
Abstract
Background:
Understanding the roles of circulating microRNAs (miRNAs) can provide important and novel information regarding disease pathogenesis and a patient's clinical condition. Circulating miRNAs, such as exosomal miRNA, may regulate various bioactivities related to intercellular communication. However, the circulation of miRNAs in Graves' disease (GD) in relation to disease activity has never been elucidated. This study aimed to identify circulating miRNAs in GD in relation to disease activity and whether their exosomes play a role in the pathogenesis of GD.
Methods:
Circulating miRNAs were measured in serum obtained from seven intractable GD patients, seven GD patients in remission, and seven healthy controls using the miScript miRNA PCR Array. Altered miRNAs selected from array data were validated in 65 subjects. To investigate exosome biology, peripheral blood mononuclear cells (PBMCs) were incubated with exosomes isolated from the subjects' sera. mRNAs were quantified for cytokines using quantitative real-time polymerase chain reaction.
Results:
Circulating miR-23b-5p and miR-92a-39 were increased in GD patients in remission compared with intractable GD patients (p < 0.05). On the other hand, let-7g-3p and miR-339-5p were decreased in GD patients in remission compared with intractable GD patients (p < 0.05). Exosomes from intractable GD patients stimulated mRNA expression for IL-1β and TNF-α compared with GD patients in remission or healthy controls.
Conclusions:
This study demonstrates that different levels of circulating miRNAs are associated with intractable GD. Moreover, serum exosomes of patients with intractable GD may activate immune cells, which may play an important role in GD pathogenesis.
Introduction
G
miRNAs can be detected in serum and plasma as circulating RNAs. Interestingly, specific expression profiles of circulating miRNAs are associated with several diseases, including some cancers and autoimmune diseases (7 –11), suggesting that circulating miRNAs represent a new class of prognostic and diagnostic disease biomarkers. It was revealed that different levels of serum miRNAs were detected in patients with GD and Hashimoto's thyroiditis (HT), and they may play a role in the pathogenesis of these diseases (12). In GD, an important question is whether different levels of circulating miRNAs are associated with disease activity. However, circulating miRNA levels in GD in relation to disease activity (intractable GD and GD in remission) have never been analyzed.
Circulating miRNAs are secreted with exosomes, apoptotic bodies, and RNA binding molecules, such as Ago2 and HDL (13). Exosomes may act as a delivery system for cells, tissues, and organs; they may also regulate various bioactivities related to intercellular communication (14,15). Emerging evidence suggests that the biological activities of exosomes are partially mediated by exosomal RNA, which forms a portion of circulating RNA. Based on a previous report demonstrating the association between GD and circulating miRNAs (12), these miRNAs may be involved in disease pathogenesis and the clinical activity of GD. However, little is known about exosome function in the serum of GD patients.
This aims of this study were to identify circulating miRNAs in patients with GD, to establish whether there is a differential association with disease activity, and to assess whether exosomes play a role in GD pathogenesis. This is the first report characterizing circulating levels of miRNAs and exosome function in GD disease as a function of its activity.
Materials and Methods
Patients
All participants provided written, informed consent. The study protocol was approved by the Ethics Committee of the Fujita Health University School of Medicine. The diagnosis of GD was established by an endocrinologist. The diagnosis was made based on clinical symptoms, biochemical confirmation of hyperthyroidism, and positive antithyrotropin receptor antibodies (TRAb). Intractable GD patients had been treated with >10 mg of methimazole or 150 mg of propylthiouracil daily for at least two years, and the patients had persistent positive TRAb. GD patients in remission had been maintained in a euthyroid state without taking medication for more than three years; they were all negative for TRAb. Serum levels of thyrotropin (TSH), free triiodothyronine (fT3), and free thyroxine (fT4) were measured using a chemiluminescent enzyme immunoassay (Ortho-Clinical Diagnostics, Raritan, NJ). Serum TRAb titers were measured using an electrochemiluminescence immunoassay (ECLusys TRAb; Roche Diagnostics, Tokyo, Japan). Serum thyroperoxidase antibody (TPOAb) and antithyroglobulin antibody (TgAb) were analyzed using a radioimmunoassay (Cosmic Corporation, Tokyo, Japan). Graves' ophthalmopathy (GO) was defined as the presence of exophthalmos, blepharedema, diplopia, or an orbital sign, such as Graefe's or Moebius sign.
A total of 82 subjects who met the criteria were recruited in the discovery and validation set (Table 1). In the discovery study, 21 subjects (seven patients with intractable GD, seven patients with GD in remission, and seven healthy subjects) were recruited. In the validation study, 61 subjects who met the criteria were consecutively enrolled: 22 patients with intractable GD, 27 patients with GD in remission, and 12 healthy subjects. Serum samples for the measurements were stored at −80°C until assay.
