
Abstract
Background:
3-Iodothyronamine (3-T1AM) is an endogenous decarboxylated thyroid hormone (TH) metabolite. Pharmacological doses of 3-T1AM decrease heart rate, body temperature, and metabolic rate in rodents—effects that are contrary to classic TH excess. Furthermore, a single dose of 3-T1AM was shown to suppress the hypothalamic–pituitary–thyroid (HPT) axis in rats. It was hypothesized that 3-T1AM might play a role in the fine-tuning of TH action and might have a direct regulatory effect on the thyroid gland.
Methods:
This study tested whether repeated 3-T1AM treatment interfered with thyroid function and the HPT axis in mice. Therefore, male C57BL/6 mice were intraperitoneally injected with 5 mg/kg of 3-T1AM or vehicle daily for seven days. Additionally, the effects of 3-T1AM on the differentiated rat thyrocyte cell line PCCL3 were analyzed.
Results:
Repeated administration of 3-T1AM decreased thyroidal mRNA content of the sodium iodide symporter (Nis), thyroglobulin, and pendrin in mice. No interference with the HPT axis was observed, as determined by unaltered pituitary mRNA levels of triiodothyronine-responsive genes, including thyrotropin subunit β. Furthermore, 3-T1AM treatment did not change transcript levels of hepatic triiodothyronine-responsive genes, such as deiodinase 1. In line with this, serum TH concentrations were not changed after the treatment period of seven days. In concordance with the in vivo findings, 3-T1AM decreased the thyrotropin-dependent expression of Nis and functional iodide uptake in PCCL3 cells in vitro. Additionally, uptake and metabolism of 3-T1AM by PCCL3 cells was observed, as well as 3-T1AM-dependent changes in intracellular Ca2+ concentration that might be involved in mediating the reported effects.
Conclusions:
In conclusion, 3-T1AM application decreased expression of selected TH synthesis genes by acting directly on the thyroid gland, and it might therefore affect TH synthesis without involvement of the HPT axis.
Introduction
T
During the last decades, a rediscovered endogenous decarboxylated TH metabolite called 3-iodothyronamine (3-T1AM) has raised attention, since it exerts profound pharmacological effects that are partially opposite to classic TH action in rodents (16 –18). A single intraperitoneal (i.p.) injection of 50 mg/kg of 3-T1AM reversibly decreased metabolic rate and rectal temperature in Djungarian hamsters and C57BL/6J mice, as well as the heart rate in C57BL/6J mice, within minutes (16,19,20). Furthermore, other manifestations of TH-dependent metabolic functions such as glucose and lipid utilization are influenced by 3-T1AM. Several studies have provided evidence for a lipolytic effect of 3-T1AM treatment in rodents (20 –22), while glucose metabolism seems to be inhibited or shifted toward gluconeogenesis (20,23). Interestingly, interference of a single dose of 3-T1AM with the HPT axis was observed in rodents. The i.p. injection of 50 mg/kg of 3-T1AM decreased plasma T4 and TSH within 120 minutes in rats (23), and the intracerebroventricular injection of 1.3 μg/kg reduced serum free T3 (fT3) in mice within 60 minutes (24). These data challenge the above-described classic feedback concept of the HPT axis. 3-T1AM is an endogenous TH metabolite, and can be produced from T4 by murine intestinal tissue (12). This gave rise to the hypothesis that apart from its activation to T3 by deiodination, an additional metabolic pathway for T4 generates 3-T1AM, a T3-antagonistic metabolite that provides an efficient rapid mechanism for sustaining homeostasis (16,25).
It was hypothesized that 3-T1AM plays a role in maintaining TH homeostasis by direct action on the thyroid gland. This study investigated whether repeated administration of 5 mg/kg of 3-T1AM influences thyroid function and the HPT axis in male mice. The in vivo data were complemented with in vitro experiments in the Fisher rat thyrocyte cell line PCCL3 (26,27), where in addition to 3-T1AM, effects of its potential metabolites thyronamine (T0AM) and 3-iodothyroacetic acid (3-TA1) were also analyzed. It was demonstrated that 3-T1AM exerts direct effects on thyrocytes in vivo and in vitro. In this study, the seven-day exposure to 5 mg/kg of 3-T1AM per day in mice, as well as 3-T1AM incubation of rat thyrocytes in vitro, suppressed expression of genes involved in TH synthesis without interference with the HPT axis.
Materials and Methods
Repeated 3-T1AM treatment of mice
Male C57BL/6J mice, aged three to four months old, were held at 21–22°C and a 12 h/12 h light/dark cycle in single cages with ad libitum access to water and food. All procedures were carried out in accordance with the European Community Council Directives (86/609/EEC) and with approval by Stockholm's Norra Djurförsöksetiska Nämnd.
Mice were treated with 5 mg/kg of 3-T1AM (dissolved in 60% dimethyl sulfoxide [DMSO] and 40% physiological saline, pH 7.4) by i.p. injection of 5 μL/g body weight once daily for seven days, as previously described (28). Control mice were sham injected with vehicle (60% DMSO and 40% saline) under the same conditions. Body weight, food and water intake were measured daily. The animals were sacrificed at day 8 (24 h after the last injection). Their organs were collected, snap frozen, and stored at −80°C until further processing.
