
Abstract
Background:
Several functional single-nucleotide polymorphisms (SNPs) at the FOXE1 locus on chromosome 9q22.33 have been associated with the risk for papillary thyroid carcinoma (PTC). This study set out to elucidate whether their effects are independent, using genotyping results in populations of Asian and European descent.
Methods:
SNPs rs965513 and rs1867277 and a polymorphic region determining the length of the FOXE1 polyalanine (poly-Ala) tract were genotyped in 501 patients with PTC and 748 healthy individuals from Japan, and in 660 patients and 820 population controls from Belarus. Functional analysis of transactivation activities of FOXE1 isoforms with varying number of alanine repeats was performed by a Dual-Luciferase® Assay.
Results:
All three polymorphisms were significantly associated with PTC in both populations on univariate analysis. However, conditional analysis revealed independent effects of rs965513 and rs1867277 SNPs but not of the FOXE1 poly-Ala polymorphism. The independent effect of the lead rs965513 SNP was observed in both populations, while that of rs1867277 was only identified in the Japanese population, in which linkage disequilibrium between the three polymorphisms is markedly weaker. Despite the strong decrease in transcriptional activity with increasing FOXE1 poly-Ala tract length, no difference in transactivation potential of the FOXE1 poly-Ala isoforms could be seen after adjustment for the minimal promoter activity in the reporter vectors. Plasmids encoding FOXE1 isoforms of increasing poly-Ala tract length were also found to produce less FOXE1 protein after cell transfection.
Conclusions:
The functional variants rs965513 and rs1867277 independently contribute to genetic predisposition to PTC, while a contributing role of the FOXE1 poly-Ala polymorphism could not be confirmed.
Introduction
T
Localized about 60 kb upstream and centromeric to FOXE1, rs966513 was the first SNP reported as a genetic determinant of susceptibility to thyroid cancer in a genome-wide association study (1), but its functional relevance was only established recently (23). The lead rs966513, as well as several other SNPs on 9q22.33 that are in linkage disequilibrium (LD) with rs966513, were shown to modify the activities of long-range enhancers involved in the transcriptional regulation of FOXE1 and PTCSC2 (PTC susceptibility candidate 2, gene ID: 101928337), a newly discovered thyroid-specific long intergenic noncoding RNA gene whose chromosomal position partly overlaps with that of the FOXE1 promoter (24).
Another functional variant, rs1867277, located in the FOXE1 5′-UTR (c.-283) has been found to confer risk to differentiated thyroid cancer in the large-scale target gene association study in individuals from Spain and Italy (2). The finding was reproduced by other groups in different populations (5,6,9,13,14,25,26) and confirmed by meta-analyses (15,16). This variant is also involved in the regulation of FOXE1 expression through differential recruitment of USF1/USF2 transcription factors.
In the coding region, FOXE1 possesses a multinucleotide polymorphism, which consists of a variable number of trinucleotides (most commonly GCC, less frequently GCT or GCA, all encoding alanine) ranging from 11 to 22 repeats, hereby referred to as the FOXE1 poly-Ala polymorphism. The most common alleles encode 14 and 16 alanine residues. Polyalanine tracts are a frequent feature of conserved transcription factors, and have been implicated in a number of congenital malformation syndromes (27,28). Variation in the FOXE1 polyalanine tract length has been associated with susceptibility to thyroid dysgenesis (29 –31) and, more recently, with thyroid cancer. The poly-Ala14 has been shown to be protective, and poly-Ala16 a risk-conferring allele (5,12,14,25,26). These observations were confirmed by meta-analysis (16). The transcriptional activity of FOXE1 poly-Ala16 was found to be diminished compared with that of poly-Ala14 (26), although the difference in transactivation potential was not observed between poly-Ala14 and shorter isoforms in an earlier study (29).
To the best of the authors' knowledge, studies on FOXE1 poly-Ala polymorphism in thyroid cancer have not been performed in individuals of Asian origin. The only information on this genetic variant in corresponding populations is from a study of 46 Japanese patients with thyroid dysgenesis (29), and 110 cases of idiopatic premature ovarian failure and 110 controls from China (32). This work is therefore the first to characterize the FOXE1 poly-Ala in a large Japanese cohort.
The objective of this study was to determine whether the three FOXE1 polymorphisms with functional roles may have independent effects on risk for thyroid cancer. For this purpose, genotypes of patients with PTC and population controls of diverse ethnic backgrounds from Japan and Belarus were analyzed, and functional analysis was performed of transactivation activities of five isoforms of FOXE1 with different lengths of polyalanine tract in a normal human thyroid cell line and thyroid cancer cell lines.
Methods
Study populations
A total of 501 patients aged 13–87 years operated for PTC at Kuma Hospital (Kobe, Japan) were enrolled (4). As population controls, 748 Japanese individuals aged 20–76 years at sampling were recruited from Nagasaki University (Nagasaki, Japan). Participants from Belarus included 660 patients with PTC aged 2–22 years at diagnosis and 820 population controls aged 16–49 years at sampling (3). None of the Japanese individuals had a history of radiation exposure. All PTC patients from Belarus and 620 (75.6%) control individuals were exposed to radiation as a result of the Chernobyl accident. Informed consent was obtained from all individual participants included in the study. The protocol of the study was approved by the ethics committees of all participating institutions.
DNA extraction
In the Japanese cohort, DNA was extracted from formalin-fixed paraffin-embedded tissues of PTC patients (4), and from the peripheral blood of control individuals using a QIAamp DNA mini kit (Qiagen, Tokyo, Japan). In the Belarusian cohort, DNA was extracted from peripheral blood mononuclear cells of all participants with a Puregene kit (Qiagen, Germantown, MD) (3). DNA concentration was measured with a Nanodrop 1000 spectrophotometer (Thermo Scientific, Waltham, MA), and samples were stored at −80°C until use.
SNP genotyping and FOXE1 poly-Ala tract length measurement
In both Japanese and Belarusian cohorts, rs965513 and rs1867277 genotyping was performed with predesigned custom ABI TaqMan SNP assays (C_1593670_20 and C_11736668_10, respectively), as described before (13).