Data are expressed as mean ± standard deviation.
Data are expressed as median and 25th–75th percentiles in parentheses.
p < 0.05 vs. healthy controls; † p < 0.05 vs. remission GD.
GD, Graves' disease; fT3, free triiodothyronine; fT4, free thyroxine; TSH, thyrotropin; TRAb, TSH receptor autoantibody; TgAb, antithyrogloblin antibody; TPOAb, thyroperoxidase antibody.
RNA isolation and microarray analysis of serum miRNAs
Serum obtained from intractable GD patients (n = 7), patients with GD in remission (n = 7), and healthy subjects (n = 7) was prepared. miRNA polymerase chain reaction (PCR) array analysis was performed to evaluate miRNA expression patterns in serum pools obtained from GD patients and healthy subjects using the miScript System according to the manufacturer's instructions (Qiagen, Valencia, CA).
Serum miRNA was isolated using a miRNAeasy Kit (Qiagen). Isolated miRNA was converted to cDNA using a miScript System (Qiagen). Prepared cDNAs were stored at −20°C until use. The miScript miRNA PCR Array for Serum and Plasma 384HC (Qiagen) containing 372 miRNAs was used for profiling the discovery set of 24 samples (Table 2). Real-time PCR (RT-PCR) results were normalized using the global mean of all miRNAs that had a cycle threshold (CT) value of <35, and miRNAeasy Serum/Plasma Spike-In Control (Syn-cel-miR-39 miScript miRNA Mimic) was used as the normalizing control. The CT values were selected based on the manufacturer's instructions from Qiagen because of a report that found that miRNAs with CT values ≥35 were associated with negative amplification (16). According to the manufacturer's instructions, Syn-cel-miR-39 miScript miRNA Mimic was spiked into the sample before nucleic acid isolation and detected with the control assay, as it provides a more useful normalization method for this application than the typical reference short noncoding RNAs, which are not normally detected in serum (16). Relative quantities of miRNA expression were calculated using healthy subjects as a control group. Expression analysis of miScript miRNA PCR Array results was performed using PCR Array Gene Expression Analysis Services (Qiagen).
miRNA, microRNA.
Quantitative RT-PCR of serum miRNAs
To validate the findings in the discovery set, several distinctive miRNAs were selected for the quantitative RT-PCR (qRT-PCR) validation studies based on the following criteria: (i) at least detectable levels in both GD patients and healthy subjects, (ii) at least twofold upregulation in intractable GD patients compared with GD patients in remission, and (iii) the presence of a report on the role of the miRNA in immune functions or the development of autoimmunity (17–33; Supplementary Table S1; Supplementary Data are available online at
Isolation of exosomes from serum
Exosomes were isolated from serum using a Total Exosome Isolation Reagent Kit (from serum; Life Sciences, Foster City, CA). All procedures were performed according to the manufacturers' protocols. Serum was obtained and stored at −80°C until use. Stored serum (500 μL) was processed by centrifugation at 3000 g for 10 min to remove cells and cellular fragments, and subsequent filtration of the supernatant was accomplished through Exosome Spin Columns (MW 3000). Total Exosome Isolation Reagent (Life Sciences) was added to the supernatants, and the exosomes were precipitated at 4°C for 1 h. Exosome pellets were collected by centrifugation at 10,000 g for 30 min and were dissolved in 50 μL of phosphate-buffered saline (10× concentrated exosome stock). The isolation of exosomes was validated by Western blot with a CD63 (exosomal marker) antibody (ab118307; Abcam, Cambridge, United Kingdom). In some studies, exosomal small RNA was extracted with a miRNAeasy Kit (Qiagen). The isolation of exosomal small RNA was confirmed by Agilent 2100 Bioanalyzer with Small RNA Analysis Kits (Agilent Techonologies, Santa Clara, CA).
Cell culture and exosome treatment
For these exosome studies in the samples from the validation set (Table 1), 22 samples were used (from eight patients with intractable GD, seven patients with GD in remission, and seven healthy subjects; Table 3). After extracting the exosomes from these serum samples, the exosome studies were performed. The efficacy of exosome extraction in all samples was confirmed using Western blot for CD63.
Data are expressed as mean ± standard deviation.
Data are expressed as median and 25th–75th percentiles in parentheses.
p < 0.05 vs. healthy controls; † p < 0.05 vs. remission GD.
GD, Graves' disease.
