
Abstract
Background:
Peroxisome proliferator-activated receptor γ (PPARγ) is a transcription factor that regulates the expression of multiple target genes involved in several metabolic pathways as well as in inflammation. The expression and cell localization of caveolin-1 (Cav-1), thyroperoxidase (TPO), and dual oxidase (DUOX), involved in extracellular iodination, is modulated by Th1 cytokines in human normal thyroid cells and in Hashimoto's thyroiditis (HT).
Objectives:
The objectives of this study were (i) to analyze the PPARγ protein and mRNA expression at the follicular level in HT versus controls in correlation with the one of Cav-1; (ii) to study the effects of Th1 cytokines on PPARγ and catalase expression in human thyrocyte primary cultures; and (iii) to study the effects of pioglitazone, a PPARγ agonist, on thyroxisome components (Cav-1, TPO, DUOX) and on catalase, involved in antioxidant defense.
Results:
Although the global expression of PPARγ in the whole gland of patients with HT was not modified compared with controls, there was great heterogeneity among glands and among follicles within the same thyroid. Besides normal (type 1) follicles, there were around inflammatory zones, hyperactive (type 2) follicles with high PPARγ and Cav-1 expression, and inactive (type 3) follicles which were unable to form thyroxine and did not express PPARγ or Cav-1. In human thyrocytes in primary culture, Th1 cytokines decreased PPARγ and catalase expression; pioglitazone increased Cav-1, TPO, and catalase expression.
Conclusion:
PPARγ may play a central role in normal thyroid physiology by upregulating Cav-1, essential for the organization of the thyroxisome and extracellular iodination. By upregulating catalase, PPARγ may also contribute to cell homeostasis. The inhibitory effect of Th1 cytokines on PPARγ expression may be considered as a new pathogenetic mechanism for HT, and the use of PPARγ agonists could open a new therapeutic approach.
Introduction
P
PPARγ agonists are well-known modulators of the inflammatory process (3) and of the interferon (IFN)-induced chemokine secretion in normal thyroid cells and in Graves' disease (GD) (5,6). However, the Th1 and Th2 cytokine effects on PPARγ expression have not been studied in the thyroid gland. Of note, it has been shown that Th1 cytokines downregulate PPARγ, for example in adipose tissue (7,8) and in human biliary cells (9), whereas Th2 cytokines upregulate PPARγ in glial cells (10) and in human biliary cells (9).
The thyroid gland is a good model to study the effects of Th1 and Th2 cytokines. Indeed, both cellular and humoral immunity have a role in the pathogenesis of autoimmune thyroid diseases. Hashimoto's thyroiditis (HT) associated with hypothyroidism is mainly due to a cytotoxic immune reaction against thyroperoxidase (TPO) and thyroglobulin (Tg). GD is associated with hyperthyroidism due to the activation of the thyrotropin (TSH) receptor by specific stimulating autoantibodies (11). GD is considered as a Th2 dominant disease, although Th1 cytokines may prevail at the onset of its development (6).
Thyroid hormone synthesis and the maintenance of euthyroidism is a very controlled cell process (for a review, see Colin et al. (12)). It requires the transfer of iodide (I–), circulating in the blood, into the colloid by different transporters. Dual oxidase (DUOX) generates H2O2, which is required at the apical border to oxidize iodide and hence to form monoiodotyrosine (MIT) and diiodotyrosine (DIT) incorporated in Tg. The combination of MIT and DIT leads to the production of triiodothyronine (T3) and thyroxine (T4), released in the circulation after endocytosis and hydrolysis of the iodinated Tg. DUOX and TPO are two essential proteins for thyroid hormone biosynthesis, which gather at the apical cellular membrane forming the thyroxisome, a complex entirely dedicated to safe and efficient thyroid hormone synthesis. Caveolin-1 (Cav-1), by creating raft-like membrane structures, is likely a required element to form the thyroxisome, as first proposed by Song et al. (13). As shown in the Cav-1 knockout mouse model, Cav-1 deficiency is associated with the cytoplasmic localization of DUOX and TPO, with the production of excessive intracytoplasmic H2O2, and thus with increased oxidative stress (14).
Cytokines have been shown to have an effect on the different partners of the thyroxisome. In HT, the immunological environment is mostly composed by Th1 cytokines, among which IFN-γ, interleukin (IL)-1α and β, IL-2, and tumor necrosis factor (TNF-α, TNF-β) are predominant. Th1 cytokines appear to play a key role for the inhibition of thyrocyte function in HT. Indeed, sodium–iodide symporter (NIS), TPO, and DUOX are downregulated by Th1 cytokines (15,16). Cav-1 is also dramatically downregulated in HT, resulting in a cytoplasmic mislocalization of TPO and DUOX, and intracellular iodination. It leads to pronounced oxidative stress with a high apoptosis rate (17), as also previously shown in Cav-1 knockout mice (14), and in Pendred's syndrome (18). In HT, this downregulation of Cav-1 is due, at least partly, to Th1-cytokines (IL-1α and IFN-γ) (17). The resulting oxidative stress is insufficiently compensated by the cellular antioxidant defenses and thus leads to cell apoptosis, follicular destruction, and hypothyroidism (17).
