
Abstract
Background:
Graves' disease (GD) is a common organ-specific autoimmune disease characterized by hyperthyroidism that has significant sex differences in prevalence and clinical expressions. Abnormal cytokine production and T cell activation may result in various manifestations of GD. Studies have shown that androgen treatment can provide protection against autoimmune diseases, but the effects of androgen treatment on GD are still unknown. Therefore, this study investigated whether a potent bioactive androgen, 5α-dihydrotestosterone (DHT), could be of benefit in a BALB/c mouse model of GD. The aims of this study were to investigate (i) whether DHT pretreatment inhibits autoimmune responses, and (ii) the mechanism of immune protection of DHT in GD.
Methods:
Female BALB/c mice were immunized three times with an adenovirus expressing the human thyrotropin receptor (TSHR) A-subunit (Ad-TSHR289). Three doses (1.5, 5, and 15 mg) of DHT or a matching placebo were implanted a week before the first immunization. Four weeks after the third immunization, mice were sacrificed, and blood, the spleen, and the thyroid were removed for further analysis.
Results:
After DHT treatment, thyroid hormones were dramatically reduced compared with placebo. In addition, a remarkable reduction in interferon-γ and interleukin-2 production was observed in DHT-pretreated mice.
Conclusions:
DHT can alleviate the severity of GD by downregulating pro-autoimmune T helper 1 cells in female BALB/c mice. The protective influence was more noticeable with 5 mg and 15 mg doses of DHT.
Introduction
G
Cytokines are potent mediators of communication that influence immune responses, differentiation and activation of T cells, and thus play a key role in the development of autoimmune thyroid diseases (3). A number of cytokines may be associated with the pathogenesis of GD, including interferon-γ (IFN-γ), interleukin-2 (IL-2), interleukin-22 (IL-22), interleukin-17 (IL-17), interleukin-12 (IL-12), interleukin-1β, and tumor necrosis factor-α (TNF-α) (4 –7). Previous studies have shown that an abnormal shifting of the balance between T helper 1 (Th1) and T helper 2 (Th2) cells may lead to the development of GD (3,4).
The incidence of GD is estimated to be approximately 5/10,000 individuals per year (3), and it occurs much more frequently in women than it does in men, with a female-to-male ratio of 7:1 (3). Basic immune responses may be different between men and women, with women producing a more vigorous immune response and elevated antibody production (8). In addition, sex hormones, such as estrogen, testosterone (T), and progesterone, are believed to participate in the immune response and be associated with sex differences in the prevalence of autoimmune diseases (9). Several studies have shown that androgens offer a protective effect in humans and animal models of autoimmune diseases (10,11). Because testosterone can be converted into both other androgens and estrogen, studying the effects of androgens on immunity is difficult. It was hypothesized that a potent bioactive androgen, 5α-dihydrotestosterone (DHT), may protect a BALB/c mouse model from GD. To date, experimental autoimmune GD (EAGD) via immunization with TSHR peptides is the most common mouse model of GD. In this study, the effects of DHT on EAGD were determined, and the mechanism of benefits was investigated.
Materials and Methods
Mice
Female BALB/c mice, aged six to eight weeks, were randomly divided into eight groups: (i) normal mice injected with a null adenovirus (Con); (ii) mice immunized with Ad-TSHR289 (EAGD); (iii) EAGD mice implanted with 1.5 mg of placebo (placebo 1.5 mg); (iv) EAGD mice implanted with 1.5 mg of DHT (DHT 1.5 mg); (v) EAGD mice implanted with 5 mg of placebo (placebo 5 mg); (vi) EAGD mice implanted with 5 mg of DHT (DHT 5 mg); (vii) EAGD mice implanted with 15 mg of placebo (placebo 15 mg); and (viii) EAGD mice implanted with 15 mg of DHT (DHT 15 mg). There were 12 mice in each group, but a few mice died during the experiment. The final mice numbers were 12 (Con), 10 (EAGD), 10 (placebo 1.5 mg), 11 (DHT 1.5 mg), 10 (placebo 5 mg), 11 (DHT 5 mg), 9 (placebo 15 mg), and 11 (DHT 15 mg). Mice were housed in specific pathogen-free conditions at the Laboratory Animal Center (Xi'an Jiaotong University Health Science Center, China). Animals were maintained throughout the study in a controlled environment (23°C and 60% humidity) and a 12-hour light/dark cycle. Water and standard lab chow were available ad libitum. All animal experiments were performed in accordance with protocols approved by the animal Care and Use Committee of Xi'an Jiaotong University.
