at complex IV (which correlates with the maximal mitochondrial respiratory capacity) were observed (data not shown). However, LPS stimulation decreased basal respiration and non-mitochondrial respiration (Fig. 6C and D; p = 0.002 and p = 0.008, respectively).
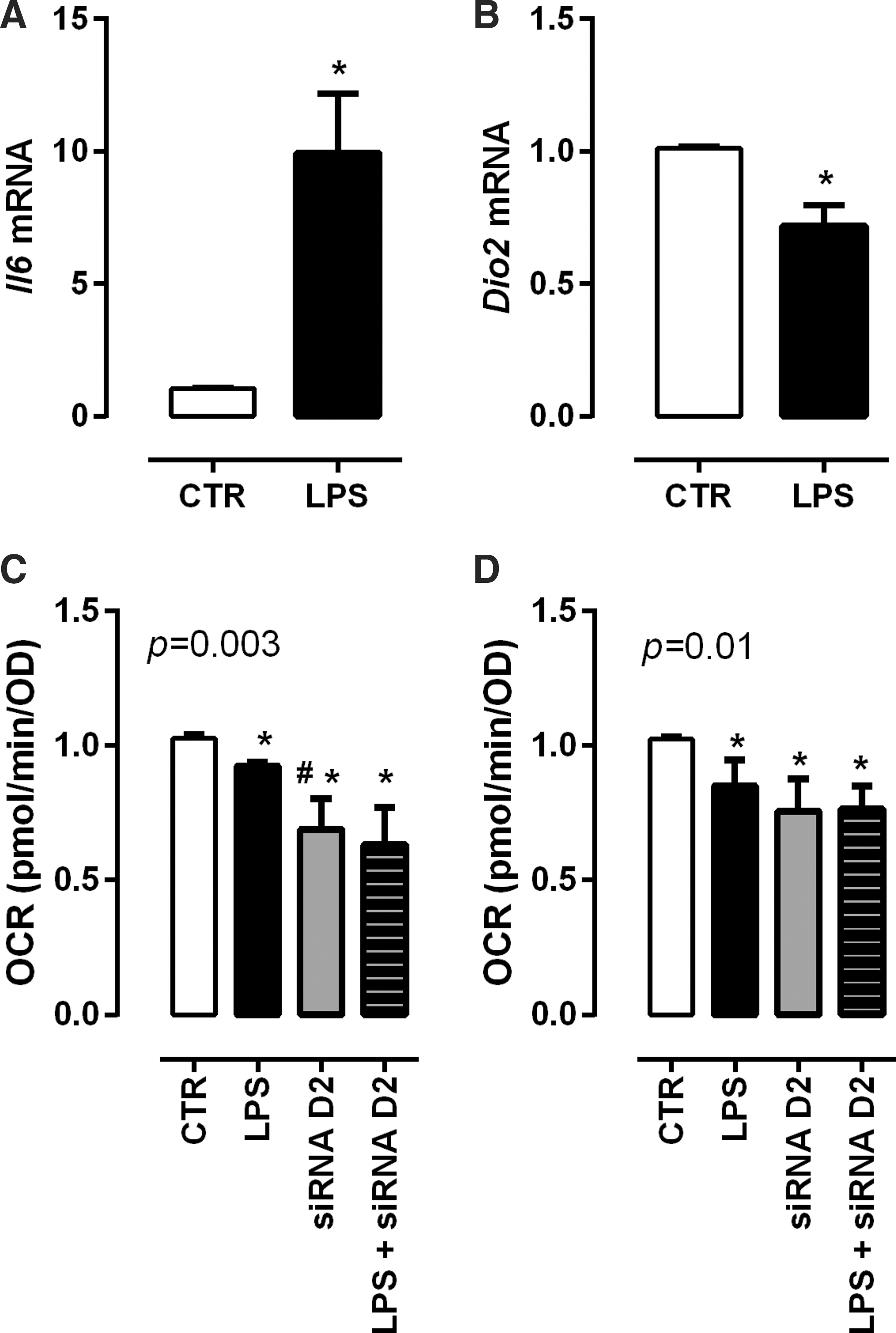
Relative expression of Il6 (
In order to link the LPS-induced effects to TH metabolism, Dio2 was inhibited using a siRNA approach. Dio2 knockdown reduced basal respiration and non-mitochondrial respiration of the myotubes independently of LPS (Fig. 6C and D; p = 0.008). Dio2 knockdown in C2C12 myotubes decreased basal oxygen consumption by 31%, which is substantially more than the 10% decrease observed after LPS stimulation (Fig. 6C; p = 0.02). This is in agreement with the observed Dio2 decrease. Of interest, the siRNA knockdown resulted in a Dio2 knockdown of 80%, while LPS administration reduced Dio2 by 30% expression (Supplementary Fig. S3).