The pathways implicated in Cav-1 regulation in normal thyroid cells remain unknown. However, it has been shown that PPARγ upregulates Cav-1 gene expression in different models, for example human macrophages and mouse hepatocytes (19,20).
The aims of this study were first to analyze the PPARγ expression in HT and to correlate it with that of Cav-1 and the functional status of the follicles in term of T4 synthesis. Second, the effects of Th1 cytokines on PPARγ and catalase expression in human primary thyrocytes were analyzed. Third, the effects of pioglitazone, a PPARγ agonist, were investigated in human primary thyroid cells on thyroxisome components (Cav-1, TPO, and DUOX) and on antioxidant defense (catalase) of the thyrocytes. The understanding of the intracellular mechanism leading to thyrocyte destruction in HT and the demonstration of the beneficial impact of PPARγ agonists could open new therapeutic approaches.
Materials and Methods
Thyroid samples
Tissue samples from seven HT patients were obtained through the Biolibrary of the Université Catholique de Louvain (Brussels, Belgium). The patients underwent surgery because of suspicious nodules detected by echography, but they were finally diagnosed as HT. Thyroid samples removed in the paranodular zone of multinodular goiters, considered as normal (n = 7), were used as control samples. The surgical samples were obtained with the informed consent of patients.
The clinical data of the HT and control patients are shown in Supplementary Table S1 (Supplementary Data are available online at
Preparation of tissue samples for light microscopy and immunohistochemistry
The thyroids were fixed in 4% paraformaldehyde in phosphate buffered saline (PBS) for 24 hours and embedded in paraffin. Immunostaining was performed on 5-μm-thick paraffin sections. Sections were dewaxed and rehydrated, and endogenous peroxidases were quenched with 1% H2O2 for 15 min. For the antigen retrieval, tissue sections were pretreated in a microwave oven in citrate buffer (pH 6.6) for one 3 min cycle at 750 W, followed by four cycles of 3 min 30 sec, each at 350 W. All sections were washed with PBS supplemented with 1% bovine serum albumin (PBS-BSA) and thereafter incubated in 5% PBS-BSA containing 2% non-immune goat serum at room temperature (RT) for 1 h. Sections were incubated at RT for 24 h with a PPARγ polyclonal rabbit antibody (1:250; SAB4502262; Sigma, Saint-Louis, MO) for 3 h with a Cav-1 polyclonal rabbit antibody (1:400; BD610059; BD Transduction Laboratory, Franklin Lakes, NJ) for 3 h with a catalase monoclonal mouse antibody (1:200; C-0979; Sigma), or overnight with a T4 monoclonal mouse antibody (1:400; MBS592059; MyBioSource, San Diego, CA).
The binding of antibodies was detected using a secondary antibody conjugated to a peroxidase labeled polymer for 1 h at RT (1:50; 32260; Pierce® goat anti-rabbit; ThermoScientific, Rockford, IL; or 1:50; 32230; Pierce® goat anti-mouse; ThermoScientific). The peroxidase activity was revealed with 3-3′ diaminobenzidine tetrahydrochloride (DAB; Sigma). Sections were counterstained with Mayer's hematoxylin, and rinsed and mounted in Faramount aqueous mounting medium (DakoCytomation, Carpinteria, CA). To verify the binding specificity, some sections were incubated by omitting the primary antibody.
The quantification of the immunolabeling was made by ImageJ software developed by the National Institute of Health (NIH; Bethesda, MD).
Primary cell cultures
Thyroid samples (n = 5) were obtained after surgery of multinodular goiters (Cliniques Universitaires St. Luc, Brussels, Belgium). The samples were removed in the paranodular zone, considered as normal. These tissues were isolated as previously described (21) and suspended in modified Earle's medium (BRL-Gibco, Paisley, United Kingdom), containing 5% newborn calf serum (Gibco), penicillin (100 IU/mL; Gibco), streptomycin (100 IU/mL; Gibco), glutamine (2.4 mM; Sigma), and fungizone (2.5 μg/mL; Gibco). The cells were plated in 12-well plates and cultured in a humidified atmosphere (37°C, 5% CO2) with 1 mIU/mL of TSH (Sigma) for one week.
Cytokine effects
Cells were then incubated or not for three additional days, with 2 ng/mL of recombinant murine IL-1α and 10 ng/mL of recombinant murine IFN-γ (RD systems, Abingdon, United Kingdom) in modified Earle's medium containing 0.5% newborn calf serum, the Th1 cytokines inducing a downregulatory effect on TPO, DUOX (16), and Cav-1 (17) at these doses.