DHT pellet implantation
Ninety-day release pellets of DHT at doses of 1.5, 5, and 15 mg and matching placebos were purchased from Innovative Research of America. Mice were anesthetized with 10% ketamine (0.03–0.05 mL/10 g) by intraperitoneal injection. A small incision was made in the skin between the neck and shoulder blades (12). A week before immunization, the DHT or placebo pellet was implanted subcutaneously, and the wound was sutured afterwards. Mice were immediately warmed after the intervention to prevent hypothermia resulting from anesthesia, and they were watched carefully each day to assess eating and drinking behavior, as well as wound healing.
Immunization of mice with Ad-TSHR289
Induction of Graves' disease with an adenovirus expressing the human TSHR A-subunit (Ad-TSHR289) was performed as described previously, with the null adenovirus as the control (13). In brief, Ad-TSHR289 and a control adenovirus were propagated in HEK293 cells and purified via ion-exchange column chromatography. The viral particle concentration was determined by measuring the absorbance at 260 nm. Immunization with recombinant adenovirus in mice has been described previously (14). Briefly, one group of mice was injected with the null adenovirus intramuscularly and the other seven groups were injected with Ad-TSHR289 in 50 μL of phosphate buffered saline (PBS) containing 109 particles. Each of the six groups was treated with different doses of DHT or the matching placebo. Each animal was injected three times at three-week intervals. Mice were sacrificed four weeks after the third immunization. Then, blood was drawn, and the spleen and thyroid tissue were removed for further analysis. Figure 1 shows the timeline for DHT implantation and immunization with Ad-TSHR289.
Diagram illustrating the timeline for human thyrotropin receptor A-subunit (Ad-TSHR289) immunization and 5α-dihydrotestosterone (DHT) pretreatment.
Examination of serum thyrotropin receptor antibodies and thyrotropin-binding inhibition
The level of thyrotropin receptor antibodies (TRAb) in undiluted serum (100 μL) was determined using a commercial kit following the manufacturer's protocol (Medipan GmbH), and thyrotropin-binding inhibition (TBI) was calculated using the formula [1 – (TSH binding in the presence of test serum-nonspecific binding)/(TSH binding in the presence of normal serum-nonspecific binding)] ×100 (13). The levels of TRAb in diluted serum (35 μL and 10 μL) were also measured in the following groups: mice receiving 5 mg of DHT (the middle dose), EAGD, and Con, respectively.
Detection of serum total thyroxine and free thyroxine
Total thyroxine (TT4) and free thyroxine (fT4) levels in undiluted serum (50 μL for TT4 or fT4) were measured with a radioimmunoassay kit. Detection was performed according to the manufacturer's protocol (Beijing Atomic Hi-Tech Co. Ltd).
Histological examination of the thyroid
Thyroid tissues were fixed in 10% buffered formalin and paraffin embedded, and serial sections were stained with hematoxylin and eosin. Sections were analyzed with an Olympus Cue-2 image analysis system connected to an Olympus compound microscope.
Concentration of serum DHT determined by enzyme-linked immunosorbent assay
To determine the effectiveness of the pellets in increasing circulating hormone levels, serum samples (50 μL) were taken from mice from each group and measured with a DHT enzyme-linked immunosorbent assay kit (Alpha Diagnostic International) according to the manufacturer's instructions.
Preparation of single-cell suspensions
Spleen cells were gently dispersed into a single-cell suspension through a nylon mesh. Red blood cells were eliminated with red blood cell lysis buffer (Tiangen Biotech Co. Ltd). Lymphocytes were washed and suspended in RPMI 1640 (HyClone) at a final concentration of 1 × 107 cells/mL.
Antibodies
The following monoclonal antibodies were used: antimouse CD4 FITC, antimouse IFN-γ APC, antimouse IL-2 PerCP-Cy5.5, antimouse IL-4 PE, antimouse IL-10 PerCP-Cy5.5, Alexa Fluor® 647 antimouse IL-9, and antimouse/rat IL-17A PE for each of the detected cytokines secreted by Th1, Th2, T helper 9 (Th9), and T helper 17 (Th17) cells. All the antibodies and appropriate isotype controls for flow cytometric analysis were purchased from eBioscience, except for Alexa Fluor® 647 antimouse IL-9 (Biolegend).
Intracellular staining of cytokines and flow cytometric analysis
To quantify any cytokines present, spleen cells were suspended in RPMI 1640 medium with 10% fetal calf serum (Hangzhou Sijiqing Biological Engineering Materials Co. Ltd), transferred to each well of a 48-well plate, and stimulated with a PMA/ionomycin mixture (250×, 4 μL/mL final concentration; eBioscience) for 6 h in the presence of BFA/monensin mixture (250×, 4 μL/mL final concentration; eBioscience) for intracellular staining and flow cytometric analysis of IFN-γ, IL-2, IL-4, IL-10, IL-9, and IL-17. Briefly, cells were stained with antimouse CD4 FITC for 30 min, incubated with IC Fixation buffer (eBioscience) for 20 min, washed twice with a permeabilization buffer, and stained with antimouse antibodies. Flow cytometric analysis was performed as above.