FOXE1 poly-Ala tract length was measured by resolving polymerase chain reaction (PCR) products obtained by amplification with a 5′FAM-labeled forward primer and an unlabeled reverse primer flanking the region encoding the FOXE1 poly-Ala polymorphism in an ABI Prism 3130xl genetic analyzer (Applied Biosystems, Foster City, CA) in GeneScan mode. Data were processed using GeneMapper v3.7 software. Direct sequencing of PCR products obtained from 47 randomly chosen samples was used for fine adjustment of GeneScan data. An example of a chromatogram and the detailed protocol are presented in Supplementary Figure S1 (Supplementary Data are available online at
Cell cultures
The immortalized normal human thyroid cell line Nthy-ori 3-1 and the PTC cell line KTC-1 were grown in RPMI-1640 medium supplemented with 5% fetal bovine serum (FBS) and 1% penicillin–streptomycin. The human PTC cells line TPC1 and the follicular thyroid cancer WRO cell line were maintained in Dulbecco's modified Eagles medium supplemented with 10% FBS, 1% nonessential amino acids, 1 mM of sodium pyruvate, and 1% penicillin–streptomycin. All cell lines were cultured in monolayers at 37°C in a humidified 5% CO2 environment.
Expression vectors and reporter plasmids
PCR products of FOXE1 with 12, 14, 15, 16, or 19 alanine repeats variants were obtained using corresponding genomic DNA as a template with the forward 5′-
Reporter plasmids were based on the pGL4.23[luc2/minP] luciferase vector (Promega, Madison, WI), which originally contains a 32 bp minimal promoter that regulates the expression of the firefly luc2 gene (
To prepare the thyroperoxidase (TPO) promoter-driven reporter plasmid, a fragment of the TPO promoter was PCR-amplified, as described before (33), with the following primers: forward 5′-act g
To prepare the FOXE1 response element (FRE)-driven reporter plasmid, 1 μg of each sense 5′-phospho-tcg aTA CTT AAA CAA ACA GAA-3′ and antisense 5′-phospho-tcg aTT CTG TTT GTT TAA GTA-3′ oligonucleotides were annealed and catenated with T4 DNA ligase (NEB) at 16°C. The sequence of the putative FOXE1 response element (capital characters) was derived from previous work (34); overhangs (tcga) corresponding to XhoI sites and allowing subsequent ligation are shown in lower case. Catenated products were resolved in 1% TAE-agarose, fragments between 200 and 300 bp were excised from the gel, purified using a FastGene Gel/PCR Extraction kit (Nippon Genetics, Kawaguchi City, Japan), and ligated with XhoI-digested and shrimp alkaline phosphatase-treated (Takara Bio Inc., Otsu, Shiga, Japan) pGL4.23[luc2/minP]. Plasmids obtained from individual colonies were screened by PCR and subsequent sequencing in order to identify a clone containing the 10xFRE insert in the sense orientation upstream of the minimal promoter; this plasmid was further propagated, sequenced, and used in downstream experiments.
Transfection and Dual-Luciferase® Assay
Assays were performed with the Dual-Luciferase Reporter Assay System (Promega) according to the manufacturer's protocols. Cells were co-transfected by electroporation using a Neon Transfection System (Invitrogen) with 0.6 μg of FOXE1 expression plasmid, 0.6 μg of firefly, and 6 ng renilla luciferase reporter vectors, maintained in 24-well plates and assayed for luciferase activity after 48 h.
The transactivation of TPO or FRE promoters by different FOXE1 poly-Ala variants was determined as the ratio between firefly and renilla luciferase signals, relative to the ratio obtained in the cells co-transfected with the corresponding FOXE1 expression plasmids and a non-modified pCMV6-AC-IRES-GFP as control. All experiments were performed in quadruplicates and were reproduced several times.
Western blotting and quantitative real-time PCR
One day before transfection, 6 × 105 Nthy-ori 3-1 cells were plated in a 10 cm dish in medium without antibiotics. The following day, cells were co-transfected with 4 μg of pCMV6-AC-IRES-GFP-FOXE1 expression plasmids or empty pCMV6-AC-IRES-GFP vector, 4 μg of pGL4.23-10xFRE, and 80 ng of pGl4.74 luciferase reporter vectors using 20 μL of Lipofectamine 2000 (Invitrogen) according to the manufacturer's protocol. After 3 h of incubation with DNA-Lipofectamine complexes, the medium was replaced with fresh medium without antibiotics. After 48 h, cells were scraped in ice-cold phosphate-buffered saline for subsequent protein extraction for the Dual-Luciferase Assay, Western blotting, and DNA isolation.
For the Dual-Luciferase Assay, approximately one-third of cells were treated with 1 × Passive Lysis Buffer (Promega), and the assay was performed as described above.
For Western blotting, approximately one-third of cells were lysed in a buffer containing 20 mM of Tris-HCl, 150 mM of NaCl, 1 mM of EDTA, 0.5% Triton X-100, 5% glycerol, 2 mM of PMSF, 50 mM of NaF, 10 mM of sodium pyrophosphate, 1 mM of sodium orthovanadate, and 1 × Complete Protease Inhibitor Cocktail (Roche Diagnostics K.K., Tokyo, Japan). After measuring protein concentration with a Pierce BCA Protein Assay Kit (Thermo Scientific, Rockford, IL), 40 μg of lysates were resolved in 4–15% gradient SDS-polyacrylamide gel (Mini-Protean TGX; Bio-Rad, Hercules, CA), and blotted onto a PVDF membrane (Trans-Blot Turbo Transfer Pack mini; Bio-Rad). After blocking with 10% skim milk in TBST for 1 h, the membrane was incubated overnight at 4°C with anti-TTF2 rabbit polyclonal antibodies (PA0200; Biopat, Perillo Sant'Angelo a Cupolo, Italy) diluted 1:1000 in 5% skim milk in TBST. After 2 × washing for 5 min with TBST, secondary HRP-conjugated anti-rabbit antibodies (sc7074; Cell Signaling Technology, Tokyo, Japan) diluted 1:1000 in 5% skim milk in TBST were applied for 1 h at room temperature. After three wash steps for 5 min in TBST, the membrane was incubated in Pierce Western Blotting Substrate (Thermo Scientific, Rockford, IL) for 1 min at ambient temperature. Luminescence detection was performed in a LAS-4000 mini imaging system (Fujifilm, Tokyo, Japan). Next, after incubation with Western Blot Stripping Buffer (Thermo Scientific, Rockford, IL) for 45 min at 37°C, the membrane was reprobed for 1 h at room temperature with a primary anti-β-actin mouse monoclonal antibody (sc-827; Santa Cruz Biotechnology, Santa Cruz, CA) diluted 1:1000 in 5% skim milk in TBST and secondary HRP-conjugated anti-mouse antibody (sc7076; Cell Signaling Technology) diluted 1:1000 in 5% skim milk in TBST. β-Actin signal was detected as described above. After densitometry, FOXE1 levels were normalized to the corresponding β-actin levels.