Pioglitazone effects
Cells were incubated or not for 24 h with 15 μM of pioglitazone (Sigma) in modified Earle's medium containing 0.5% newborn calf serum. The pioglitazone concentration was chosen according to previous studies by Antonelli et al. (5,6,22).
RNA purification and quantitative reverse transcription polymerase chain reaction
mRNA levels of PPARγ, cav-1, TPO, DUOX1, DUOX2, and catalase were measured by quantitative reverse transcription polymerase chain reaction (RT-PCR). These experiments were performed, as previously described (17), using the iQ5 (BioRad, Hemel Hempstead, United Kingdom) as follows: 30 cycles at 95°C for 1 min, followed by 40 cycles at 95°C for 15 sec, the appropriate annealing temperature (see Supplementary Table S2), for 45 sec, and at 81°C for 15 sec. The amplified level of the target gene was normalized against that of β-actin. Amplification of genomic DNA was avoided by using primers spanning two adjacent exons.
Western blotting
Cells were homogenized in Laemmli buffer containing a protease inhibitor cocktail (Roche, Basel, Switzerland). Western blotting and its quantification were performed, as previously described (17). All the characteristics of primary antibodies are summarized in Supplementary Table S3. All values are expressed as mean ± standard error of the mean.
Cav-1 and PPARγ immunofluorescence
Thyrocytes were cultured on cover glasses in appropriate medium. They were fixed for 10 min in 4% paraformaldehyde, rinsed with PBS, and blocked for 1 h in a PBS saponin 0.1%/BSA 5% solution at 37°C. Cells were then incubated overnight (ON) at 4°C with a Cav-1 monoclonal mouse primary antibody (1:100; SAB4200216; Sigma) diluted in PBS saponin 0.1%/BSA 1%. The co-staining was realized by adding a mix of the Cav-1 monoclonal mouse primary antibody (1:100; SAB4200216; Sigma) and the PPARγ polyclonal rabbit antibody (1:250; SAB4502262; Sigma) diluted in PBS saponin 0.1%/1% BSA. After being washed in PBS saponin 0.1%, the cells were incubated for 1 h at 37°C with an Alexa fluor (488) secondary antibody (1:300; A11029; Invitrogen, Genk, Belgium). For the co-staining, a mix was introduced containing the same Alexa fluor 488 secondary antibody (1:300; A11029; Invitrogen) and the Alexa fluor 568 secondary antibody (1:300; A11036; Invitrogen), which was then incubated for 5 min with Hoechst at a dilution of 1:5000. Cover slides were mounted in fluorescent mounting medium (DakoCytomation) for microscopic observation.
A Axio Zeiss microscope (Zeiss, Zaventem, Belgium) was used, and the quantification of the fluorescence was made by ImageJ software developed by the NIH.
Statistical analysis
RT-PCR and Western blotting results were normalized against levels of β-actin. Statistical analysis was performed using a t-test or a Mann–Whitney test when results did not pass the normality test (GraphPad, San Diego, CA). A p-value of <0.05 was considered statistically significant.
Results
Immunohistochemical detection reveals a strong follicular heterogeneity of T4 synthesis in HT
In control thyroids, the majority of the follicles contained T4 homogeneously distributed in the colloid. They were large with a flat or cuboidal epithelial layer (Fig. 1A). These normal follicles were classified as type 1.

Immunohistochemical detection of thyroxine (T4), peroxisome proliferator-activated receptor γ (PPARγ), caveolin-1 (Cav-1), and catalase in a normal thyroid gland and Hashimoto's thyroiditis (HT). In control thyroids (
Hashimoto's thyroids were modified by inflammatory infiltrates, which, in some samples, invaded very large parts of the glandular parenchyma. Zones of type 1 follicles were still observed, their surfaces being variable among HT patients. At the periphery of inflammatory zones, two other types of follicles were observed: type 2 follicles, which were usually small and contained T4 in their lumina, and type 3 follicles, which were small but without T4 (Fig. 1B). In type 3 follicles located inside the inflammatory infiltrates, T4 was also observed in the cytoplasm rather than in the colloid, suggesting intracellular iodination (Fig. 1C).
Immunohistochemical detection reveals a strong follicular heterogeneity regarding PPARγ, Cav-1, and catalase expression
By analyzing the mRNA and protein PPARγ expression by RT-PCR and Western blotting, no significant differences were observed between HT samples and control glands (Supplementary Fig. S1A and B). The same applied to the quantification of the immunohistochemical labeling of PPARγ, which did not show any differences between HT and controls (Supplementary Fig. S1C). However, these results were obtained on whole samples, which, in HT, are very heterogeneous regarding the morphological and functional phenotype of the follicles that were classified in three types on histological sections.