Statistical analysis
Values are presented as the means ± standard error of the mean. Statistical differences between two groups were analyzed using an unpaired t-test. For nonparametric mean values, a Mann–Whitney U-test was used to compare two independent samples. Statistical differences between multiple treated groups were determined by one-way analysis of variance, if there were significant differences among groups, followed by a Duncan multiple range test. A p-value of <0.05 was considered statistically significant. Statistical analyses were performed with GraphPad Prism v5.
Results
Development of EAGD
Signs of hyperthyroidism were observed in EAGD mice following immunization, including irritability, greed, and weight loss. Other signs included positive TBI data that reached 100%, elevated TT4 and fT4 levels relative to the control, enlarged thyroid glands, larger thyroid follicles with epithelial intrusions into the follicular lumen, and decreased colloid in the majority of mice upon examination of thyroid histology. Lymphocytes were not found in the thyroid gland.
Concentrations of serum DHT
Serum DHT levels in androgen-pretreated mice were markedly increased, indicating successful DHT supplementation (Fig. 2).
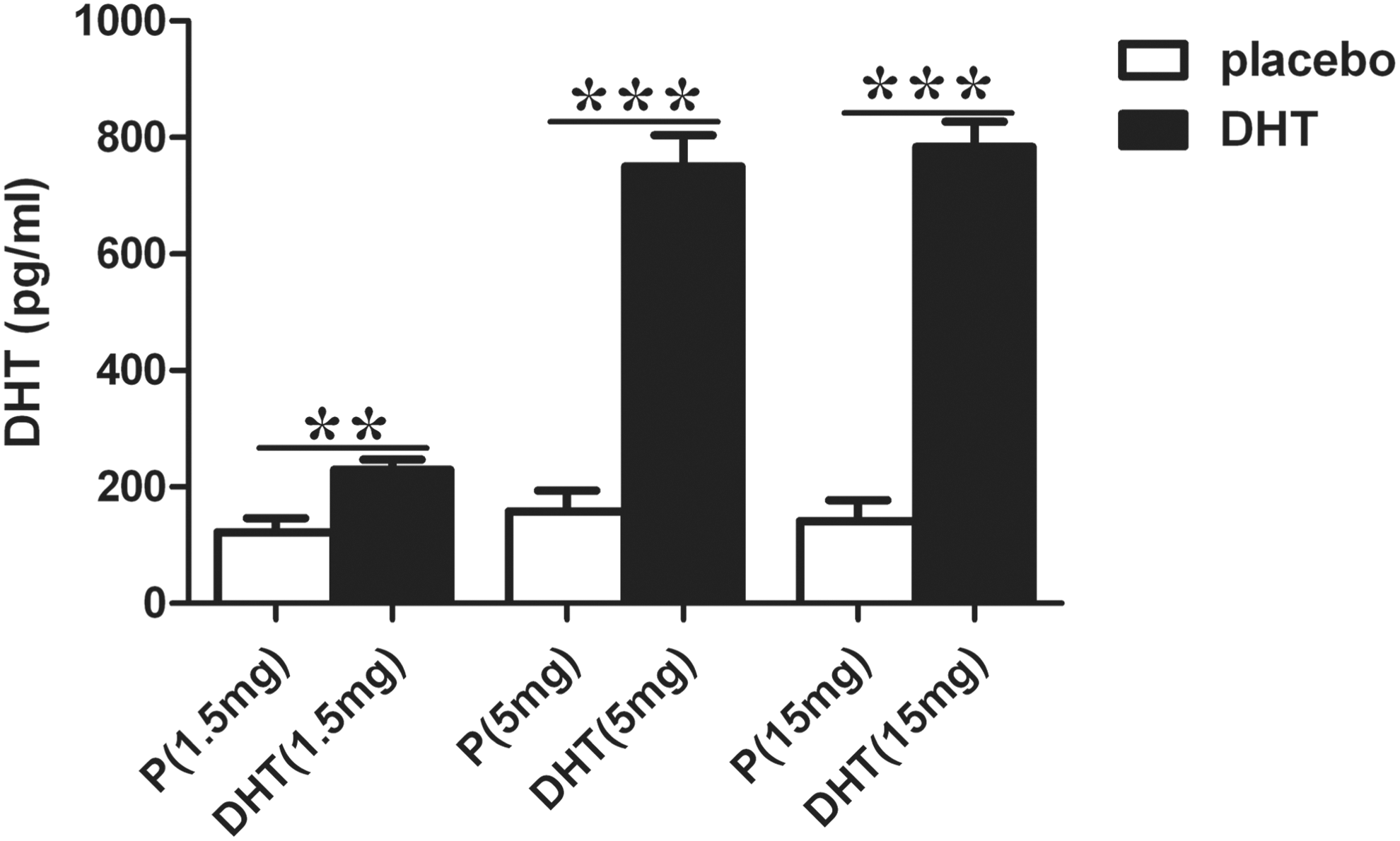
Serum DHT levels of immunized mice supplemented with different doses of DHT or placebo. Values are expressed as means ± standard error of the mean (SEM). Animal numbers were 12 (Con), 10 (EAGD), 10 (placebo 1.5 mg), 11 (DHT 1.5 mg), 10 (placebo 5 mg), 11 (DHT 5 mg), 9 (placebo 15 mg), and 11 (DHT 15 mg). The white bars show placebo groups, and DHT-pretreated groups are represented as black bars. Concentrations of DHT in mice pretreated with DHT were significantly higher than in mice in the placebo groups (**p < 0.01; ***p < 0.001). P, placebo.