PBMCs were isolated with Lymphoprep (AXIS-Shiled, Oslo, Norway) as previously reported. PBMCs were cultured with RPMI 1640 medium with exosome-depleted fetal bovine serum (System Biosciences, Mountain View, CA). Twenty-four hours after isolation, PBMCs were incubated with diluted exosome stock solution (1 × concentrate) for 24 h. PBMCs were collected from female healthy donors.
qRT-PCR of mRNAs for cytokines
After incubating PBMCs with exosomes for 24 h, total RNA was obtained with TRIzol reagent according to the manufacturer's protocol (Invitrogen). cDNA was synthesized using M-MLV Reverse Transcriptase (Invitrogen) with a Random Primer (TaKaRa, Otsu, Japan). RT-PCR analysis was performed using the 7900 Real-Time PCR system (Applied Biosystems, Carlsbad, CA) with TUNDERBIRD SYBR qPCR Mix (Toyobo, Osaka, Japan). The sequences of primers used in this study are listed in Supplementary Table S2. Comparative threshold cycle values were used to determine relative mRNA expression levels. RT-PCR results were normalized by glyceraldehyde-3-phosphate dehydrogenase (GAPDH) expression levels.
Fluorescence microscopy analysis
Exosome stocks (10 × ) were incubated (at 37°C for 20 min) with SytoRNA Select Green Fluorescent (Invitrogen) that specifically binds RNA molecules. After incubation, exosomes were purified with Exosome Spin Columns (MW 3000; Thermo Fisher Scientific, Tokyo, Japan). Isolated human PBMCs were treated with 4′,6-diamidino-2-phenylindole (DAPI). Fluorescent exosomes were added to the medium, and cells were cultured for an additional 6 h. Subsequently, purified fluorescent exosomes and cells were detected with fluorescence microscopy (BZ-9000; KEYENCE, Osaka, Japan).
Statistical analysis
Data processing and analysis were conducted using tools from JMP v9.0 statistical software (SAS Institute, Cary, NC) and PCR Array Gene Expression Analysis Services (Qiagen). Comparisons of quantity and quality data were performed using the Mann–Whitney or Kruskal–Wallis tests, depending on data distribution. For multiple testing, the Bonferroni method was used to counteract the problem of multiple comparisons. Correlations between gene expression levels were analyzed by the Pearson product–moment correlation coefficient method. p-Values of <0.05 were considered statistically significant. For validation studies, the sample size was determined based on the results of the discovery study (intractable GD vs. GD in remission). Sample size and power calculations were conducted using tools from JMP v9.0 statistical software (SAS Institute).
A volcano plot was constructed for the detectable miRNAs. A volcano plot is a scatter plot that can be used to identify changes quickly in large data sets composed of replicate data. The x-axis shows the log2-fold change in the expression of the circulating miRNAs between cases and controls, whereas the y-axis shows the −log10 of the p-value for each miRNA, which represents the strength of the association. Therefore, miRNAs that are highly dysregulated are farther to the left and right sides, whereas highly significant changes appear higher on the plot.
Results
Differential circulating miRNA profiles of GD patients in relation to clinical activity
To determine whether there was a difference in the circulating levels of miRNAs in patients with GD, depending on disease activity, 21 sample sets were tested (intractable GD = 7, GD in remission = 7, healthy controls = 7) using miScript miRNA PCR Array for Serum and Plasma 384HC. As shown in Table 2, multiple differentially expressed circulating miRNAs were found with p-values of <0.05 in the comparisons of the different groups. Twenty-nine miRNAs distinguished intractable GD patients from healthy subjects, 35 miRNAs distinguished GD patients in remission from healthy subjects, and 14 miRNAs distinguished intractable GD patients from GD patients in remission (Table 2 and Supplementary Table S3). To assess the differences between the groups further, volcano plots were performed for each of the group comparisons. As shown in Figure 1, volcano plots demonstrated a distinction between intractable GD patients and GD patients in remission versus healthy subjects and between intractable GD patients versus GD patients in remission. These volcano plots provided a visual aid of potential differences between groups.