RNA extraction from mouse tissue and quantitative real-time polymerase chain reaction
RNA was extracted from frozen tissues with the Aurum Total RNA Mini Kit (BioRad, Munich, Germany) according to the manufacturer's instructions, including a DNaseI digestion. A total of 250 ng of RNA from the thyroid or 500 ng of RNA from all other tissues were reverse transcribed to cDNA using the iScript cDNA Synthesis Kit (BioRad). Real-time quantitative polymerase chain reaction (qPCR) was carried out with the CFX Connect Real-Time PCR Detection System (BioRad) using ABsolute qPCR SYBR Green Fluorescein (Thermo Scientific, Waltham, MA). Specific primers were designed to be separated by at least one intron in the corresponding genomic DNA with the NCBI Primer-BLAST online tool (Supplementary Table S1; Supplementary Data are available online at
Measurement of TH concentrations in mouse sera
Total TH serum concentrations were measured with competitive radioimmunoassays (no. 4525 for total T3 and no. 4524 for total T4) from DRG Instruments GmbH (Marburg, Germany) according to the manufacturer's instructions, as described before (30). The coefficient of variation was ≤5% for both assays.
Culture of PCCL3 thyrocytes
Non-transformed rat thyroid PCCL3 cells (26,27) were grown at 37°C, 5% CO2, and a humidified atmosphere in Coon's F12 cell culture medium (Biochrom, Berlin, Germany) supplemented with 5% fetal bovine serum (FBS; Biochrom), 1 mIU/mL of bovine TSH, and five hormones (H5; i.e., 10 μg/mL of insulin, 5 μg/mL of transferrin, 10 nM of hydrocortisone, 20 ng/mL of glycyl-histidyl-lysine, and 10 ng/mL of somatostatin; Sigma–Aldrich, St. Louis, MO). Cells were grown without TSH and FBS to deplete them from TSH and TH(-metabolites) and to increase their responsiveness for the following stimulation in cell viability assays, qPCR analyses, Western blot, iodide uptake determination, and measurements of cellular energy metabolism. After seeding, the cells were grown for 24 hours in medium containing FBS, TSH, and H5. When the cells were fully attached to the cell culture dish, TSH depletion was initiated by using medium that only contained FBS and H5. Eighteen hours before the actual experiment, the cells were depleted from FBS by culturing them in medium that only contained H5. A prolonged depletion protocol was applied for a subset of experiments testing iodide uptake and Dio1 activity. In this case, TSH and FBS depletion were prolonged for 24 hours compared with the standard protocol.
Viability test of PCCL3 cells
TSH- and FBS-depleted PCCL3 cells in 96-well plates were stimulated with 0.1 mIU/mL or 0.5 mIU/mL of TSH ±1 μM of 3-T1AM, iodine-free thyronamine (T0AM), or 3-iodothyroacetic acid (3-TA1). Thyronamines and 3-TA1 were replenished after 24 h. After 48 h, 20 μL of a 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) stock solution (5 mg/mL) were added, and the cells were further incubated for 1.5 h at 37°C. Thereafter, cells were lysed in 12% HCl in isopropanol, and the optical density of the lysates containing the oxidized MTT dye was measured at 595 nM.
Measurements of TH metabolites by liquid chromatography–tandem mass spectrometry
PCCL3 cells were grown in 12-well plates for 24 h. After another 24 h under FBS depletion, cells were incubated ±1 μM of 3-T1AM in Krebs Ringer buffer (119 mM of NaCl, 4.74 mM of KCl, 1.19 mM of KH2PO4, 25 mM of NaHCO3, 2.54 mM of CaCl2, 1.19 mM of MgCl2, 10 mM of Hepes, and 11 mM of glucose; pH 7.4) for up to 180 min. For enzyme inhibition experiments, cells were pre-incubated with 100 μM of the monoamine oxidase inhibitor iproniazid, 10 μM of the Dio1 inhibitor 6n-propyl-2-thiouracil (PTU; Sigma–Aldrich), or solvent for 30 min before 3-T1AM was added to the medium. PTU was present during the whole duration of 3-T1AM incubation. Thyronamines and thyroacetic acids were concentrated from the conditioned cell culture supernatants by liquid/liquid extraction and measured by liquid chromatography–tandem mass spectrometry (LC-MS/MS) as previously described (31).
RNA extraction from cells and real-time qPCR
PCCL3 cells were grown in six-well plates and were TSH and FBS depleted as described above. Subsequently, cells were stimulated with 0.1 mIU/mL of TSH ± 3-T1AM for 3 h. RNA extraction, cDNA synthesis, and qPCR analysis were performed as described above. Hprt1 served as reference gene, and n-fold inductions were calculated with the 2(–ΔΔCt) method corrected for primer efficiency (E). Primer sequences are listed in Supplementary Table S2.