DHT ameliorates the development of GD in female BALB/c mice
Mice were immunized three times with Ad-TSHR289. Four weeks after the third immunization, histological examination of the thyroid was analyzed, serum TRAb levels were determined with a commercial kit, and TBI was calculated. After DHT treatment, the thyroid follicles were smaller, and thyroid follicular cells were reduced (Fig. 3). In a dose-independent manner, TBI percentages in undiluted serum (100 μL) were slightly reduced in the groups pretreated with DHT, but there was no statistical difference compared to placebo (Fig. 4A). In subsequent analyses, TBI activities were measured in the group with an intermediate dose of DHT (i.e., 5 mg), with lower amounts of serum (35 μL and 10 μL). A decreasing trend was identified in the group treated with 5 mg DHT, but a statistical difference could not be found (Fig. 4B). There was a marked fall in TT4 levels in the presence of DHT compared with the placebo group, and this effect was not dose dependent (Fig. 4C). fT4 was also examined, because androgens can reduce TT4 levels by reducing the binding of thyroxine by thyroxine-binding globulin. The results showed a significantly lower fT4 level in mice pretreated with 5 and 15 mg of DHT. As for the smaller dose of 1.5 mg of DHT, a small reduction was observed, but it did not reach statistical significance (Fig. 4D).

Histological examination of the thyroid. Con, normal mice injected with a null adenovirus; EAGD, mice immunized with Ad-TSHR289; Placebo, placebo 5 mg; DHT, 5α-dihydrotestosterone 5 mg. Histological examination revealed larger thyroid follicles with epithelial intrusions into the follicular lumen and less colloid in EAGD mice. The thyroid follicles were smaller and thyroid follicular cells were reduced after DHT treatment.
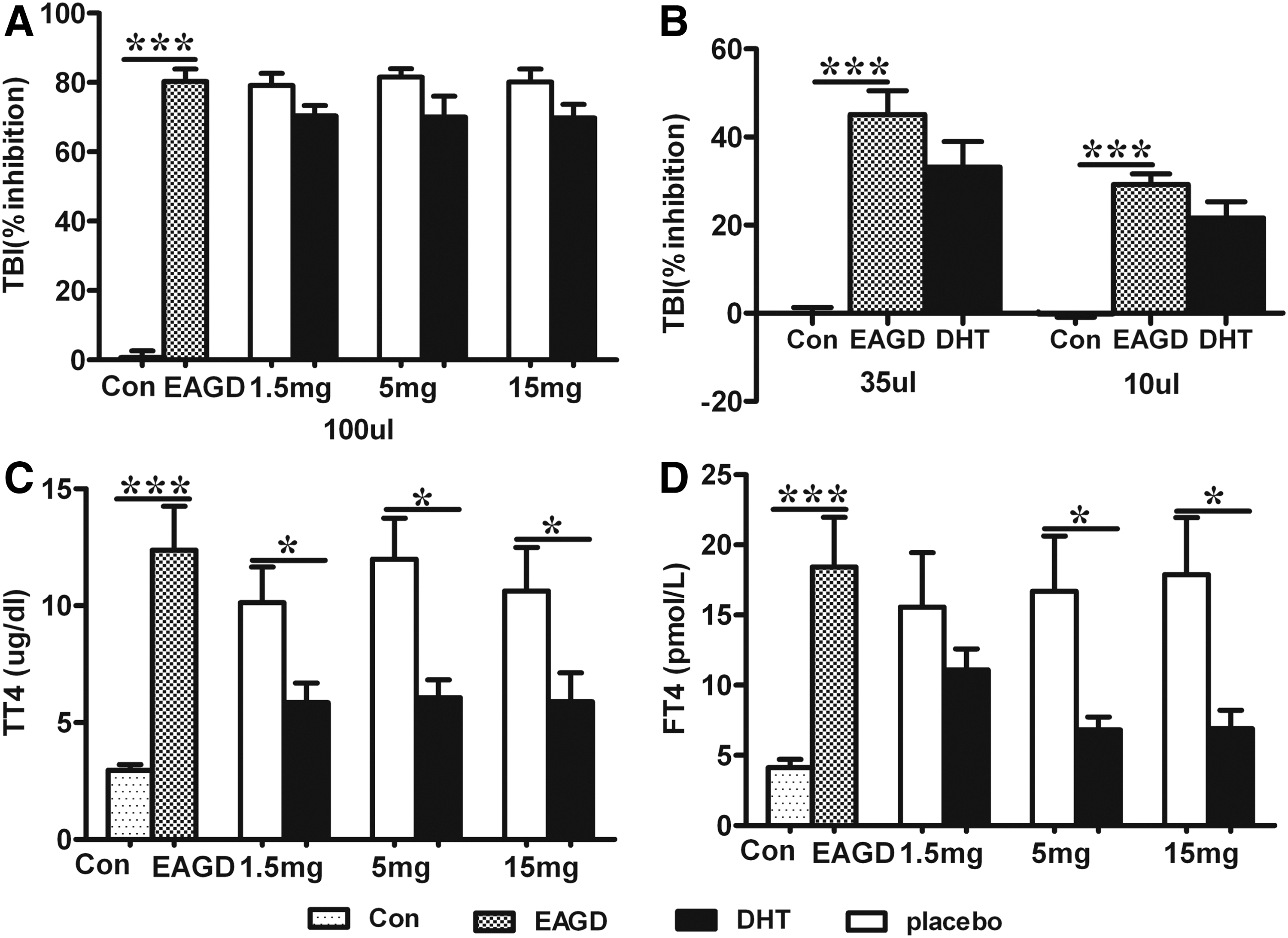