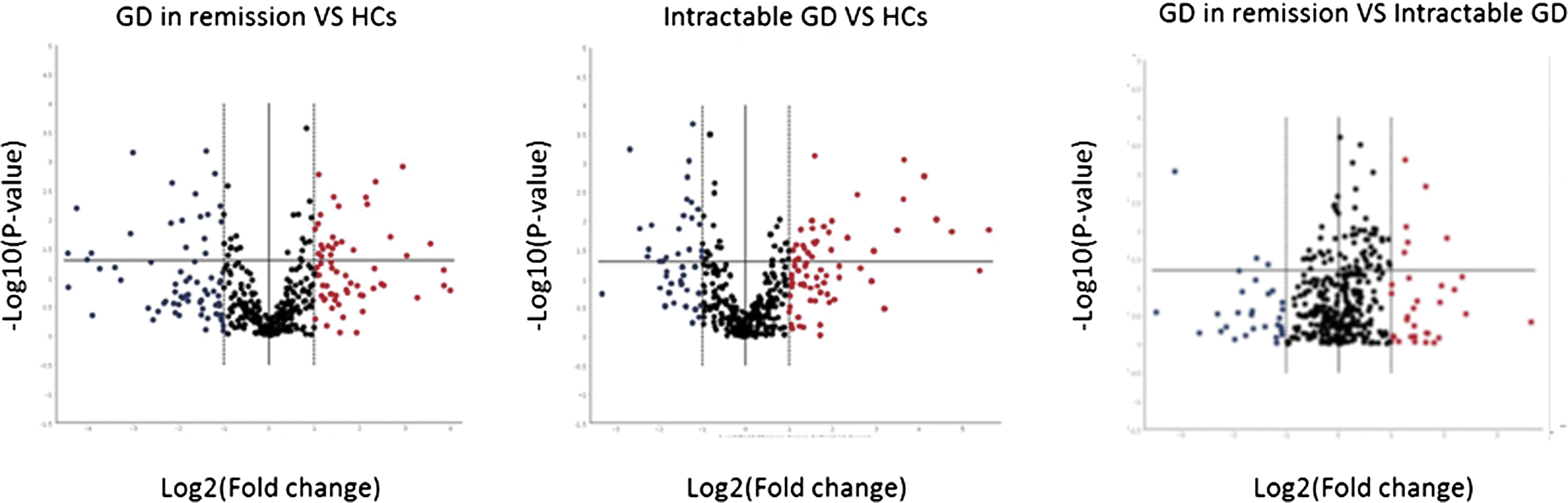
Volcano plots showing circulating microRNA (miRNA) profiles in patients with intractable Graves' disease (GD), GD in remission, and in healthy subjects. The volcano plots demonstrate a difference between intractable GD patients and GD patients in remission versus healthy subjects and between intractable GD patients versus GD patients in remission. Microarray data show significantly changed serum miRNA profiles. This plot displays both large-magnitude fold changes (x-axis) as well as high statistical significance (–log10 of p-value, y-axis).
To validate the findings obtained in the discovery set, several distinctive miRNAs were selected for qRT-PCR validation studies based on the following criteria: (i) at least detectable levels in GD patients and healthy subjects, (ii) at least twofold upregulation in intractable GD patients compared with GD patients in remission, and (iii) at least a report on a potential role in immune functions or the development of autoimmunity. Based on these criteria, six distinctive miRNAs were identified (Let-7g-3p, miR-23b-5p, miR-92a-3p, miR-92b-3p, miR-183-3p, and miR-339-5p). A set of 65 samples was tested (GD in remission = 27, healthy controls = 16, intractable GD = 22; Table 1). The serum levels of miR-23b-5p, miR-92a-3p, miR-92b-3p, and miR-183-3p were increased in GD patients in remission compared with patients with intractable GD (p < 0.05; Fig. 2). On the other hand, the serum levels of let-7g-3p and miR-339-5p were decreased in GD in remission compared with intractable GD (p < 0.05; Fig. 2). These results were consistent with the findings obtained in the discovery data set. Consequently, multiple testing demonstrated that miR-92b and miR-183 levels in GD in remission were not changed compared with intractable GD (p = 0.12 and p = 0.27, respectively). Furthermore, the levels of these miRNAs distinguished intractable GD from GD in remission and had no clear correlation with TRAb levels or having GO, respectively.

Circulating miRNAs in the validation study. Six distinctive circulating miRNAs (Let-7g-3p, miR-23b-5p, miR-92a-3p, miR-92b-3p, miR-183-3p, and miR-339-5p) measured in 61 samples (GD in remission = 27, intractable GD = 22, healthy controls = 12). The circulating miRNA levels were calibrated relative to healthy controls. The results are presented as the means from duplicate determinations. *p < 0.05.
Effect of serum exosomes on cytokine expression from PBMCs
It has been well established that exosomes contain various miRNAs, which are transferred into blood cells. Therefore, it was hypothesized that the physiological action of exosomes may differ between patients with GD in remission and intractable GD. First, the RNA transfer between exosomes and recipient cells (PBMCs) was examined. Exosomes were isolated from healthy individuals followed by purification with Exosome Spin Columns. It was found that purified exosomes express CD63 (exosomal marker) and contain small RNAs (Fig. 3A and B, respectively). After staining exosomal RNA with SYTO RNAselect Greeen fluorescent, purified fluorescent exosomes were added to the cell culture medium. As expected, the transfer of exosomal RNA into PBMCs was confirmed (Fig. 3C), suggesting the possibility that exosomal small RNA may exerts biological activities.