Surrounding the inflammatory zones of Hashimoto's glands, the type 2 follicles expressing T4 (Fig. 1D) showed very high nuclear expression of PPARγ (Fig. 1E). The same follicles also showed strong expression of Cav-1 (Fig. 1F) and catalase (Fig. 1G). The expression of PPARγ, Cav-1, and catalase was low or absent in type 3 follicles, having lost the ability for T4 synthesis.
Correlation between the expression of PPARγ and Cav-1 in the three types of follicles in HT
In normal type 1 follicles, T4 was detected in the entire colloid (Fig. 2A). PPARγ was detected in a few nuclei (Fig. 2B), and Cav-1 was located at the apical pole of the thyrocytes where normal iodination takes place (Fig. 2C).
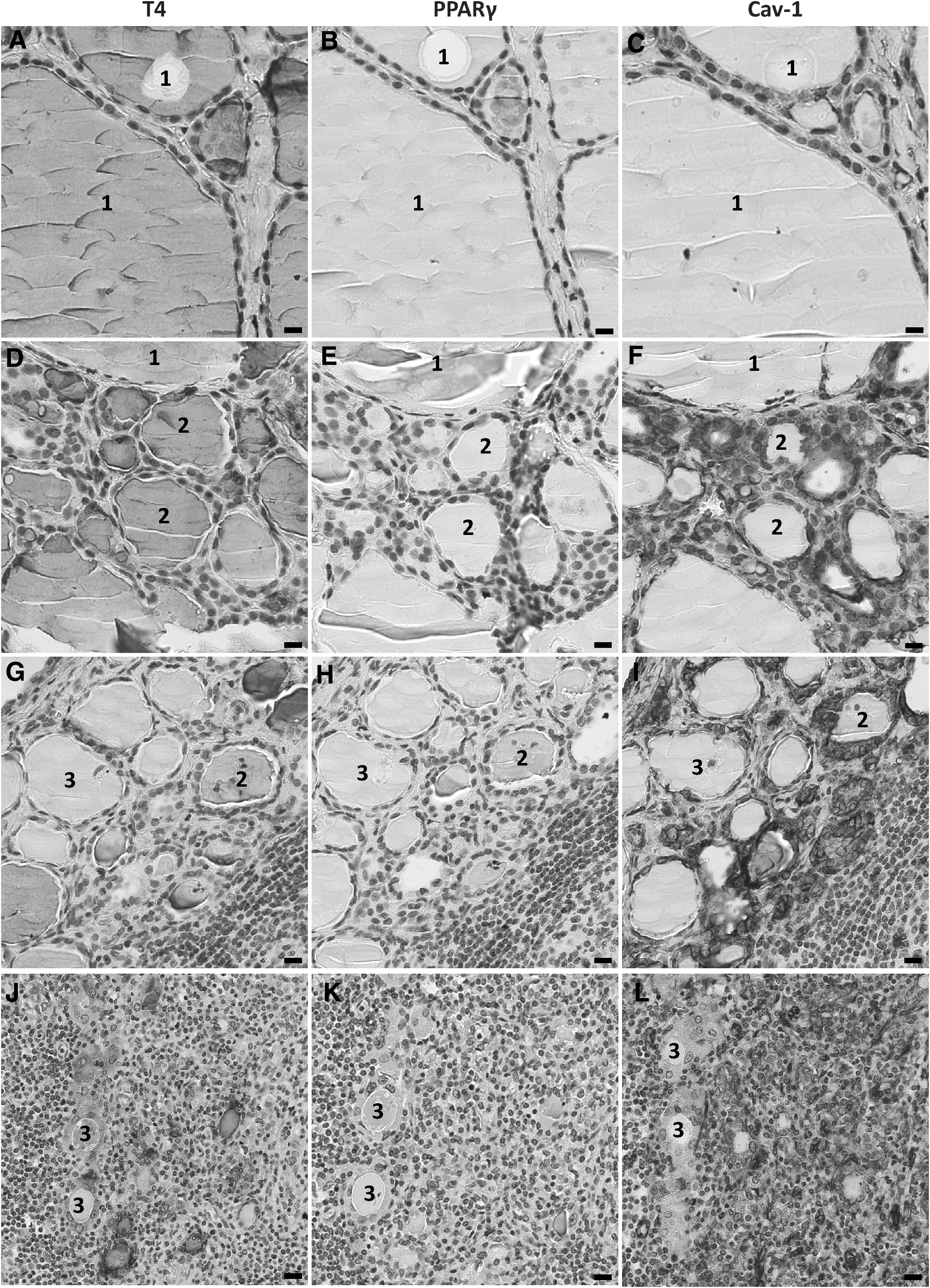
Immunohistochemical detection of T4, PPARγ, and Cav-1 in HT. In normal type 1 follicles (1), T4 is detectable in the entire colloid (
Type 2 follicles, which were smaller and had a higher epithelial layer, did actively synthesize T4, which was detected in the colloid (Fig. 2D). The expression of PPARγ was highly increased and present in nearly all of the cell nuclei (Fig. 2E). Cav-1 was also highly expressed in these follicles, as shown in serial sections (Fig. 2F).