TBI and serum levels of thyroid hormones were determined in mice. Values are expressed as means ± SEM. Animal numbers were 12 (Con), 10 (EAGD), 10 (placebo 1.5 mg), 11 (DHT 1.5 mg), 10 (placebo 5 mg), 11 (DHT 5 mg), 9 (placebo 15 mg), and 11 (DHT 15 mg). The white bars show placebo groups, and DHT-pretreated groups are represented as black bars. TBI, TT4, and fT4 were significantly elevated in EAGD mice (Con vs. EAGD; p < 0.001). A downtrend for TBI was observed in the mice pretreated with three different doses of DHT in 100 μL of undiluted serum (
Shifting of Th1 to Th2 cells in response to DHT
Th1 cells are the most important mediators of autoimmune inflammation, and the shifting of Th1 to Th2 cells could prevent the occurrence of autoimmune diseases (15). This study investigated whether there were changes in Th1 and Th2 cell numbers after immunization, and whether DHT was capable of inhibiting the autoimmune Th1 response and inducing Th2 cytokine profiles. Th1 cells are characterized by secretion of IFN-γ and IL-2, while IL-10 and IL-4 are produced by Th2 cells as anti-inflammatory factors (16). Intracellular staining was performed after stimulation with a PMA/ionomycin mixture, and CD4+ splenic T lymphocytes were analyzed with flow cytometry.
First, the percentages of Th1 cells in CD4+ T cells were examined, and the analyses revealed that the proportion of CD4+IL-2+ and CD4+IFN-γ+ T cells increased significantly in immunized mice. The percentage of CD4+IL-2+ cells reduced significantly in all three DHT-treated groups (Fig. 5B). In addition, 5 and 15 mg of DHT reduced the proportion of CD4+IFN-γ+ T lymphocytes, which was consistent with the effect of DHT on thyroid function (Fig. 5B). Second, Th2 cytokines such as IL-4 and IL-10 were also detected. In contrast to Th1 cells, the percentages of CD4+IL-4+ and CD4+IL-10+ cells decreased after immunization (Fig. 5B). Taken together, the results indicate that there was an increase in Th1 cell numbers and a reduction in the number of Th2 cells after immunization, but DHT could switch these phenomena to maintain the balance between Th1/Th2 cells, primarily by suppressing the Th1 response (Fig. 5).