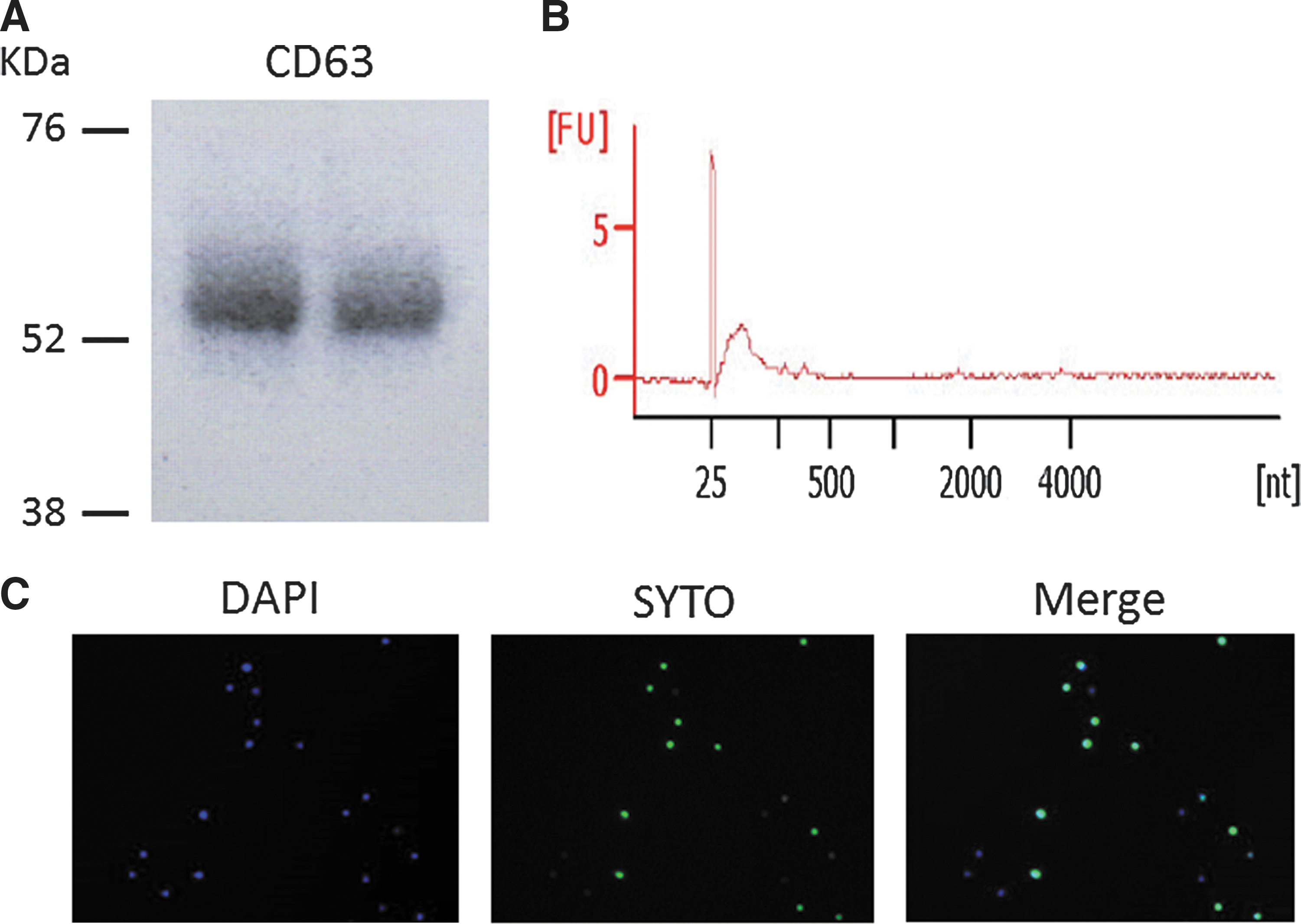
The transfer of miRNA between exosomes and peripheral blood mononuclear cells (PBMCs). (
Next, PBMCs were incubated with purified exosomes from healthy individuals, GD patients in remission, or patients with intractable GD for 24 h. Then, mRNAs for cytokines were quantified with RT-PCR (Fig. 4). No significant alteration of mRNA expression patterns was observed after incubating PBMCs with exosomes from GD patients in remission. However, exosomes from patients with intractable GD stimulated mRNA expression for IL-1β, TNF-α, and IL-6. Notably, particularly for IL-1β and TNF-α, drastic increments (4- and 10-fold, respectively) were observed after treatment with exosomes from patients with intractable GD (Fig. 4).