Type 3 follicles were hypofunctional. They did not contain T4 in their lumina (Fig. 2G) and had lower expression of PPARγ (Fig. 2H) and Cav-1 (Fig. 2I) compared with type 2 follicles. The expression of PPARγ (Fig. 2K) and Cav-1 (Fig. 2L) was absent in type 3 follicles, showing T4 synthesis in the cytoplasm rather than in the colloid (Fig. 2J).
PPARγ and catalase mRNA expression is decreased by IL-1α and IFN-γ (Th1 cytokines) in primary cultures of human thyrocytes
A downregulatory effect of Th1 cytokines on Cav-1 expression in human primary thyroid cells has been recently published (17). Consistent with this observation, treating cells with IL-1α and IFN-γ, the predominant Th1 cytokines in HT, strongly decreased PPARγ mRNA expression (p < 0.01 compared with controls; Fig. 3A).

Effects of Th1 cytokines on PPARγ and catalase mRNA expression in primary cultures of human thyrocytes. (
Treatment with IL-1α and IFN-γ significantly decreased catalase mRNA expression (p < 0.05 compared with controls; Fig. 3B).
Pioglitazone, a PPARγ agonist, upregulates mRNA and protein expression of Cav-1 and TPO in primary cultures of human thyrocytes, and increases DUOX1 and DUOX2 mRNA expression, without modifying DUOX protein expression
Treatment of human thyrocytes in primary cultures with 15 μM of pioglitazone significantly increased PPARγ mRNA expression (p < 0.001 compared with controls; Fig. 4A). Similar results were obtained with 10 μM of pioglitazone (data not shown). Treatment of thyrocytes with pioglitazone also increased Cav-1 mRNA and protein expression (p < 0.05 compared with controls; Fig. 4B and C). The increase of Cav-1 protein expression was confirmed by quantification of immunofluorescence (Fig. 4D). In comparison with controls (Fig. 4E), Cav-1 expression was higher after treatment with pioglitazone (Fig. 4F). The co-staining of Cav-1 and PPARγ showed that these proteins were co-expressed and increased in the same cells after pioglitazone treatment (Fig. 4H) compared with controls (Fig. 4G). PPARγ protein expression was nuclear (Fig. 4H).