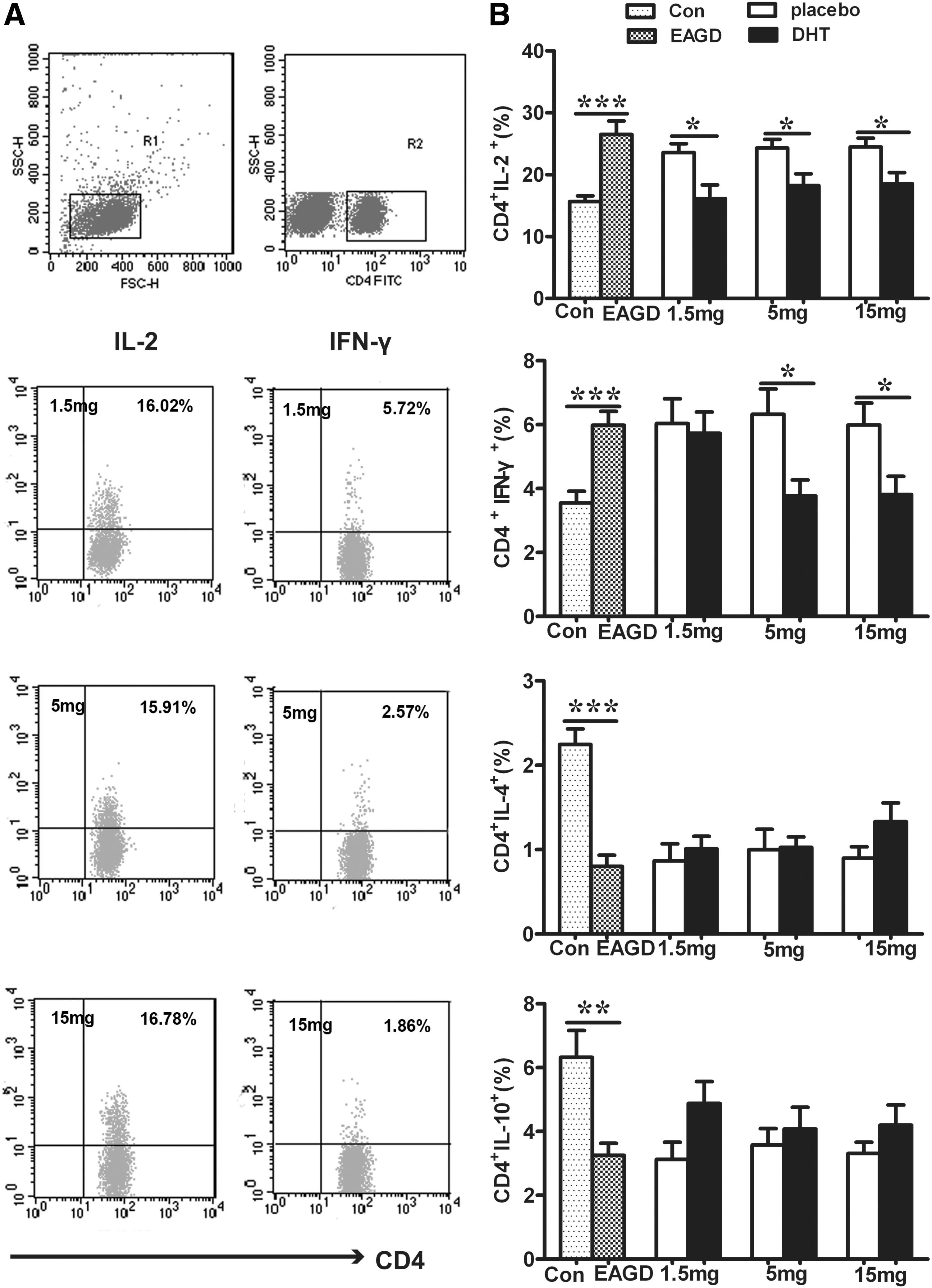
Numbers of T helper 1 (Th1) and T helper 2 (Th2) cells were determined by flow cytometry. Mice were pretreated with DHT or a placebo a week before immunization. Splenic cells were harvested four weeks after the third immunization. Values are expressed as means ± SEM. Animal numbers were 12 (Con), 10 (EAGD), 10 (placebo 1.5 mg), 11 (DHT 1.5 mg), 10 (placebo 5 mg), 11 (DHT 5 mg), 9 (placebo 15 mg), and 11 (DHT 15 mg). The left photographs present representative Th1 cells in the DHT-pretreated groups (
Influence of DHT treatment on Th9 and Th17 cells
Th17 cells are thought to play a more active role than Th1 cells in the process of GD (5,17), while Th9 cells have been thought to play a pleiotropic function in the autoimmune response. This study investigated whether there were changes after immunization, and whether DHT was able to modulate IL-9 and IL-17 levels in this animal model of GD. The proportion of CD4+IL-9+ and CD4+IL-17+ T cells was examined in mouse spleens using flow cytometry. After immunization with Ad-TSHR289, the percentage of Th9 cells was markedly reduced in EAGD mice, but a statistically significant difference was not observed between the DHT and placebo groups (Fig. 6B, left panel). An alteration in IL-17 levels was not found in immunized mice or in DHT-pretreated mice in this study, suggesting that both immunization with Ad-TSHR289 and immune protection provided by DHT are not likely to be associated with Th17 cells in BALB/c mice (Fig. 6B, right panel).