Discussion
The present study reports the effects of illness on TH metabolism and muscle fiber type profile in the diaphragm using two well-defined NTIS models: bacterial sepsis and chronic inflammation (3). It was found that illness induced significant changes in the diaphragm with regard to mRNA expression of genes coding for muscle fiber types, local TH metabolism, and mitochondrial function. In vitro experiments established a role of D2 in cellular respiration. In addition, the joint alterations in TH metabolism, established by determination of Dio2, Dio3, Thra1, Thrb1, and Slc16a2 (MCT8) expression, were dependent of illness type and resulted in changes in T3 responsiveness. It is tempting to speculate that the decrease in mitochondrial function is associated with low intracellular T3 concentrations as reflected by the decreased expression of T3 responsive genes.
mRNA expression of a variety of muscle fiber type markers in the diaphragm was evaluated during illness. Bacterial sepsis might increase type I fibers shortly after infection, since an increase in the expression of Myh7 (MHC I) and Atp2a2 (SERCA2a) mRNA was observed, both of which are type I fiber markers. However, no significant difference was observed at a later stage. S. pneumoniae–induced sepsis is lethal within 48–72 hours, and the half-life of a muscle myosin fiber is between 10 and 18 days (18,19). Therefore, a prolonged illness model might be necessary to study long-term effects on fiber type. Nevertheless, the increase of Atp2a2 (SERCA2a) expression could stimulate contraction of the diaphragm during the onset of sepsis due to intensification of the relaxation rate of the type I fibers. Zolfaghari et al. demonstrated a decrease in isometric force and maximal power generated in the diaphragm of 24-hour septic animals (12), which correlates with an increase in fiber type I. LPS stimulation of C2C12 cells, a mouse muscle cell line, reduced Myh7 (MHC I) expression, which is associated with increased NFκB signaling (20). Additionally, TNFα, a pro-inflammatory cytokine, reduces Atp2a2 (SERCA2a) mRNA expression in smooth muscle cells (21). Thus, the increase of Myh7 and Atp2a2 in the diaphragm during bacterial sepsis is unlikely to be a direct result of inflammatory mediators on the muscle fibers.
Sepsis seems to modulate tissue T3 availability and responsiveness (4). This study observed an early increase in Dio3 expression and decreased Dio2, Trha1, Trhb1, and Slc16a2 (MCT8) expression in the diaphragm of septic mice later on, suggesting decreased TH responsiveness. To confirm the hypothetical low intracellular T3 concentrations, Tnni2 and Myog expression, both positively regulated genes, were evaluated in the diaphragm. Indeed, both Tnni2 and Myog expression decreased, suggesting decreased TH responsiveness. This could lead to a shift of fiber type IIa, the oxidative and slowest type II fiber (fast twitch), into type I (slow twitch), thereby increasing Myh7 and Atp2a2 mRNA expression. To explore the hypothesis that sepsis induces a slower type profile by increase fiber type I–related gene expression, the gastrocnemius, a fast-twitch muscle, and the soleus, a slow-twitch muscle, were analyzed in septic mice. In support of the hypothesis, an increase in Myh7 and Atp2a2 expression in the gastrocnemius was observed, while no changes in these genes were observed in the soleus.
During chronic inflammation, an increase was observed in both Myh7 and Myh4 (MHC IIb) expression, the slowest and fastest myosin types, respectively, and in Atp2a1, a marker of fiber type II, at all time points studied. An increase in Myh1 (MHC IIx) was also observed at 48 hours and in Myh2 (MHC IIa) at 120 hours, both expressed in type II fibers. The increase in Myh4 and Atp2a1 could be due to a rise in intracellular T3, since these genes are positively regulated by T3. However, the increase in TH responsiveness in the diaphragm should also lead to a decrease in Myh7, a gene that is negatively regulated by T3, which was not the case. Nevertheless, these data suggest that chronic inflammation resulted predominantly in increased expression of genes expressed in fast-twitch fibers of the diaphragm. Muscle fiber types are controlled by two major stimuli: the rate of the action potential in the motor neuronal plate, and intracellular T3 signaling (14). T3 exerts its action by increasing Myh2, Myh1, and Myh4 (MHC II a, x, and b, respectively) expression and decreasing Myh7, thereby switching to the fast fiber twitch profile. T3 also increases Atp2a2a and Atp2a1 expression, which increase relaxation rate and energy demand (14). Consequently, increased TH availability might explain the changes in fiber type genes in the diaphragm. A decrease in Dio3 expression was also observed, while Dio2 mRNA tended to increase, favoring the formation of T3 in the diaphragm, during chronic inflammation (Fig. 3). Although Tnni2 expression did not change, Myog expression increased, suggesting increased T3 responsiveness.