Effect of pioglitazone on PPARγ mRNA expression and Cav-1 mRNA and protein expression in primary cultures of human thyrocytes. (
mRNA (Fig. 5A) and protein (Fig. 5B) expression of TPO was significantly increased in human thyrocytes incubated with pioglitazone compared with controls (p < 0.01 and <0.05, respectively).
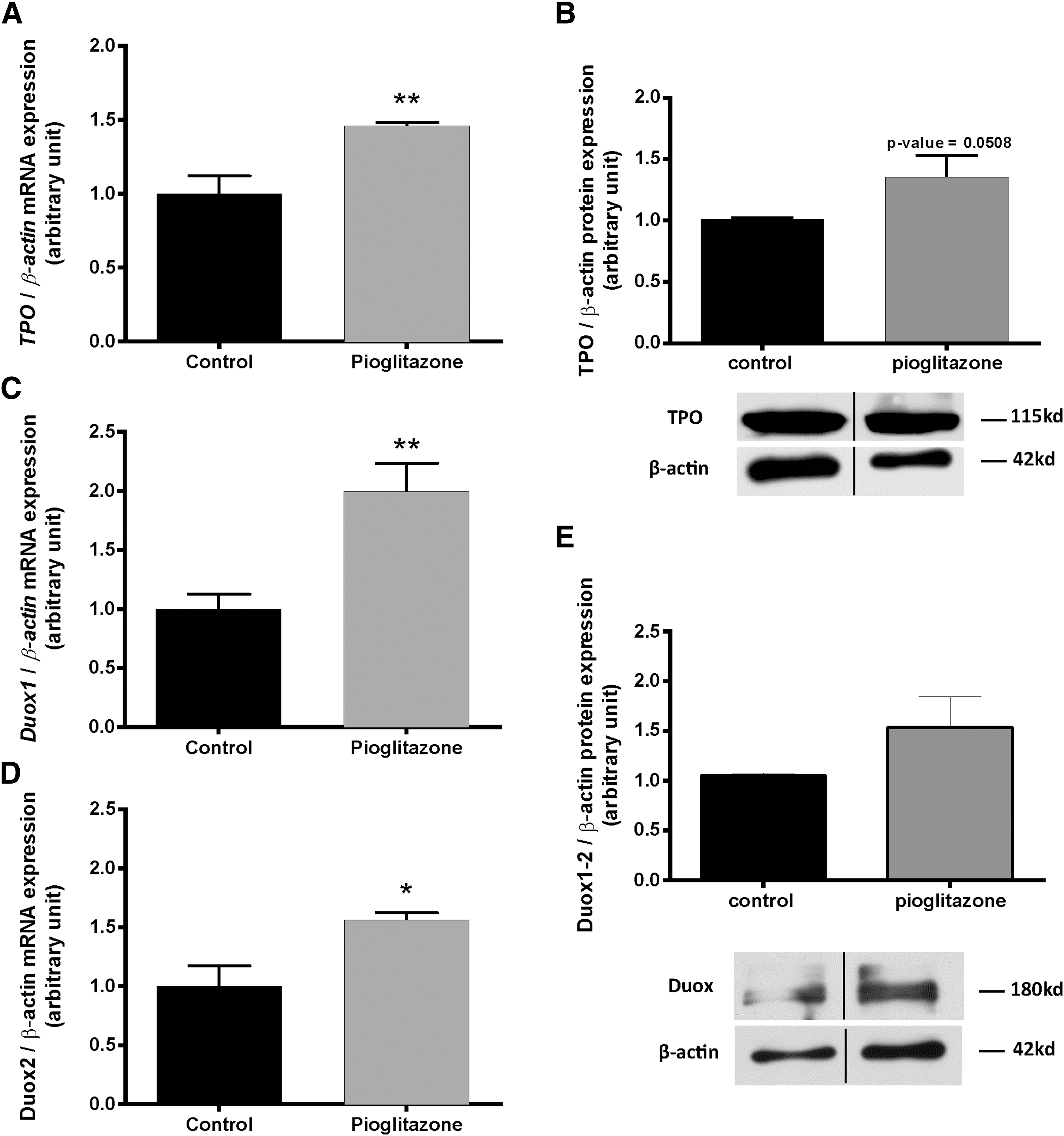
Effects of pioglitazone on TPO, DUOX 1–2 mRNA, and protein expression in primary cultures of human thyrocytes. The expression of TPO mRNA (
In addition, treatment with pioglitazone also significantly increased DUOX1 (Fig. 5C) as well as DUOX2 mRNA expression (Fig. 5D; p < 0.05 and <0.01 compared with controls, respectively). The total DUOX protein showed a non-significant tendency to increase upon PPARγ agonist treatment (Fig. 5E).
Pioglitazone, a PPARγ agonist, upregulates mRNA and protein expression of catalase in primary cultures of human thyrocytes
mRNA (Fig. 6A) and protein (Fig. 6B) expression of catalase was significantly increased in human primary thyrocytes incubated with pioglitazone compared with controls (p < 0.05).
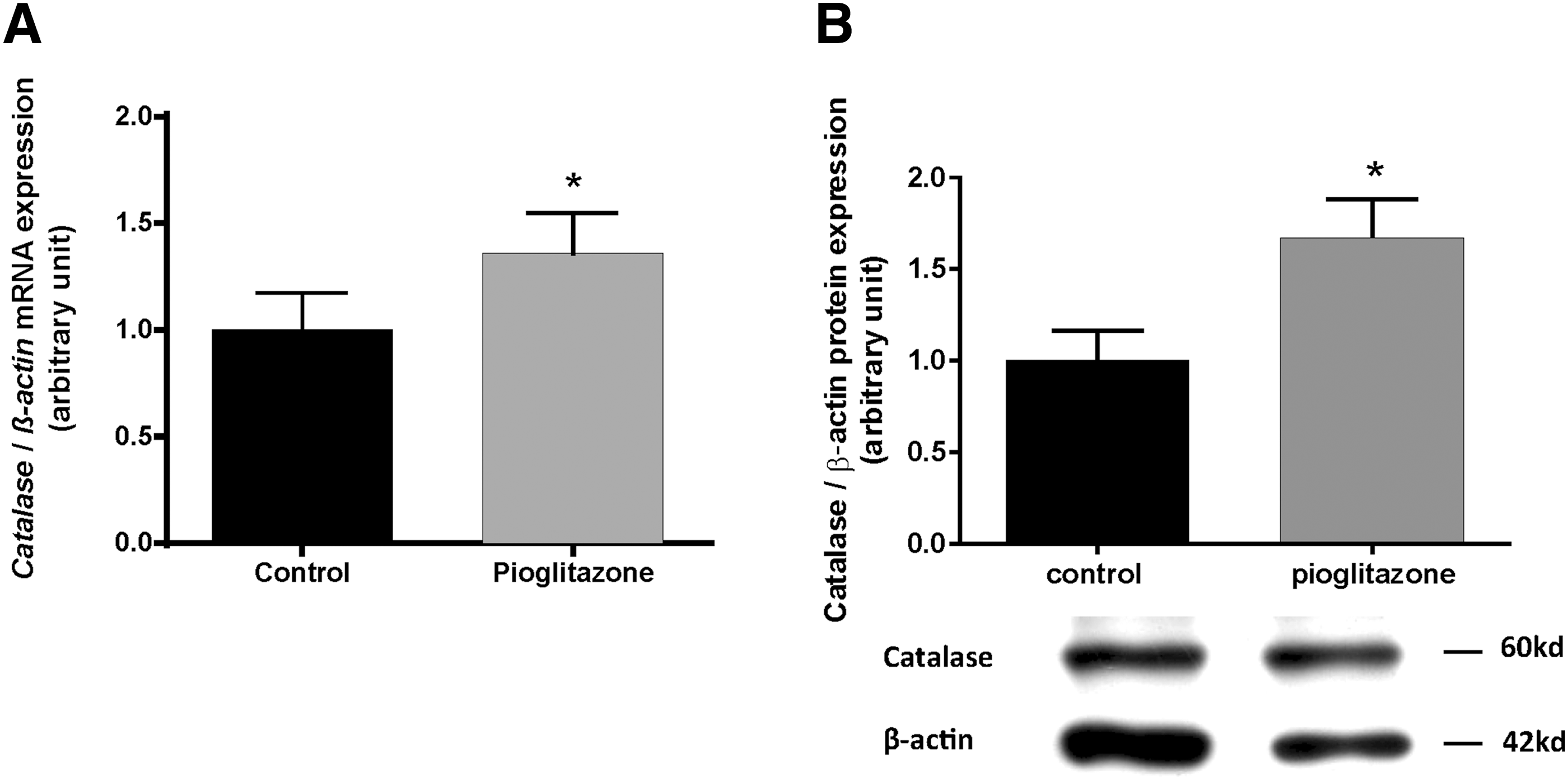
Effects of pioglitazone on catalase mRNA and protein expression in primary cultures of human thyrocytes. The expression of catalase mRNA (
Discussion
Pioglitazone and rosiglitazone—synthetic PPARγ ligands—have been shown to have a beneficial effect on the development of Th1-autoimmune thyroiditis by inhibiting the IFN-induced secretion of chemokines by thyrocytes (6,23). This study also demonstrate a beneficial effect of pioglitazone on Cav-1 expression and thus on the integrity of the thyroxisome, which is essential for thyroid hormone synthesis. It is indeed the first time that pioglitazone is shown to upregulate Cav-1 mRNA and protein expression in primary cultures of human thyroid cells, as already demonstrated in human macrophages (19) and mouse hepatocytes (20).