DNA was extracted from the remaining cells using a QIAamp DNA Mini Kit (Qiagen, Tokyo, Japan). Quantitative real-time PCR was performed with the following primers: TG forward, GTG AGG GCA CAC ATG CTT CAT and TG reverse, CGG AGC TTT GCT TCC TCA CA (amplifying a 113 bp fragment of the human thyroglobulin [TG] gene, gene ID: 7038), and FOXE1 forward, CGC CAT GCT GCC GCT TAT and FOXE1 reverse, CTT ATC GTC GTC ATC CTT GTA ATC CAG (amplifying a 126 bp region of plasmid-encoded FOXE1), and SYBR Premix Ex Taq II reagent (Takara Bio, Inc.). All reactions were performed in a Thermal Cycler Dice Real Time System II (Takara Bio, Inc.) under the same conditions: 95°C for 30 sec, then 40 cycles of 95°C for 5 sec and 60°C for 30 sec followed by dissociation curve analysis to ensure the signal from target amplicon. Plasmid DNA quantity was normalized for nuclear DNA.
Statistical analysis
Differences between case and control groups for each FOXE1 polymorphism were examined using logistic regression analysis in the multiplicative model of inheritance. Association, LD, and haplotype analyses were performed using “gap” and “haplo.stats” R packages. Analysis of transactivation effects of different FOXE1 isoforms and correlation analysis was performed using SPSS Statistics for Windows v21 (IBM Corp., Armonk, NY) and GraphPad InStat v3.10 (GraphPad Software, San Diego, CA). p-Values were two-sided and considered significant if <0.05.
Results
Genotyping
All individuals from Japan (501 patients with PTC and 748 population controls) were successfully genotyped for rs965513, rs1867277, and the FOXE1 poly-Ala polymorphism. In the Belarusian series (660 patients with PTC and 820 population controls), rs965513 genotypes were obtained for all participants, rs1867277 genotypes were determined in 624 PTCs and 760 controls (0.945 and 0.927 call rates, respectively), and the FOXE1 poly-Ala polymorphism was characterized in 635 PTCs and 777 controls (0.962 and 0.948 call rates, respectively). Genotyping results are shown in Table 1.
Presented p-values are not adjusted for multiple testing; all p-values, except that for the poly-Ala other/16 analysis in the Japanese series, remained statistically significant after Bonferroni correction.
Risk allele.
PTC, papillary thyroid carcinoma; HWE, Hardy–Weinberg equilibrium; MAF, minor allele frequency; OR, odds ratio; CI, confidence interval.
Information on FOXE1 polyalanine tract length is extremely limited for Asian populations. This study therefore presents the prevalence and distribution of the corresponding alleles and genotypes in the present series in some detail. In the Japanese cohort, the length of the poly-Ala tract varied from 14 to 17 repeats (except for 13 and 15) in healthy individuals, and from 11 to 16 (except for 13 and 15) in PTC patients. In the Belarusian cohort, the range was from 12 to 19 repeats (except for 15 and 18) in healthy individuals, and from 12 to 19 (except for 13, 15, and 18) in PTC patients. The FOXE1 Ala14 allele was the most prevalent in both cohorts (Fig. 1), accounting for 99.0% in healthy Japanese individuals and 97.5% in Japanese PTC patients, and for 59.7% and 51.5%, respectively, in the Belarusian cohort. The second prevalent allele was Ala16, which was observed with the frequencies of 0.9–1.2% in the Japanese series and 34.6–41.7% in the Belarusian cohort.
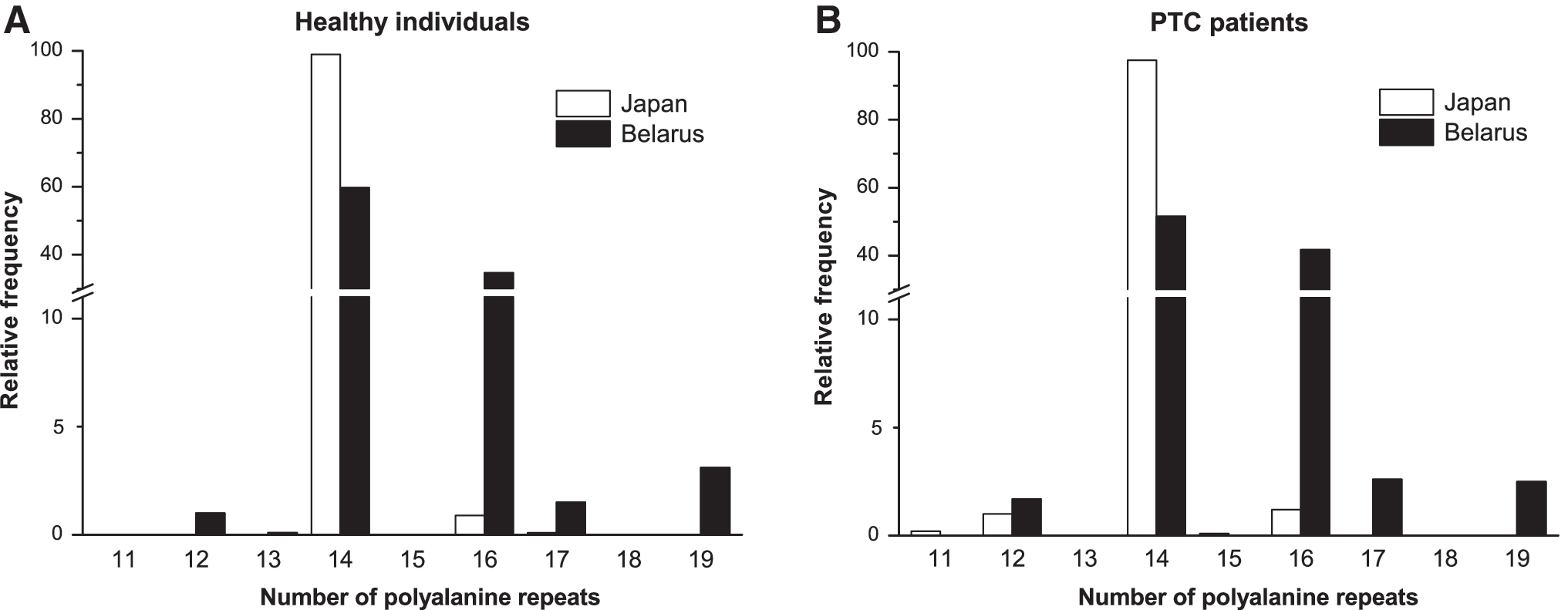
Relative frequency of FOXE1 poly-Ala alleles in the Japanese and Belarusian cohorts. Frequencies are shown for the groups of (
Homozygosity for the Ala14/14 genotype was the most frequent and found in 98.0% in healthy Japanese individuals and 95.4% in Japanese PTC patients (Supplementary Table S1). The second prevalent genotype in the Japanese cohort presented by the two most frequent alleles, heterozygosity for Ala14/16 was observed only in 1.9% of healthy individuals and 2.2% patients; no homozygous Ala16/16 genotype was found. In the Belarusian cohort, corresponding genotypes accounted for 36.0%, 40.9%, and 12.1% in healthy individuals, and for 27.1%, 42.0%, and 18.0% in PTC patients.