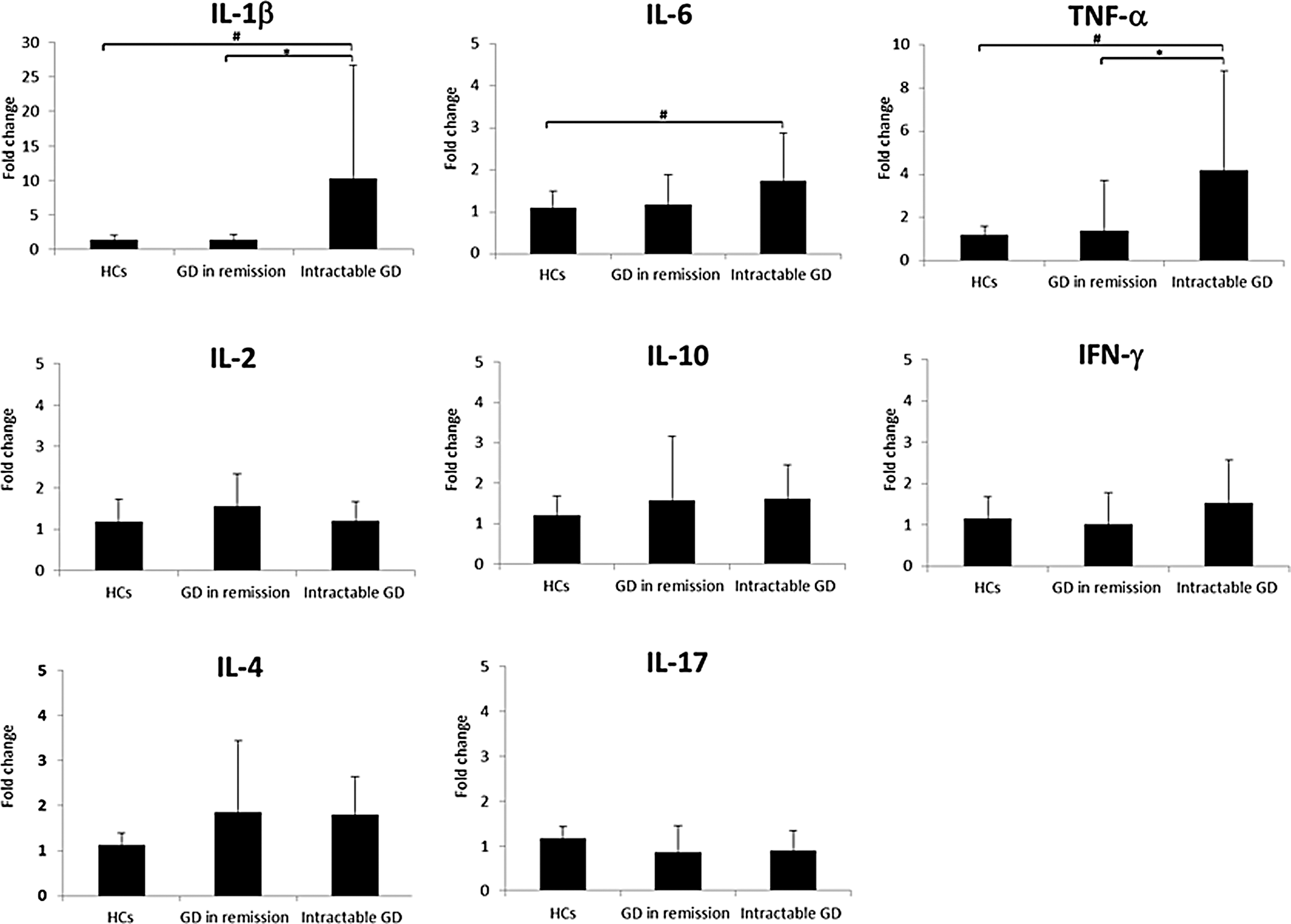
Effect of exosomes on cytokine expression in PBMCs. The expression of mRNA levels was calibrated relative to healthy controls. The results are presented as the means from duplicate determinations. *p < 0.05.
Discussion
This study demonstrates that different levels of circulating miRNAs are associated with the disease activity in GD patients. Moreover, the serum exosomes from patients with GD display bioactivity in terms of cross talk between immune cells, leading to the promotion of cytokine expression in PBMCs.