The observed alterations in TH metabolism (decreased Dio2 and increased Dio3) during sepsis might be a direct effect of proinflammatory cytokines, as stimulation of C2C12 myotubes induced a similar profile, while chronic inflammation, characterized by an absent local IL-6 response, resulted in decreased Dio3 expression. In both in vivo inflammatory models, increased Socs3 mRNA expression was observed, which indicates active inflammatory signaling in the diaphragm. The differential IL-6 response, however, suggests that the inflammatory response differs in both models. This might explain the observed differences in TH metabolism and thereby muscle physiology during sepsis and chronic inflammation.
Muscle dysfunction is associated with mitochondrial failure (22). Increased Ucp3 associated with decreased Slc2a4 (GLUT4) and Ppargc1a (PGC1α) mRNA expression, as observed in this study during sepsis, suggests reduced mitochondrial biogenesis and energy uptake by the diaphragm cells. Additionally, infection decreases fatty acid oxidation in the diaphragm (23), thereby reducing the availability of fuel. The diaphragm consists of a mix of fast- and slow-twitch fibers (24), and the increase in slow-twitch fibers and the reduction in the fatty acid oxidation in the diaphragm during sepsis could contribute to muscle failure.
It has been shown that the mitochondrial membrane potential is decreased in diaphragm from septic animals (12), which could be associated with increased Ucp3 expression (25). Indeed, increased Ucp3 expression was observed during chronic inflammation and sepsis, while a differential response of TH metabolism was observed in both models. This could be explained by the fact that the sepsis-induced increase in skeletal Ucp3 mRNA is associated with an increase in glucocorticoid signaling (26). Thus, based on the changes in Ucp3, Ppargc1a, and Slc2a4 expression, it is currently only possible to speculate about the changes in the mitochondrial function and the role of TH.
An in vitro model was used to study muscular mitochondrial function. To this end, C2C12 myotubes were stimulated with LPS. Recently, Frisard et al. demonstrated a decrease in maximal respiratory function in C2C12 myotubes after acute expose to low doses of LPS (27). Decreased basal oxygen consumption in C2C12 myotubes after LPS treatment, which is associated with a decrease in Dio2 expression in these cells, was also observed in this study. To study a causal role of D2, Dio2 was silenced using a siRNA approach. The knockdown of Dio2 mimicked the effect of LPS, reducing basal and non-mitochondrial oxygen consumption, suggesting a role for T3 in mitochondrial function. However, a recent study by Werneck-de-Castro et al. (28) showed that D2 knockdown in myocytes does not affect T3 responsive gene expression in muscle greatly, casting doubt about the potential importance of myocyte-specific D2 expression. Alternatively, D2 expression in adipose tissue or macrophages, also present in skeletal muscle, could contribute to muscle TH metabolism. The macrophage marker Emr1 in the diaphragm was studied during illness, and indeed, Emr1 is expressed in the diaphragm.
The present study demonstrates that illness differentially modulates TH metabolism in the diaphragm of mice in association with changes in the fiber-type profile. Sepsis induced an acute increase in the expression of genes reflecting diaphragm fiber type I while reducing mitochondrial function. These effects were accompanied by alterations in TH metabolism indicative of decreased TH responsiveness (Fig. 7). Mitochondrial function of C2C12 myotubes was reduced after LPS stimulation in association with reduced Dio2 expression. Dio2 silencing in C2C12 cells recapitulated reduced mitochondrial function, indicating that part of the changes in mitochondrial function might be due to reduced T3 availability. The use of cell-specific D2 knockout models will be necessary to establish a causal role for TH in illness-induced alterations in diaphragm muscle physiology.