Association analysis of FOXE1 polymorphisms with PTC
All three genetic variants displayed statistically significant individual association signals, which remained significant after Bonferroni correction for multiple testing in both Japanese and Belarusian cohorts (Table 1). The differences in effect sizes between the two ethnic groups were not statistically significant (p > 0.08–0.90). There was, however, a noticeable difference in the minor allele frequencies of all studied polymorphisms ranging from 0.01 to 0.15 in the Japanese cohort compared with 0.35–0.48 in the Belarusian cohort. The frequency of FOXE1 poly-Ala variants other than 14 repeats was particularly low in the Japanese individuals; this was the most likely reason for insufficient power to detect a significant association of the FOXE1 poly-Ala16 allele with PTC (p > 0.5) clearly seen in the Belarusian cohort (p = 0.0001219).
To determine whether the effects of the tree polymorphisms are independent, a conditional analysis was performed (Table 2). In both ethnic groups, rs965513 remained significant under any condition (i.e., after it was conditioned on rs1867277, poly-Ala, or both polymorphisms), indicating that this is the lead SNP with an independent signal (lowest p = 0.0001967).
The FOXE1 poly-Ala14/other model.
A variance inflation factor ≥5 was observed in the model, suggestive of considerable collinearity between predictors.
NA, not applicable.
In the Japanese cohort, rs1867277 remained significant under all conditions, although with smaller effect size and weaker significance (lowest p = 0.001386) than for rs965513, strongly suggestive of its independent effect. In contrast, the FOXE1 poly-Ala was weakly significant after conditioning on the distal rs965513 (p = 0.039), but lost the association signal after conditioning on the proximal rs1867277 or on both rs965513 and rs1867277 (p = 0.070 and 0.253, respectively). Although because of low frequency of variants other than poly-Ala14 this result needs to be interpreted with some caution, it suggests that the FOXE1 poly-Ala may not have an independent effect. In the Belarusian cohort, rs1867277 lost its significance after conditioning on rs965513 or on rs965513 and poly-Ala combined (p = 0.475 and 0.338, respectively) but remained weakly significant after conditioning on the latter (p = 0.040). Thus, the effect of rs1867277 could not be detected upon comparison with that of rs965513, but could be distinguished from the poly-Ala in this ethnic group. An independent role of FOXE1 poly-Ala was not observed after conditioning on either rs965513 or on both rs965513 and rs1867277 (lowest p = 0.589), corroborating these findings in the Japanese cohort.
LD analysis and FOXE1 haplotype association with PTC
The above-mentioned two observations—the drastic differences in allelic frequencies of genetic variants between the Japanese and Belarusian cohorts, and a high collinearity between rs1867277 and FOXE1 poly-Ala in the Belarusian cohort in the conditional regression models (Table 2 footnote)—prompted an evaluation of LD between the three polymorphisms and an examination of haplotype associations with PTC in the two ethnic groups.
LD in the Japanese cohort was substantially weaker compared with the Belarusian series (Fig. 2). In the Japanese cohort, only four haplotypes were identified, likely due to the low frequency of polymorphisms under analysis. All these haplotypes were associated with PTC (Table 3). The most prevalent haplotype was observed with about 80% prevalence, included all protective alleles (i.e., rs965513[G], rs1867277[G], and poly-Ala14), and negatively associated with PTC risk (odds ratio [OR] = 0.547, p = 0.000000006601). Of note, two consequent haplotypes associating with an increased risk for PTC (OR = 1.548, p = 0.0007432; and OR = 1.968, p = 0.0001088, respectively) harbored either one rs1867277[A] or one rs965513[A] allele, but both contained the protective poly-Ala14. This observation again supports the limited contribution of the FOXE1 poly-Ala to the risk of PTC compared with those of rs965513 and rs1867277.