Western blot analysis
FBS- and TSH-depleted PCCL3 cells were stimulated with 0.5 mIU/mL of TSH ±1 μM of 3-T1AM for 48 h. 3-T1AM was refreshed after 24 h. At the end of the incubation time, cells were snap frozen and stored at −20°C until further processing. Cells were lysed using a homogenization buffer (20 mM of Hepes, 1 mM of EDTA, 250 mM of saccharose; pH 7.4). Homogenates were centrifuged at 13,000 g for 15 min at 4°C, and pellets containing the membrane fraction were resuspended in homogenization buffer. Protein concentration in the enriched membrane fraction was determined by the method of Bradford with Coomassie Brilliant blue G-250 dye (BioRad) (32). Twenty-five micrograms of protein were separated by SDS-PAGE and transferred to a nitrocellulose membrane (GE Healthcare, Little Chalfont, United Kingdom). Membranes were probed with antibodies against the sodium iodide symporter (NIS) diluted 1:10,000 (EUD410; Acris, Herford, Germany) and to β-Actin diluted 1:2000 (600-401-886; Rockland, Limerick, PA) overnight at 4°C followed by incubation with a horseradish peroxidase (HRP) conjugated secondary goat anti-rabbit antibody (P0448; Dako, Glostrup, Denmark). The specific immunosignals were visualized by chemiluminescence using ECL (GE Healthcare) as HRP substrate and films to detect the light reaction.
Nonradioactive measurement of iodide uptake
PCCL3 were depleted as described above and stimulated with 0.1 mIU/mL of TSH ± co-stimulation with 1 μM of 3-T1AM/T0AM/3-TA1 for 48 h. The co-stimulants were refreshed after 24 h. Cells were washed with 1 × phosphate-buffered saline and subsequently incubated with 10 μM of NaI (Sigma–Aldrich) for 20 min at 37°C/5% CO2. Intracellular iodide content was measured using the Sandell–Kolthoff reaction, as previously described (33,34), and corrected for nonspecific iodide as well as uptake of iodinated compounds by using 100 μM of perchlorate controls for every individual treatment condition.
Nonradioactive measurement of Dio1 activity
PCCL3 were depleted as described above and stimulated with 0.1 mIU/mL of TSH ±1 μM of 3-T1AM. Dio1 activity was measured with a nonradioactive iodide release assay, as previously described (35). In brief, PCCL3 cells were homogenized, and approximately 10 μg of protein was incubated with 10 μM of 3,3′,5′-triiodothyronine (rT3) (Sigma–Aldrich) for 2 h at 37°C. Afterwards, the iodide released from rT3 was measured with the Sandell–Kolthoff reaction. Specific Dio1 activity was calculated as pmol iodide/(min·mg protein) with the use of a NaI standard curve.
Measurement of cellular energy metabolism
A Seahorse XFe96 Extracellular Flux Analyzer (Seahorse Bioscience, North Billerica, MA) was used to measure glucose breakdown as extracellular acidification rate (ECAR) and mitochondrial respiration as oxygen consumption rate (OCR) in intact cells simultaneously (36). To this aim, PCCL3 cells were grown in Seahorse 96-well cell culture plates and were TSH- and FBS-depleted as described above. Before the Seahorse assay, cells were incubated for 60 min in a CO2-free incubator at 37°C with 1 mIU/mL of TSH ±1 μM of 3-T1AM in assay medium (D5030; Sigma–Aldrich), supplemented with 11 mM of glucose, 2 mM of L-glutamine, and H5. After 12 min of temperature equilibration in the Seahorse analyzer, measurements of OCR and ECAR were performed in cycles of 3 min mixing and 3 min measurement steps. The basal ECAR and OCR values were defined as the third measurement of each individual Seahorse assay (37). After completion of the Seahorse measurement, the protein content of every well was determined with the Micro BCA Protein Assay Kit (Thermo Scientific) according to the manufacturer's instruction. ECAR and OCR values of each well were normalized to the corresponding amount of protein.
Calcium imaging
PCCL3 cells were grown on glass cover slips for 24 h in medium with FBS, H5, and 1 mIU/mL of TSH. On the day of the experiment, cells were pre-incubated with 2 μM of fura-2/AM (Tocris Bioscience, Bristol, United Kingdom) for 15–45 min and subsequently treated with 3-T1AM ±20 μM of the TRPM8 blocker 4-(3-chloro-2-pyridinyl)-N-[4-(1,1-dimethylethyl)phenyl]-1-piperazine-carboxamide (BCTC; Tocris Bioscience) onstage on an inverted microscope, which was combined with a digital imaging system with UV excitation capability (TILL Photonics, Munich, Germany), as previously described (38). Fura-2 fluorescence was alternately excited at 340 and 380 nm. Emission was measured at 510 nm. The fluorescence ratio f340nm/f380nm served as a relative index of changes in cytosolic free calcium. Before the experiments, stable control baseline levels were obtained for 8–10 min. The control traces are designated with open circles in the figures.
Measurement of intracellular cAMP accumulation
PCCL3 cells were grown in 96-well plates. After TSH and FBS depletion, the cells were stimulated with 0.1–1 mIU/mL of TSH, 0.1–1 μM of 3-T1AM, or TSH +3-T1AM for 45 min in a HEPES-buffered solution that contained 1 mM of 3-isobutyl-1-methylxanthine (IBMX; Sigma–Aldrich) to inhibit phosphodiesterases (PDE). After cell lysis, the cAMP concentration was measured with a competitive immunoassay using the AlphaScreen technology (PerkinElmer Life Science, Boston, MA), as previously described (39).