In the thyroid cell, Cav-1 has an important functional role. In Cav-1 knockout mice (14), the loss of Cav-1 leads to the mislocalization of TPO and DUOX, to intracellular iodination and intracellular H2O2 production associated to an oxidative stress, cell apoptosis, and finally hypothyroidism. The association between the loss of Cav-1 and the development of hypothyroidism has also been demonstrated in Pendred's syndrome (18) and in HT (17). The present study also shows that pioglitazone upregulates the mRNA and protein expression of Cav-1 and TPO, as well as DUOX mRNA expression. This further supports that Cav-1, TPO, and DUOX are important elements of the thyroxisome, which is key for maintaining thyroid cell homeostasis and euthyroidism. By increasing Cav-1 expression, pioglitazone may thus be facilitating normal thyroid hormone synthesis.
Disruption of the thyroxisome is a key event in the development of HT and is induced by Th1 cytokines (IFN-γ and IL-1α), which downregulate Cav-1, TPO, and DUOX (17). It has been previously observed that Th1 cytokines inhibit NIS mRNA expression (15,24) and thyroid hormone release (25). IL-1 and IFN-γ also downregulate TPO, DUOX, and Tg expression in human thyrocytes (16,26,27).
Moreover, this in vitro study demonstrates that Th1 cytokines downregulate PPARγ expression in human thyroid cells. A similar effect has already been observed in adipose tissue (7), but it is the first time that it has been reported in the thyroid gland. In HT, the suppressive effect of IFN-γ and IL-1α on PPARγ could be mediated by early growth response gene 1 (Egr-1), at least in part, as demonstrated in osteoarthritis chondrocytes (28). It could also be mediated by an excessive production of reactive oxygen species (ROS) by the inflammatory infiltrate surrounding the HT follicles and inside the thyrocytes. In addition to reactive nitrogen species (RNS), such as nitric oxide (NO) (29), ROS are also thought to act as intracellular mediators of Th1 cytokine-induced effects in thyroid cells (26).