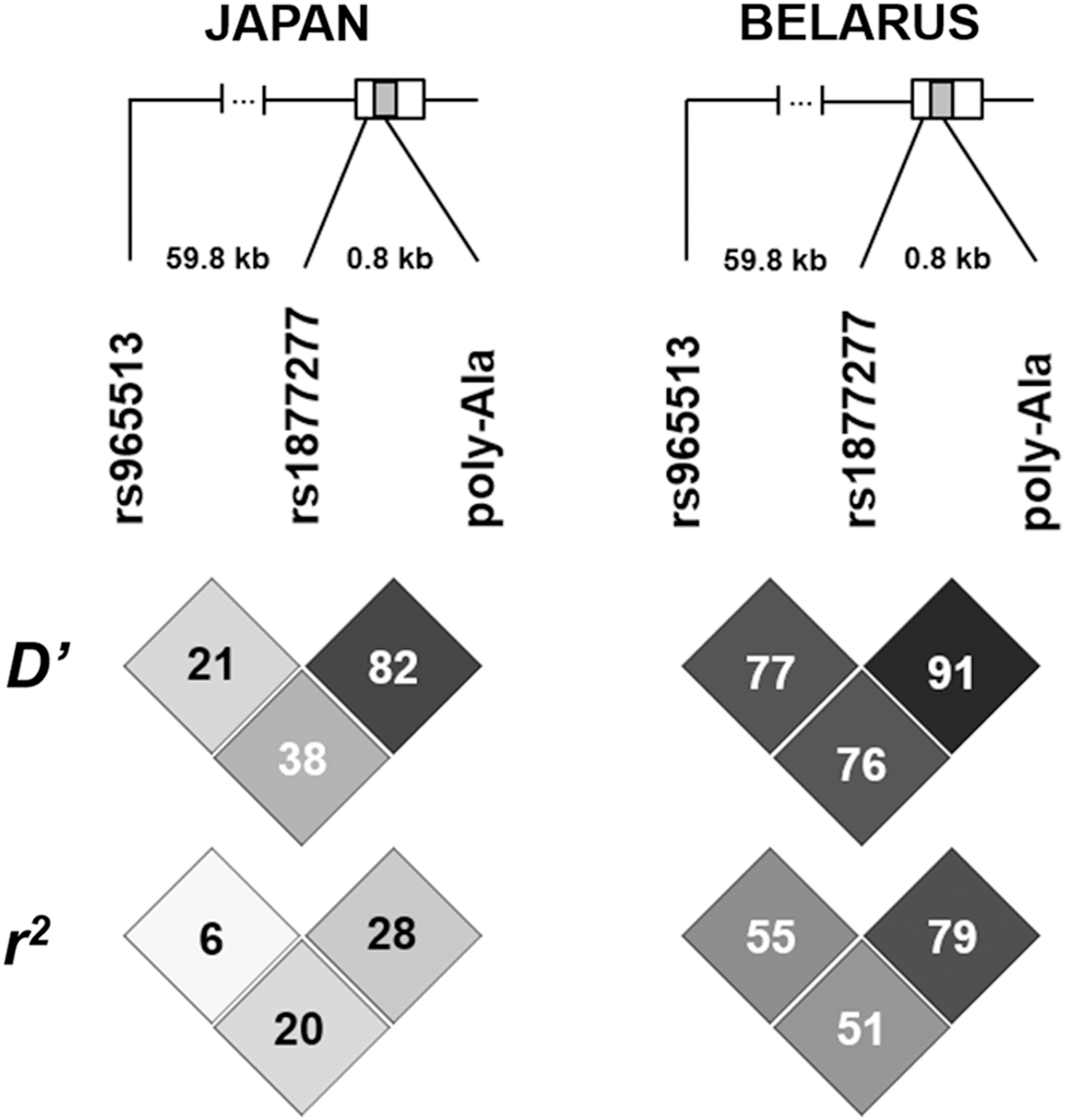
Schematic representation of three polymorphisms in the FOXE1 locus at chromosome 9q22.33 and corresponding linkage disequilibrium (LD) measures. The FOXE1 gene is shown as a rectangle with coding region of a single exon shaded; unshaded regions represent the 5′- and 3′-UTR. Linear distances between polymorphic sites are indicated (59.8 kb and 0.8 kb). For D′ and r 2, the intensity of box shading is proportional to the corresponding measures (black and white colors represent the strong or weak LD, respectively).
Indicated OR is for comparison of a given haplotype versus all other.
Any nucleotide.
In the Belarusian cohort, among seven haplotypes, only two were significantly associated with PTC (Table 3). Similar to the results in the Japanese cohort, the most prevalent haplotype, accounting for about 50%, included all protective alleles and was negatively associated with PTC (OR = 0.688, p = 0.0000004810). The only haplotype conferring elevated risk for PTC contained risk alleles of all there polymorphisms (i.e., rs965513[A], rs1867277[A], and poly-Ala_non-14). Findings in the Belarusian series again do not enable distinguishing independent contributions of polymorphisms to the risk for developing PTC, most likely due to strong LD in this ethnic group.
In view of differences in allelic frequencies of the three polymorphisms under investigation, this study additionally assessed whether other genetic variants in the 100 kb region of chromosome 9q22.33 encompassing rs965513 and the FOXE1 gene may have similar frequencies in populations of Asian and European ancestry. In Asian populations (JPT and HCB), minor allele frequencies of the common SNPs varied from 0.003 to 0.453 (M = 0.108, median 0.091; Supplementary Table S2). In European individuals, the range was from 0 to 0.498 (M = 0.278, median 0.337). The difference between allelic frequencies in the two ethnic groups was statistically significant (p = 0.00000006705), demonstrating that polymorphic variants in this chromosomal region are less frequent in Asian than in European populations.
Transactivation effects of FOXE1 isoforms
Five different FOXE1 expression constructs (encoding 12, 14, 15, 16, and 19 poly-Ala repeats) were functionally examined in dual luciferase assays for their ability to activate the reporter expression driven by the minimal 32 bp promoter, or TPO or 10xFRE promoters in an immortalized normal human thyroid cell line (Nthy-ori 3-1) and three differentiated human thyroid cancer cell lines (TPC1, KTC-1, and WRO), which have relatively low level of endogenous FOXE1 protein (Supplementary Fig. S2).
A strong decrease in transactivation potential of FOXE1 isoforms with increasing poly-Ala tract length was observed for all cell lines and promoters (Supplementary Fig. S3 and Supplementary Table S3). The negative correlation was also seen for data aggregated by cell or promoter type (Fig. 3 and Table 4 univariate analysis). Of note, the FOXE1 poly-Ala tract length–dependent activation of the minimal promoter was also significant (Fig. 3 and Table 4 univariate analysis) and deserves special attention, since the minimal promoter is a constituent part of TPO- and 10xFRE-driven reporter vectors. When these activities were controlled for minimal promoter activation, the effect of the FOXE1 poly-Ala tract length was no longer observed in regression models (Table 4 multivariate analysis).