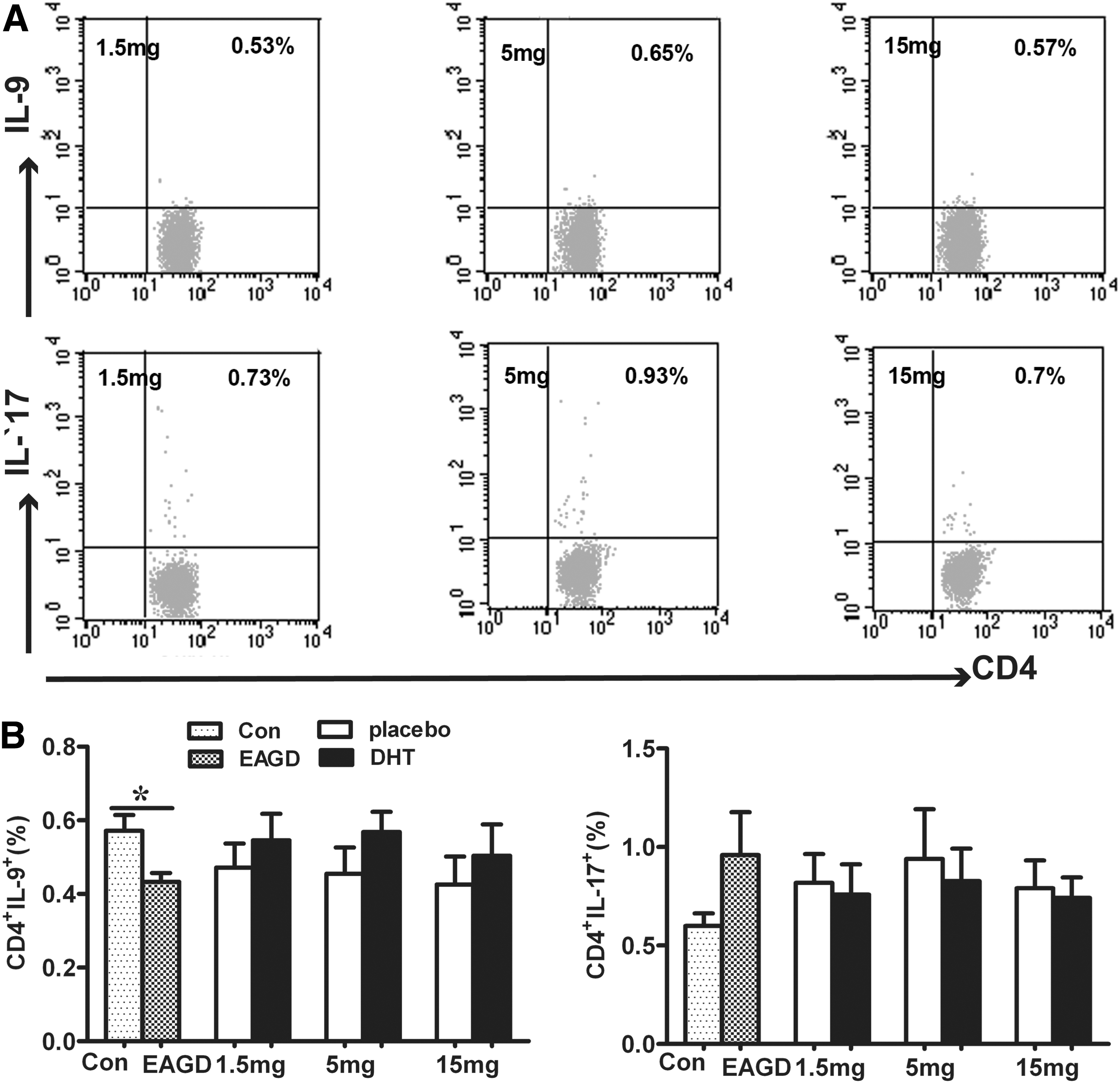
Examination of T helper 9 (Th9) and T helper 17 (Th17) cells in the CD4+ T cell population from the spleen of mice. Pictures represent percentages of CD4+IL-9+ Th9 cells and CD4+IL-17+ Th17 cells in DHT-implanted mice (
Discussion
Immune responses have been classified into two types. Cellular immunity is mainly induced by Th1 cells, while Th2 cells are primarily responsible for humoral immunity (4). GD is generally considered an autoimmune disease mediated predominately by Th2-type cells. However, a number of studies on cytokine profiles have shown that GD is characterized by a mixed Th1 and Th2 immune response (18,19). In other studies, a Th1 immune response has been found to play a key role in mouse models of GD immunized by TSHR antigen (20,21). Consequently, the influence of the Th1/Th2 cytokine production in the development of GD is still controversial, and the immune responses are different in distinct animal models (4).