Statistical analyses
GraphPad Prism v5 was used for all statistical analyses. Results are depicted as means ± standard error of the mean of the indicated number of mice or independent in vitro experiments (biological replicates). For comparison of two groups, an unpaired Student's t-test or a Mann–Whitney test was used. For comparison of more than two groups, one-way analysis of variance was applied, and if significant differences between groups were found, Dunnett's post-test was used.
Results
Body weight and food intake in mice are unchanged after seven days of repeated 3-T1AM treatment
To test whether 3-T1AM influenced thyroid function and the HPT axis, male C57BL/6 mice were treated for seven days with 5 mg/kg of 3-T1AM or vehicle i.p. daily. No significant differences between the treatment groups were observed for food and water intake or for body weight at any time during the study (Table 1).
Body weight of C57BL/6 mice measured 24 h after the last 3-T1AM (5 mg/kg) or vehicle injection and accumulated food and water intake over a treatment period of 7 days showed no differences between the treatment groups. Mann–Whitney test, n.s., p > 0.05, n = 8 per group.
3-T1AM, 3-iodothyronamine.
Repeated 3-T1AM treatment decreases expression of distinct TH synthesis genes without affecting the HPT axis in male mice
After repeated treatment with 3-T1AM, thyroidal mRNA concentrations of Nis, thyroglobulin (Tg), and pendrin (Pds) were significantly decreased in comparison with sham-injected mice (Fig. 1A). In contrast, the thyroidal expression of other genes involved in TH synthesis (TSH receptor [Tshr], thyroperoxidase [Tpo], dual oxidase 2 [Duox2], and Dio1) was not altered (Supplementary Fig. S1). In order to investigate if the described changes in mRNA expression were direct effects of 3-T1AM on the mouse thyroid, the study analyzed whether the HPT axis was affected in the treatment model. Serum total T3 and T4 concentrations were not different between treatment groups (Fig. 1B), and the expression of the hepatic T3 responsive genes Dio1 and Spot 14 was not altered by 3-T1AM treatment (Fig. 1C). In line with this, the evaluation of β TSH (Tshb), TRH receptor (Trhr), TRH degrading enzyme (Trhde), TH receptor β (Thrb), and Dio2 transcripts showed no differences between pituitaries of 3-T1AM- and sham-treated mice (Fig. 1C). This indicates a direct effect of 3-T1AM on the thyroid gland. However, it cannot be excluded that a compensatory adaptation of the HPT axis had already occurred after the treatment period of seven days.