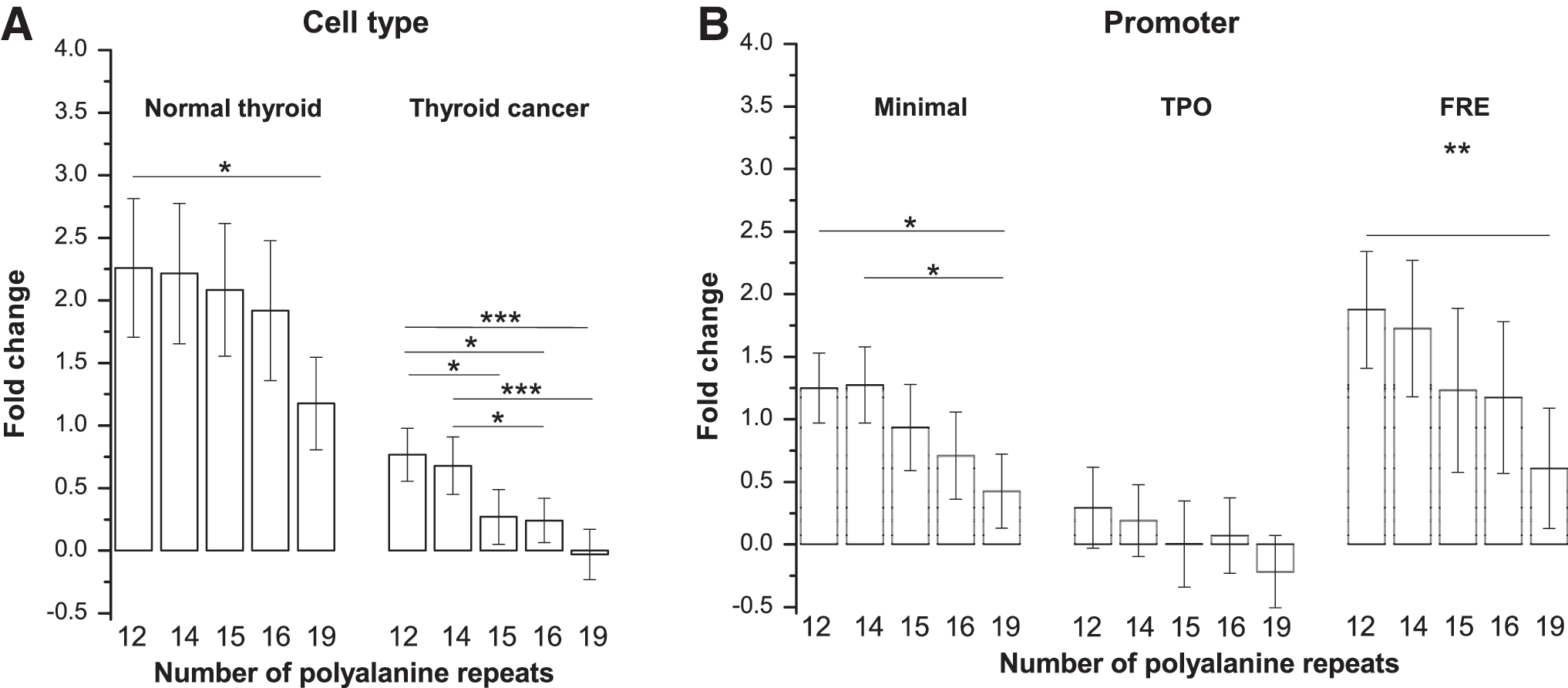
Effect of different FOXE1 isoforms on activation of the reporter expression. (
Transcriptional activity of FOXE1 isoforms with 12, 14, 15, 16, or 19 alanine repeats were tested. Mean fold change in luciferase activity compared to an empty pCMV6-AC-IRES-GFP vector served as an outcome variable. All variables were ln-transformed and standardized by promoter type.
Statistical significance of the regression model for each type of cells.
Regression coefficient of the factor.
Standard error of the regression coefficient.
Standardized regression coefficient.
Statistical significance of the regression coefficient.
Effects of FOXE1 poly-Ala isoforms and of minimal promoter tested independently.
Effects of FOXE1 poly-Ala isoforms and of minimal promoter tested simultaneously.
ANOVA, analysis of variance.
To address the reason for the decline in reporter signal with increasing FOXE1 poly-Ala tract length, Western blotting, Dual-Luciferase Assay, and real-time PCR analysis of corresponding materials from the same transfection experiments were performed in Nthy-ori 3-1 cells. The declining pattern of reporter signal was reproduced as in previous functional assays (Fig. 4). Surprisingly, it was found that despite equal amounts of different FOXE1 poly-Ala isoform-encoding plasmids being used for transfections, FOXE1 protein levels and the levels of cell-associated FOXE1 expression vectors also changed as a function of the poly-Ala tract length. Strong positive correlations between the three endpoints of these assays were confirmed statistically. A plausible explanation could be that plasmids encoding FOXE1 isoforms of increasing length may display declining transfection efficiencies, decreased translation with increasing trinucleotide length, or a declining stability inside cells after transfection, eventually affecting FOXE1 protein levels (but this is not due to vector degradation before transfection; Supplementary Fig. S4). Further experiments would be necessary to distinguish between these scenarios and shed light on the underlying mechanism.