This study induced a BALB/c mouse model of GD using Ad-TSHR289, and pretreated the immunized mice with three doses of DHT. The results show that TBI and serum levels of TT4 and fT4 were highly elevated in immunized mice compared with the control group. Furthermore, there were higher percentages of CD4+IFN-γ+ and CD4+IL-2+ cells but lower CD4+IL-4+, CD4+IL-10+, and CD4+IL-9+ cell levels in the immunized group compared with the control. In groups pretreated with DHT, the thyroid hormone levels decreased dramatically compared with the placebo group, which indicates that DHT could ameliorate the severity of GD in female BALB/c mice. Significant reductions in IFN-γ and IL-2 production, but not IL-17 levels, were observed in DHT-pretreated mice, suggesting that DHT might influence the production of cytokines in splenic cells.
It is well accepted that there are at least two ways through which TRAb result in functional consequences in autoimmune thyroid diseases. One consists in TSH stimulating antibodies (TSAb), which are thought to be responsible for the development of hyperthyroidism. Another consists of TSH blocking antibodies (TBAb), which inhibit TSH stimulation and result in hypothyroidism (22). The present results show that TBI percentages in undiluted serum (100 μL) were slightly reduced in the groups pretreated with DHT, but this did not reach statistical significance. In order to characterize TBI activity in the group treated with the intermediate dose of 5 mg DHT further, TBI percentages were measured in two smaller volumes of serum (35 μL and 10 μL), respectively, and the results were similar to those detected in 100 μL of undiluted serum. Although there is no statistical difference in TBI between the DHT and placebo groups, there is a possibility that DHT may have some effect on TSAb or TBAb. Unfortunately, the study failed to measure TSAb and TBAb due to the limited amounts of serum.
GD is thought to be a failure of the normal immune tolerance mechanism due to pathogenic T cells (23). Helper CD4+ T cells have been classified into several subsets with distinct cytokine secretion profiles. Th1 cells are characterized by the secretion of IL-2, IFN-γ, and TNF-α, and Th17 cells are thought to produce IL-17 (24). In contrast, IL-4, IL-5, IL-10, and IL-6 are secreted by Th2 cells (25). Recently, a novel subset of CD4+ T cells, Th9 cells, has been characterized, and these cells primarily produce IL-9 (26). Cytokines such as IFN-γ, IL-2, and IL-17A are generally known as pro-inflammatory cytokines associated with inflammatory autoimmune diseases (27). On the other hand, the anti-inflammatory cytokines IL-4 and IL-10 play an important role in the prevention of autoimmune responses. It has been shown that Th9 cells play a role in the induction of experimental autoimmune encephalomyelitis (EAE) (28). In contrast, another study has shown that Th9 cells may enhance the suppressive capacity of regular T cells, and the absence of the IL-9 signaling pathway can lead to greater severity in a mouse model of EAE (29). Therefore, there are some conflicting data concerning the role of Th9 cells in immunity. In the present study, Th9 cell numbers were reduced in immunized mice, while increased slightly with the treatment of DHT.
Protection against autoimmunity mediated by endogenous testosterone has been demonstrated in a number of castrated male animal models, including thyroiditis, EAE, diabetes in non-obese mice, and arthritis (30 –33). In addition, exogenous testosterone treatment can alleviate numerous autoimmune diseases in humans and animal models (10,33 –36). In vitro, T cells from female mice stimulated with auto-antigens produced higher levels of IL-10 and lower levels of IFN-γ in the presence of testosterone (37). Splenocytes from dihydrotestosterone-treated female mice with EAE were found to secrete higher levels of the anti-inflammatory cytokine IL-10 and lower levels of IL-12 (38). Moreover, testosterone has been shown to prolong survival by delaying the development of lupus in a mouse model (39,40). Recently, an adrenal steroid, dehydroepiandrosterone (DHEA), has been used as a therapeutic agent for systemic lupus erythematosus in humans (41). DHEA can be converted into androgens and estrogens in peripheral tissues, with mild intrinsic androgenic properties (42). These studies support the hypothesis that androgens play an important role in the prevention of autoimmune diseases, regardless of whether the androgen is endogenous or exogenous and treatment is in vivo or in vitro. So far, however, the effects of androgens on humans or animals with GD are still unclear.
This study has found that DHT pretreatment can alleviate the severity of GD and regulate the balance between Th1 and Th2 cells in female BALB/c mice by downregulating pro-autoimmune Th1 cells. This protective influence was more obvious with doses of 5 and 15 mg of DHT. These results indicate that DHT is an immune regulator that protected these animals from GD.
Footnotes
Acknowledgments
We would like to thank Chunrong Chen (University of California, Los Angeles, CA) for providing us the plasmid expressing human TSHR289 (psv2-neo-ECE).
This work was supported by the Natural Science Foundation of China (grant number 81170729, 30972701); basic scientific research business expenses of Xi'an Jiaotong University (grant number xjj2012164).
Author Disclosure Statement
The authors disclose that no competing financial interests exist.