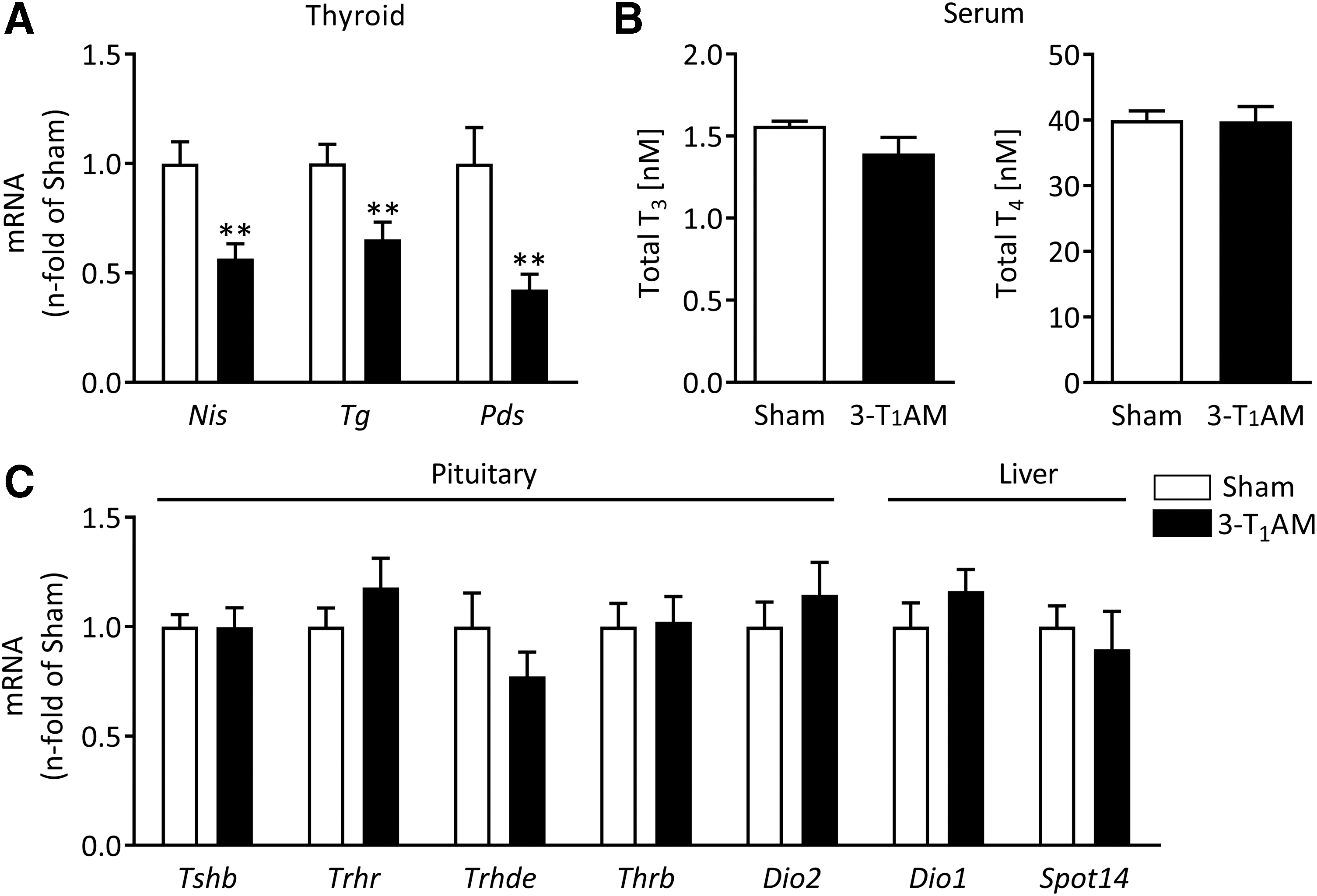
Effects of repeated 3-iodothyronamine (3-T1AM) treatment on the thyroid gland and the hypothalamic–pituitary–thyroid (HPT) axis. Thyroidal mRNA expression of sodium iodide symporter (Nis), thyroglobulin (Tg), and pendrin (Pds) was significantly decreased in 3-T1AM-treated mice compared with the control group, as determined by real-time quantitative polymerase chain reaction (qPCR; sham n = 8, 3-T1AM n = 7) (
3-T1AM exerts direct effects on thyrocytes in vitro
In order to analyze whether 3-T1AM could directly act on isolated thyrocytes, PCCL3 cells were chosen as an established in vitro model. This cell line is fully differentiated and dependent on TSH for growth (26,27).
3-T1AM, T0AM, and 3-TA1 do not affect the viability of PCCL3 cells
As expected, stimulation of PCCL3 cells with 0.1 mIU/mL and 0.5 mIU/mL of TSH alone led to an increase in viability compared with cells grown in the absence of TSH, as measured by a MTT test. In contrast, the test revealed no changes in viability of cells incubated for 48 h with TSH +1 μM of 3-T1AM/T0AM/3-TA1 compared to cells that were only incubated with TSH (Supplementary Fig. S2).
3-T1AM accumulates in PCCL3 cells and is enzymatically metabolized to T0AM and 3-TA1
3-T1AM is metabolized to T0AM and/or 3-TA1 by deiodination and oxidative deamination, respectively, in different in vivo and in vitro models (chemical structures are shown in Fig. 2A) (40 –42). To analyze the capacity of PCCL3 cells for 3-T1AM metabolism, first the expression of candidate enzymes for 3-T1AM metabolism was screened for using real-time qPCR. Figure 2B shows the specific bands representing qPCR products for Dio1 and monoamine oxidase A (Mao A), indicating that PCCL3 cells express two classes of enzymes that are known to be involved in 3-T1AM metabolism (40,43). No qPCR product from PCCL3 cDNA was obtained for Mao B. In contrast, real-time qPCR with the same Mao B primers in a rat liver cDNA positive control yielded a specific product of the expected size (data not shown).