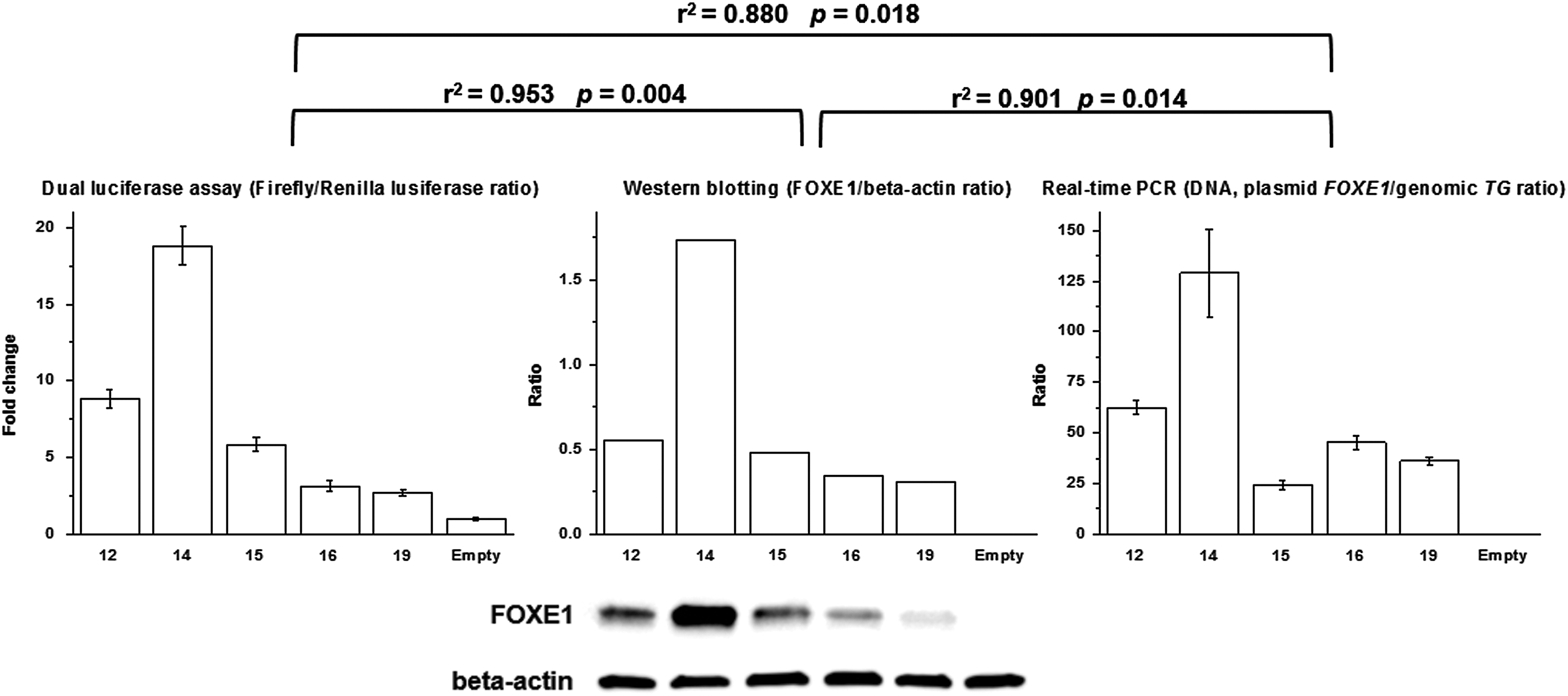
Transactivation activity of different FOXE1 isoforms (on the 10xFRE promoter), FOXE1 protein levels, and real-time quantification of cell-associated FOXE1 isoform expression vector levels in Nthy-ori 3-1 cells. Protein extracts for dual luciferase assays and Western blotting, and DNA for real-time polymerase chain reaction assays were obtained from the portions of cells collected from the same dishes 48 hours after transfection. The coefficient of determination (r 2) and statistical significance of Pearson correlation coefficient are indicated for each pair of endpoints. Shown are the results of a representative experiment. All experiments were reproduced three times with a similar result.
Discussion
This study aimed to answer whether three functional polymorphisms on chromosome 9q22.33, which have been reported in association with thyroid cancer, may play independent roles. It is noteworthy that the associations of genetic variants in the FOXE1 locus have been initially reported in genome-wide studies for both adult sporadic and radiation-related PTC in young patients with similar effect sizes in the cohorts of Caucasian origin (1,3). The associations were later replicated in Japanese and Chinese studies of adult sporadic thyroid cancer, also identifying similar effect sizes (4,7). Thus, there is no evidence for age-dependent, etiological, or ethnical correlations for inherited genetic variants at this locus that would cause potential bias once association analyses are performed separately within the ethnic groups.
Several converging lines of evidence indicated that two SNPs, rs965513 and rs1867277, located 60 kb upstream or immediately in the FOXE1 gene, respectively, are likely to have independent signals. In contrast, an independent role for FOXE1 poly-Ala could not be demonstrated.
First, the association signals of rs965513 and rs1867277 were replicated in the Japanese and Belarusian populations with effect sizes very similar to those reported before, that is with an OR of 1.6–1.9 for rs965513 and of 1.5–2.0 for rs1867277 (1 –14,25,26). An association of the FOXE1 poly-Ala tract with PTC was also confirmed in both populations with an allelic OR comparable to previously published values of 1.3–2.5 in different ethnicities (5,14,26). In the Belarusian population, the association signal of the FOXE1 poly-Ala tract was seen in the poly-Ala14/other and the poly-Ala other/16 models, confirming the protective effect of the poly-Ala14 allele and the risk-conferring role of poly-Ala16, which is in line with earlier reports. In the Japanese cohort, a significant association signal was revealed only in the poly-Ala14/other model. The reason is that the FOXE1 poly-Ala variant has a very low degree of variability in the Japanese population, with a minor allele frequency of about 1–2%. It, however, did not hamper statistical analysis, demonstrating a significant association with risk for PTC for non-poly-Ala14 alleles. In contrast, the association could not be demonstrated for the poly-Ala16 allele in the Japanese cohort due to its low frequency. A statistical power estimate of case-control sample size indicates the study should have enrolled about 38,000 participants to detect an effect of this allele at OR = 1.3.
The frequency of the FOXE1 poly-Ala14 homozygotes among healthy Japanese individuals was 98.0%, in good agreement with 96.4% found in the Chinese population (32). Importantly, among Japanese patients with PTC, homozygous carriers of the protective FOXE1 poly-Ala14 allele accounted for 95.4% (allelic frequency 97.5%). Given the apparent rarity of the risk-associated non-poly-Ala14 alleles in this group, it would be difficult to assign them a causative role in conferring predisposition to thyroid cancer at the population level, although rare variants may well be used for the identification of disease-associated genes or chromosomal regions (35).
Second, despite several studies having simultaneously genotyped more than one polymorphism in the FOXE1 locus with or without the poly-Ala (5,6,12,14,26), only one examined their independent associations with thyroid cancer. The work by Jones et al. reported that rs965513 and rs1867277 are independent risk alleles based on the analysis of a large series of patients of Caucasian origin and controls from the United Kingdom (6). A conditional analysis was performed, which unambiguously demonstrated that rs965513 is a lead SNP with an independent association signal in both Japanese and Belarusian ethnic groups (Table 2). With regard to rs1867277 in the FOXE1 5′-UTR, the results were different between the Japanese and Belarusian cohorts. While in the Japanese series rs1867277 remained significant under all conditions, indicative of its independent signal, the association was lost in the Belarusian group after conditioning on rs965513, or on rs965513 and poly-Ala together, likely due to a strong LD between rs1867277 and rs965513. This finding in the Belarusian group is at variance with the report by Jones et al., and may stem from the difference in sample size or different LD relationships in the populations enrolled in the two works. However, the analysis in the Japanese population is in line with the report by Jones et al., and supports the independent effect of rs1867277.