There are multiple sources of ROS in thyroid cells, including free radicals, O2 ċ–, OHċ–, peroxyl (RO2 ċċ), alkoxyl (ROċ), and several non-radical molecules such as H2O2. H2O2, essential for thyroid hormone synthesis, is part of the physiological oxidative load. Although Th1 cytokines downregulate DUOX expression and therefore the production of extracellular H2O2 (16,17,26,27), the Th1-induced disruption of the thyroxisome leads to cytoplasmic localization of DUOX and excessive H2O2 production in the cytoplasm (17). The superoxide radical (O2ċ–) also plays an important role in the modulation of thyroid cell function, as shown by an increase of NOX2 mRNA and protein expression in response to the Th1 cytokines, in vitro in human thyroid cells and in vivo in HT (30).
To face the overproduction of RNS/ROS, the thyroid cells have strong antioxidant defenses of various types. Peroxiredoxins and catalase allow intracellular H2O2 to be detoxified: their expression is increased in HT, but is insufficient to compensate for the oxidative stress (17). The present data also show a decrease of catalase mRNA expression in the thyroid cells exposed to Th1 cytokines. Moreover, it is shown that pioglitazone increases catalase mRNA expression allowing the overproduction of intracellular H2O2 to be prevented and avoiding oxidative stress and cell apoptosis. The regulation of catalase by PPARγ has previously been demonstrated in human adipocytes (31), and it has also been shown that PPARγ activation is cardioprotective to H2O2-induced stress in the newborn heart by the upregulation of catalase (32). PPARγ ligands have a protective effect on renal ischemia/reperfusion injury by reducing iNOS (33). They also suppress NADPH oxidase (34) and restore superoxide dismutase in endothelial cells (35).
The extrapolation of these in vitro results, showing a downregulatory effect by Th1 cytokines on PPARγ expression, to the in vivo situation in HT is at first not obvious. Indeed, global quantification of PPARγ mRNA and protein expression showed no significant decrease in whole HT glands compared to controls. This could be explained by the great heterogeneity among Hashimoto's patients regarding inflammatory extent and by the fact that the inflammatory cells did express PPARγ (36). Moreover, within a single gland, the follicles are very heterogeneous from both a morphological and a functional point of view. This is already the case in normal thyroid tissue where active follicles and hypofunctionning follicles coexist (37). This follicular heterogeneity was even more accentuated in HT (17), and was very well demonstrated in the present study when considering PPARγ expression. Indeed, at the periphery of the inflammatory infiltrates, hyperactive type 2 follicles and inactive type 3 follicles were observed. In type 2 follicles, PPARγ expression was very high, and this could indicate a protective mechanism to face the Th1 cytokines produced by the inflammatory cells. In the same way, PPARγ could increase the expression of the thyroxisome components, mainly Cav-1, to maintain a normal apical iodination. PPARγ could also increase the antioxidant defenses of the follicular cells as shown by the strong catalase expression in the type 2 active follicles. At a higher degree of inflammation, Th1 cytokines completely impair the PPARγ expression as well as the one of Cav-1 and catalase, leading to a loss of normal hormonal synthesis. In some follicles, there are also signs of intracellular iodination, associated with the absence of PPARγ and Cav-1. The proportion of these inactive type 3 follicles will determine the level of hypothyroidism.
In conclusion, PPARγ is decreased by Th1 cytokines as shown by the present in vitro and in vivo results. This downregulation leads to a decreased expression of Cav-1, a key element of the thyroxisome, and alters the adequate targeting of TPO and DUOX at the apical membrane required to perform extracellular iodination. This opens up a new perspective in the physiopathology of HT, which might be due to a decrease of PPARγ by Th1 cytokines.
Activation of PPARγ by pioglitazone upregulates Cav-1 expression. This allows proper localization of TPO and DUOX at the apical membrane of the thyrocytes and efficient T4 formation in the colloid. Pioglitazone also upregulates catalase mRNA expression, allowing efficient protection against oxidative stress, which could be due to intracellular H2O2 accumulation in absence of Cav-1 (Fig. 7).

Schematic representation of the potential role of PPARγ in normal thyroid physiology and in the Th1-induced HT. PPARγ is thought to play a central role in normal thyroid physiology by upregulating Cav-1, essential for thyroxisome organization and extracellular iodination. By upregulating catalase expression, PPARγ also maintains cell homeostasis and prevents oxidative stress. The inhibitory effect of Th1 cytokines on PPARγ expression may be considered as a new pathogenic mechanism for HT. A downregulation of PPARγ leads to a disruption of the thyroxisome and a downregulation of catalase. The disruption of the thyroxisome is associated with intracellular iodination and an increased oxidative stress in the thyrocytes. If the thyrocytes cannot counteract the increased oxidative stress because of a decreased antioxidant defense, this can result in cell damage.
Footnotes
Acknowledgments
The authors wish to thank Marc de Bournonville for his technical help and Professors F. Miot and X. De Deken (IRIBHM, ULB, Brussels) for their collaboration and gifts of DUOX and TPO antibodies.
Author Disclosure Statement
The authors have nothing to disclose.