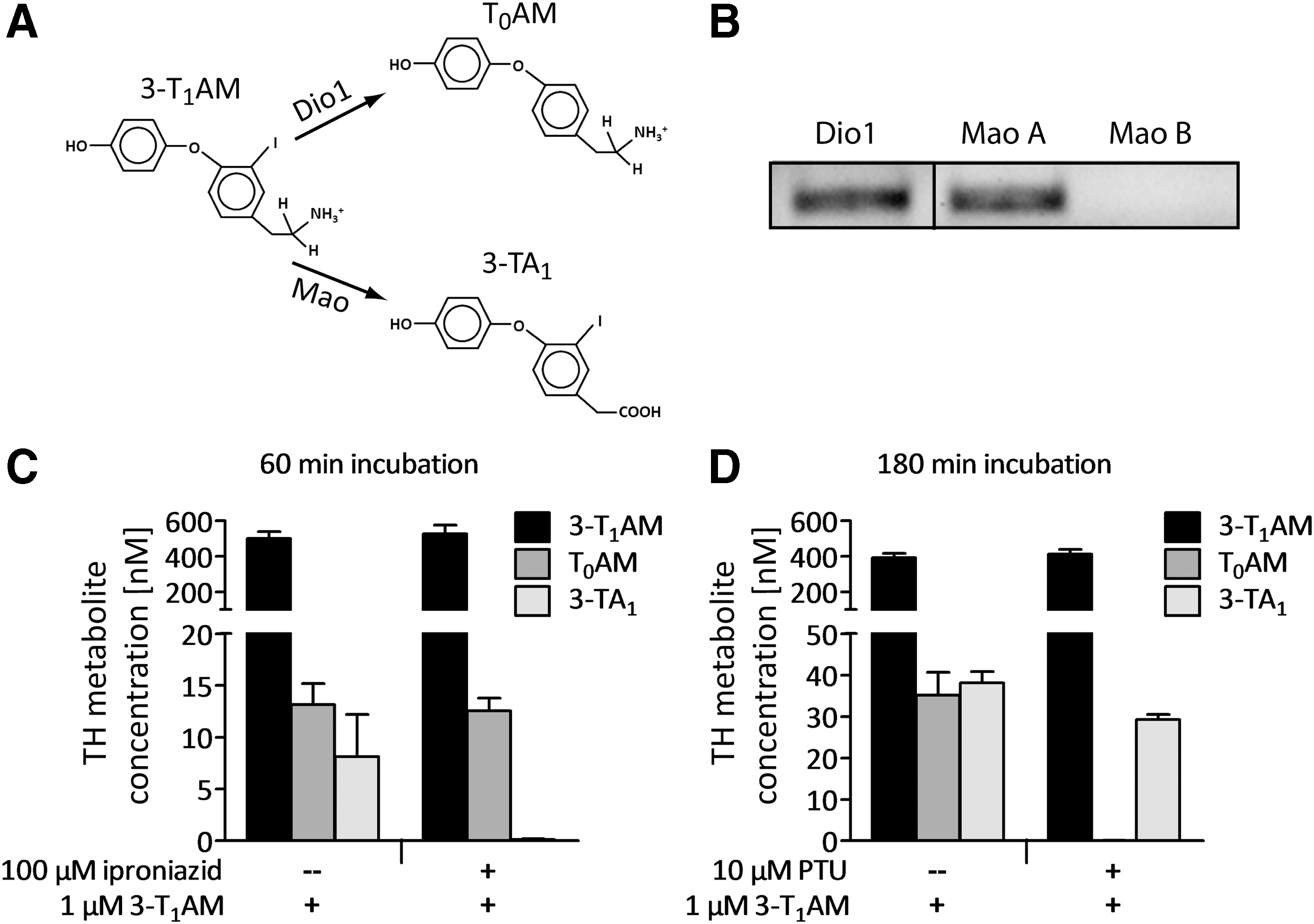
Uptake and metabolism of 3-T1AM by PCCL3 cells. Structures of 3-T1AM and its putative downstream metabolites, with arrows indicating the enzymes catalyzing the reactions (
Based on this transcript pattern of PCCL3 cells, the functional expression of Mao A and Dio1 was analyzed by incubating PCCL3 cells with the substrate 1 μM of 3-T1AM ± the Mao inhibitor iproniazid or PTU as specific Dio1 inhibitor. After 60 and 180 min of incubation with 3-T1AM, the expected products T0AM and 3-TA1 were detectable in culture supernatants (Fig. 2C and D). After pre-incubation with iproniazid, 3-TA1 was no longer detectable in the supernatants, indicating an involvement of Mao A in the formation of 3-TA1 from 3-T1AM (Fig. 2C). Likewise, if 3-T1AM incubation was performed in the presence of PTU, no T0AM was detected in the conditioned media, providing evidence for a contribution of Dio1 in the deiodination of 3-T1AM in PCCL3 cells (Fig. 2D).
3-T1AM decreases TSH-dependent NIS expression in PCCL3 cells
Co-incubation with 3-T1AM attenuated the TSH-dependent expression of Nis mRNA by approximately 50%, starting at a concentration of 100 nM (Fig. 3A). Other genes involved in TH synthesis, namely Tg, Tpo, Dio1, Pds, and Tshr, were not significantly altered in this experimental setting (data not shown). To assess whether the effect on the Nis gene was also manifested by protein translation, cells were stimulated for 48 h with 0.5 mIU/mL of TSH to allow for full NIS synthesis, post-translational modification, and integration into the plasma membrane (44). Co-incubation with 1 μM of 3-T1AM recapitulated the mRNA results and led to an approximately 40% reduction in TSH-dependent Nis protein expression (Fig. 3B and C).