The expression patterns of several miRNAs in the serum from GD patients was evaluated using miRNA PCR Arrays (Fig. 1). There were 14 miRNAs that distinguished intractable GD from GD in remission (Table 2). To validate this finding, six representative immune-related miRNAs were selected and were validated by qRT-PCR. Compared with intractable GD patients, the serum levels of miR-23b-5p and miR-92a-3p were increased in GD patients in remission. In contrast, compared with intractable GD patients, the serum levels of let-7g-3p and miR-339-5p were decreased in GD patients in remission. Two reports confirmed that circulating miRNAs show differential profiles in autoimmune disease activity such as rheumatoid arthritis (RA) and multiple sclerosis (36,37). Some of the circulating miRNAs identified in this study were previously reported in autoimmune disease staging/activity other than GD. Gandhi et al. identified circulating miRNAs (let-7 and miR-92) that were differentially expressed in relapsing-remitting MS patients versus secondary progressive MS patients (37). A circulating miRNA signature (miR-23 and miR-223) may serve as a biomarker of response to anti-TNF-alpha therapy in RA patients (36). Consistent with these results, the present results show that different circulating miRNA profiles (miR-92a-3p and miR-23b-5p) were associated with disease activity in GD patients. Autoimmune disorders may have common proinflammatory regulatory mechanisms. Thus, circulating miRNAs may be accessible biomarkers for monitoring or predicting disease activity or therapy outcomes.
Recently, many miRNAs have been found to be important for the survival, development, differentiation, and function of many immune cell types (38,39). The results in Figure 2 and Table 2 indicate that several circulating miRNAs are associated with disease activity in GD. Altered miRNAs have emerged as crucial mediators involved in inflammation and autoimmunity processes. In GD remission, increasing miRNAs (miR-23b-5p and miR-92a-3p) can be central regulators of immune-suppression in the process of pro-inflamation. Let-7 and miR-23b regulate the nuclear factor-kappa B (NF-κB) signaling pathway, which is associated with inflammatory cytokine and chemokine production (17,40). Zhu et al. identified a potent anti-inflammatory role for miR-23b, which regulates the NF-κB signaling pathway, and they showed that this process is blocked by IL-17 through its repression of miR-23b expression during autoimmunity (41). miR-23b can limit tissue inflammation; its expression is repressed by IL-17 during autoimmunity. It is well known that Th17 cells develop and produce IL-17 during autoimmune processes. Th17 cells have been implicated in the pathogenesis and severity of autoimmune diseases, including AITD (18,34,37). miR-183 is known to be a key factor in TGF-beta1-mediated immune suppression (42). Because miR-183 silences immune cells, including tumor-associated natural killer (NK) cells by TGF-beta-mediated immune suppression, high circulating miR-183 levels could be a biomarker for the risk of tumor recurrence and poor survival in cancer patients. In GD, the TGF-beta1 gene is associated with the intractability or severity of GD (43). miR-92 belongs to the miR-17-92 cluster and has multi-functional targets (19). In an autoimmune mouse model, the miR-17-92 cluster has been shown to be involved in a lymphoproliferative disease and autoimmunity (44). miR-92a induces IL-6+ IL-10+ NKT cells; this fraction of NKT cells can suppress cytotoxic CD8+ T cells. Furthermore, the miR-17-92 cluster was specifically required for the accumulation of activated Ag-specific regulatory T cells (Tregs) and for differentiation into IL-10-producing effector Tregs (44). Inflammatory regulation or cytokine balance such as Th1/Th2 and Th17 cells play a key role in GD pathogenesis. Changes of several cytokines have been associated with disease activity in GD (45,46). This study supports that the cytokine balance is altered in intractable GD. Therefore, it is hypothesized that the differential levels of miRNAs after methimazole or propylthiouracil therapy may be associated with a reduction in inflammation and an improvement of the overall disease status of patients, leading to remission. Altered miRNAs may be a reflection of a response to therapy. Thus, further detailed analysis is required to estimate the long-term outcome data in a prospective longitudinal study in GD patients.
TRAb is generally used as a biomarker in predicting GD remissions. A previous study had demonstrated that TRAb can accurately predict short-term hyperthyroidism relapses after a course of antithyroid medications but was less effective in predicting long-term relapses or remissions (47). Thus, it will be worthwhile to identify new biomarkers to predict long-term relapses or remissions accurately. Interestingly, the level of circulating miRNAs distinguished intractable GD from GD in remission but had no clear correlation with the level of TRAb in this study. Therefore, these miRNAs might be associated with a different process than the antibody production of TRAb.