In the Belarusian cohort, rs1867277 remained weakly significant after conditioning on FOXE1 poly-Ala, supportive of the independent role of the former. In contrast, the FOXE1 poly-Ala signal lost significance after conditioning on either the proximal rs1867277 in the 5′-UTR of the gene, or on the combined distal rs965513 and rs1867277 in either population. This finding strongly suggests that the FOXE1 poly-Ala is unlikely to have an independent effect, especially from rs1867277.
Third, LD relationships corresponded well with the results of conditional analysis. In the Japanese cohort, weak LD between the three polymorphisms allowed the intragenic rs1867277 and poly-Ala to remain significant after their conditioning on distal rs965513. However, the somewhat stronger LD between rs1867277 and the FOXE1 poly-Ala results in a non-significant effect of FOXE1 poly-Ala after conditioning on rs1867277, or on rs965513 and rs1867277 combined. In the Belarusian cohort, LD between the three polymorphisms was rather strong, in line with findings in the Portuguese population (5). It is probably for this reason that both rs1867277 and the FOXE1 poly-Ala repeat lost significance after conditioning on the lead rs965513 SNP, making their independent associations with PTC undetectable in this ethnic group. Also, the stronger association signal of rs1867277 and strong LD with FOXE1 poly-Ala rendered the latter non-significant.
Assessment of haplotype associations with PTC in the two populations was in good agreement with the results of the conditional regression analysis. Data from the Japanese group demonstrated a limited, if any, contribution of non-poly-Ala14 alleles to the risk haplotype(s), while the demonstration of independent roles of intragenic FOXE1 polymorphisms in the Belarusian series has proved difficult, likely because of strong LD. Note that with regard to FOXE1 poly-Ala tract, the results of haplotype analysis pertain to its length only, which was the focus of the study. Since a detailed sequence analysis of poly-Ala tract was not performed, it is difficult, for example, to determine whether poly-Ala14 alleles abundant in the Japanese population are identical-by-decent. The low frequency of other common genetic variants in the FOXE1 locus in this ethnic group, however, suggests that the haplotype analysis is rather adequate, although not free of some potential bias.
Finally, although FOXE1 polyalanine tract length-dependent transactivation of the reporter expression was observed in the functional analyses, that is, a decrease in transactivation with increasing FOXE1 polyalanine tract length, as previously reported for the poly-Ala14 and poly-Ala16 isoforms (26), a similar effect on the reporter vectors regulated by the minimal promoter only was also noticed. The reason for the FOXE1-dependent activation of the minimal promoter remains unclear, and it is not known whether FOXE1 interacts directly with these sequences. Nevertheless, transactivation of the minimal promoter by FOXE1 should not be dismissed to avoid technical misinterpretation of the results of functional studies, which indicate that different transactivation capacities of FOXE1 isoforms with different poly-Ala tract length could not be accurately demonstrated in conventional reporter assays employing the vectors containing a particular minimal promoter. In the supportive experiments, it was also observed that the descending pattern of promoter activation with increasing FOXE1 polyalanine tract length could likely be attributed to the decline in transfection efficacies or a declining stability of corresponding FOXE1 expression vectors inside the cells after transfection seen as poly-Ala tract length–related changes in the vector DNA levels. These changes would be expected to affect corresponding FOXE1 protein levels, which were confirmed, and which may be the reason for the difference in the reporter signal intensities, thus masking the potential difference in transactivation activity of different FOXE1 isoforms, if they exist. Additional experiments, using redesigned promoter-driven reporter vectors and controlling for intracellular transgenic FOXE1 levels would be necessary to demonstrate functional differences of the FOXE1 isoforms.
Taken together, the present findings show that rs965513 and rs1867277 SNPs independently associate with risk for thyroid cancer while the multinucleotide FOXE1 poly-Ala polymorphism does not. It should be emphasized that on single-track association analysis, FOXE1 poly-Ala was nominally associated with PTC in this study, in both populations, in full agreement with previous reports (5,12,14,26). However, conditional analysis demonstrated the loss of association when the effects of other SNPs were taken into consideration. The LD analysis showed that the FOXE1 poly-Ala was in strong relationship with rs965513 and rs1867277 in the Belarusian cohort, particularly with the latter. Since rs1867277 is significantly associated with the risk for thyroid cancer, significant FOXE1 poly-Ala association signal could be expected too. The likeliest reason for this, however, is the strong LD of FOXE1 poly-Ala with bona fide risk variant(s) rather than own effect.
It is worth noting that both rs965513 (23,24) and rs1867277 (2) are functionally involved in the transcriptional regulation of FOXE1 and/or PTCSC2. The risk allele of rs965513 was associated with decreased expression of FOXE1, unspliced PTCSC2 and TSHR (thyroid stimulating hormone receptor, gene ID: 7253) in normal thyroid tissue (24). Interestingly, a recent study demonstrated that overexpression of FOXE1 in the thyroids of transgenic mice restrained the proliferation of follicular cells (36), in support of the functional effect of rs965513. In an earlier study, a correlation was also observed between immunohistochemical expression of FOXE1 in PTC tissue and the rs1867277 genotype (37). Ectopic expression of PTCSC2 in a papillary carcinoma cell line resulted in altered expression of a subset of genes implicated in cell cycle and cancer (24). Although whether rs1867277 may regulate PTCSC2 and the precise roles of FOXE1 and/or PTCSC2 in thyroid cancer remain to be established in detail, the growing body of evidence implicates the FOXE1 and PTCSC2 expression levels, which are at least in part regulated by the functional SNPs, in predisposition to PTC.
While it seems reasonable to hypothesize that the poly-Ala polymorphism in the coding region of FOXE1 may also contribute to inherited risk for thyroid cancer, the results of this study favor the notion that the associations with PTC of functional SNPs rs965513 and rs1867277 but not of FOXE1 poly-Ala polymorphism are independent. These findings provide a better understanding of the role of these genetic factors in predisposition to thyroid cancer.
Footnotes
Acknowledgments
We thank Ms. C. Hayashida (Nagasaki University) for technical assistance. This work was supported in part by KAKENHI Grant Number 16H02774, 15K09438 and 26293142 from the Japan Society for the Promotion of Science (JSPS).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.