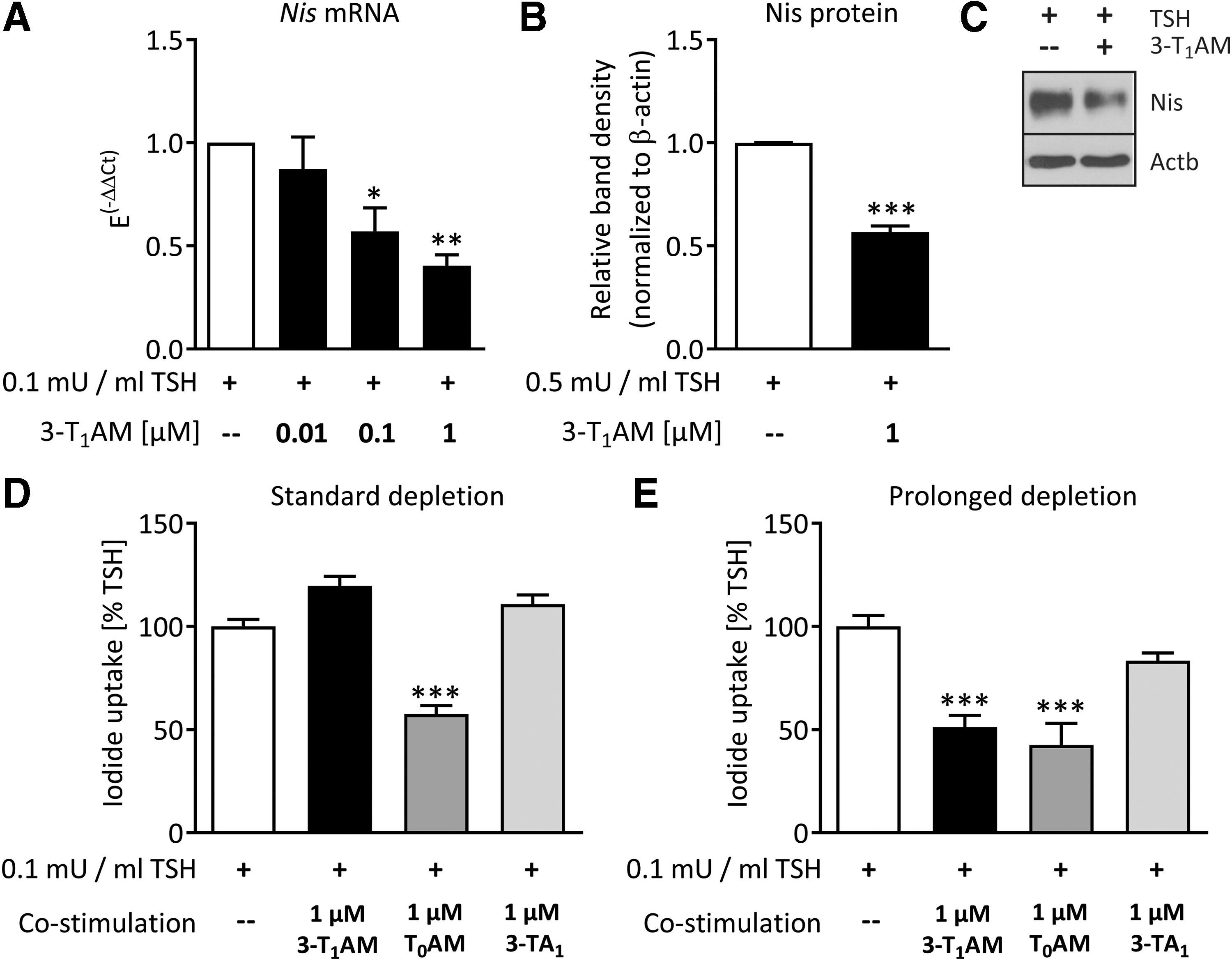
3-T1AM decreases thyrotropin (TSH)-dependent Nis expression and function in PCCL3 cells. 3-T1AM decreased the TSH-dependent Nis mRNA expression after 3 h of incubation, n = 3; *p < 0.05, **p < 0.01, one-way analysis of variance (ANOVA) followed by Dunnett's post-test (
3-T1AM decreases TSH-dependent NIS-mediated iodide uptake in PCCL3 cells
In order to analyze whether the inhibitory effect of 3-T1AM on TSH-dependent Nis mRNA and protein expression had a functional relevance, iodide uptake was measured (33). TSH incubation for 48 h substantially increased perchlorate-sensitive iodide uptake by PCCL3 cells (Supplementary Fig. S3). Using the same depletion protocol as for mRNA and protein analyses, no change in TSH-dependent iodide uptake by 3-T1AM co-incubation was observed (Fig. 3D). Therefore, a prolonged depletion protocol was applied. Cells were depleted from TSH for four days and grown for 42 h without FBS before the TSH ±3-T1AM co-incubation. Under these experimental conditions, 3-T1AM significantly decreased TSH-dependent iodide uptake in PCCL3 cells by 50% (Fig. 3E). Since PCCL3 cells metabolize 3-T1AM to T0AM and 3-TA1 (Fig. 2C and D), it was hypothesized that the different depletion protocols induced alterations in enzyme expression or activity profiles and therefore also in 3-T1AM metabolism. For this reason, PCCL3 cells grown under each depletion protocol were stimulated with TSH ±3-T1AM metabolites T0AM and 3-TA1 for 48 h. 3-TA1 co-incubation did not alter TSH-dependent iodide uptake under any depletion condition. In contrast, T0AM co-incubation led to a 50% decrease in TSH-induced iodide uptake under both depletion protocols. This indicates that in this context, T0AM may represent an active metabolite of 3-T1AM. Therefore, the question was raised whether the deiodinating metabolism of 3-T1AM and consequently the formation of T0AM were increased after prolonged compared with standard depletion. Determination of Dio1 activity revealed that Dio1 activity in TSH +3-T1AM-treated cells tended to be higher after the prolonged depletion than it was in equally stimulated cells after the standard depletion (Fig. 4).