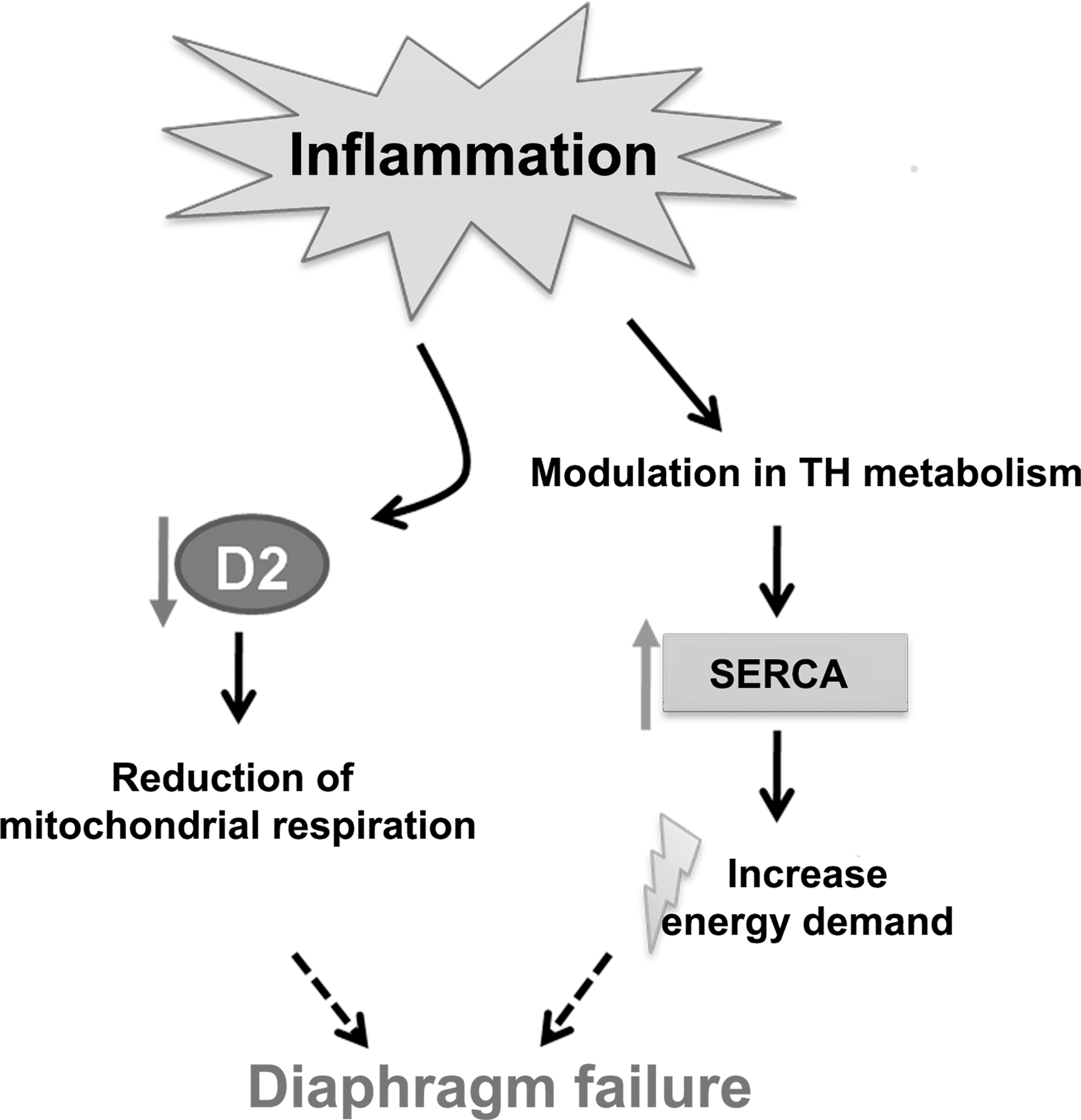
Proposed model of the effects of inflammation on diaphragm function via thyroid hormone (TH) action; inflammation influences alterations in TH metabolism in the diaphragm that leads to increase in SERCA expression, which could increase the energy demand, while the observed D2 decrease leads to reduced cellular respiration.
Footnotes
Acknowledgments
The authors would like to thank Dr. R.H.L. Houtkooper (Laboratory of Genetic Metabolic Diseases, Academic Medical Center) for the technical support. These studies were supported by Science Without Borders—CAPES Foundation, Ministry of Education of Brazil, Brasilia, Brazil.
Author Disclosure Statement
The authors do not have conflicting interests.