Recently, it has been recognized that miRNAs, as the cargo of exosomes in serum, have hormone-like effects, leading to widespread consequences on cells other than the secreting cells (13). The miRNAs cargo of exosomes can be specific to the type and activation status of their parental cells and play a role in the pathogenesis of various diseases (15,48). The role of exosomes and cross-talk between GD and immune cells has not yet been explored. Here, it was shown that the transcriptional activation of cytokines was induced by the exosomes of intractable GD patients. Several studies have shown an association between inflammatory cytokines and intractable GD (45,49). Song et al. reported that plasma inflammatory cytokine concentration in intractable GD patients was higher than in GD patients in remission (50). However, its mechanism has been largely unknown. The present study confirms the existence of a transfer of RNA between exosomes and PBMC (Fig. 3). Taking into consideration that four miRNAs (miR-92a-3p, miR-23b-5p, miR-339-5p, and let7g-3p) were present in the exosome (51), the upregulated cytokine production in intractable GD might be associated with the alternation of the exosomal miRNA profile. However, the exosomal miRNA profile was not identified.
It has been previously reported that exosomes regulate cytokine production. For example, body fluids such as amniotic fluid enhance cytokine secretion in THP-1 cells (52). On the other hand, the immunoreactivity of experimental autoimmune encephalomyelitis is suppressed by pregnancy exosomes (35). It is well known that communication between immune cells and cancer cells via exosomes play dual roles in modulating tumor immunity (53,54). Exosome-mediated immunosuppression occurs via the inhibition of antitumor responses elicited by immune cells, such as CD4+ and CD8+ T cells, and the induction of immunosuppressive cells, such as regulatory T cells. Moreover, Rodriguez-Munoz et al. demonstrated that circulating microvesicles regulate Tregs and Th17 differentiation in human autoimmune thyroid disorders (55). To the authors' knowledge, this is the first report to show that the biological activity of GD serum exosomes may vary with clinical disease activity. This phenomenon is potentially a novel clue for elucidating pathological mechanisms. Future studies should determine cell populations and other types of immune cells that are sensitive to exosomes derived from intractable GD.
Previously, differences were shown in circulating miRNA profiles between GD patients and healthy subjects (12). Enrichment was found in the circulating levels of miR-16, miR-22, miR-375, and miR-451 in GD patients, but these miRNA levels widely varied. This observation prompted the classification of GD according to disease activity and the investigation of circulating miRNA profile. Interestingly, it should be noted that upregulation of miR-22-3p has been observed in both remission and intractable GD consistent in a previous study. Although the physiological activity of miR-22-3p in GD has not been clarified, it is possible that this miRNA may play an important role in the pathogenesis of GD.
This is the first report to identify circulating miRNAs involved in GD activity. However, there are some limitations. Circulating miRNAs are associated with various types of diseases and may be potential biomarkers (56 –59). However, published studies, including the present study, lack long-term outcome data and an analysis in a large cohort. Circulating miRNAs may widely vary according to sex, race/ethnicity, or dietary intake, and may be influenced by other extrinsic factors, such as temperature (56). Moreover, smoking has been identified as a risk factor for GD, but it was not possible to assess the smoking history of the patients in this study. It will be necessary to confirm whether varied levels of circulating miRNAs are associated with smoking. Only Japanese GD patients and healthy subjects were recruited. Thus, it is unknown whether circulating miRNAs have racial/ethnic differences. The recruitment of other racial/ethnic groups may result in different circulating miRNA profiles. Furthermore, racial/ethnic differences in cytokine production have been previously reported (60). Taking into consideration that PBMC cytokine expression may be affected by the interaction between cells and exosomes, other racial/ethnic serum/exosome samples might provide a different cytokine profile from the ones observed in these Japanese patients. Further detailed investigations are required to establish the role of circulating miRNAs as useful biomarkers. According to the data on microRNA expression profiles in human tissues (20), the six circulating miRNA that were investigated (miR-92a-3p, miR-92b-3p, miR-23b-5p, miR-339-5p, let7g-3p, and miR-183-3p) are expressed throughout several tissues. Therefore, it is not possible to determine the tissue responsible for the alternating circulating miRNA levels.
In conclusion, this study demonstrates that different serum miRNAs levels are associated with disease activity in GD patients, and they may serve as novel biomarkers. Moreover, exosomes isolated from the serum of intractable GD patients may activate immune cells, which may play an important role in disease pathogenesis.
Footnotes
Acknowledgments
We thank Ms. Sayaka Nomura (Department of Endocrinology and Metabolism, Fujita Health University School of Medicine, Toyoake, Aichi) for her technical assistance.
Author Disclosure Statement
No competing financial interests exist.