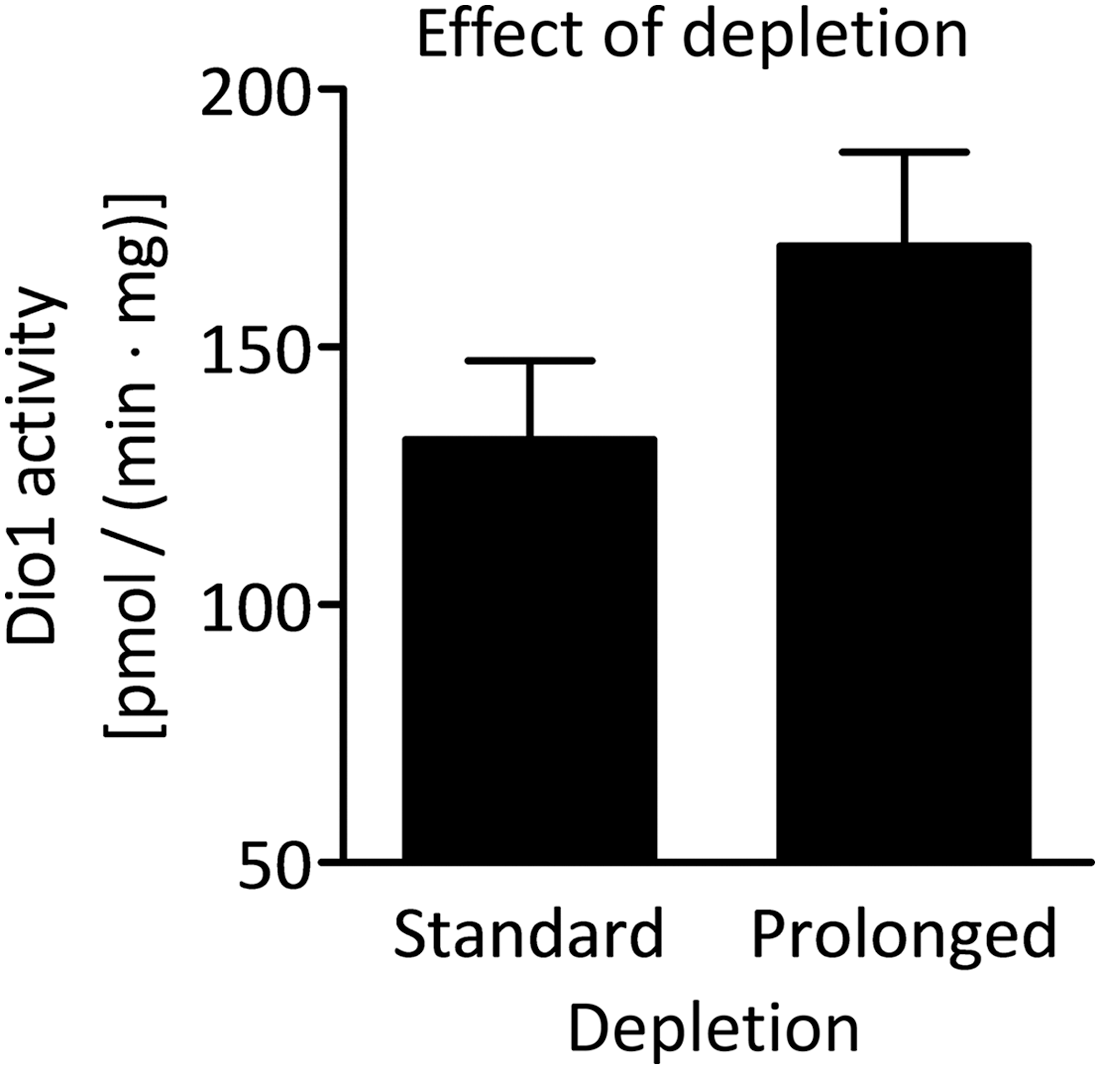
Effect of TSH and FBS depletion on Dio1 activity in PCCL3 cells. Dio1 activity in PCCL3 cells treated with 0.1 mIU/mL of TSH +1 μM of 3-T1AM after the standard and the prolonged depletion protocol, n = 3. No significant difference was observed between the two treatment groups (p > 0.05, unpaired t-test).
3-T1AM affects glucose utilization in PCCL3 cells after acute stimulation
Since 3-T1AM was previously shown to alter glucose utilization in animal models (20,23), the modulation of TSH-dependent glucose breakdown by 3-T1AM in PCCL3 cells was analyzed using the Seahorse XFe96 analyzer. Acute incubation of depleted cells with 1 mIU/mL of TSH significantly increased ECAR of PCCL3 cells (Fig. 5A). Co-stimulation with 1 μM of 3-T1AM decreased the TSH-dependent ECAR by approximately 30% (Fig. 5B). The oxygen consumption rate of the same cells, indicative for mitochondrial respiration, measured simultaneously was affected neither by TSH stimulation alone nor in co-stimulation with 3-T1AM under the described experimental conditions.
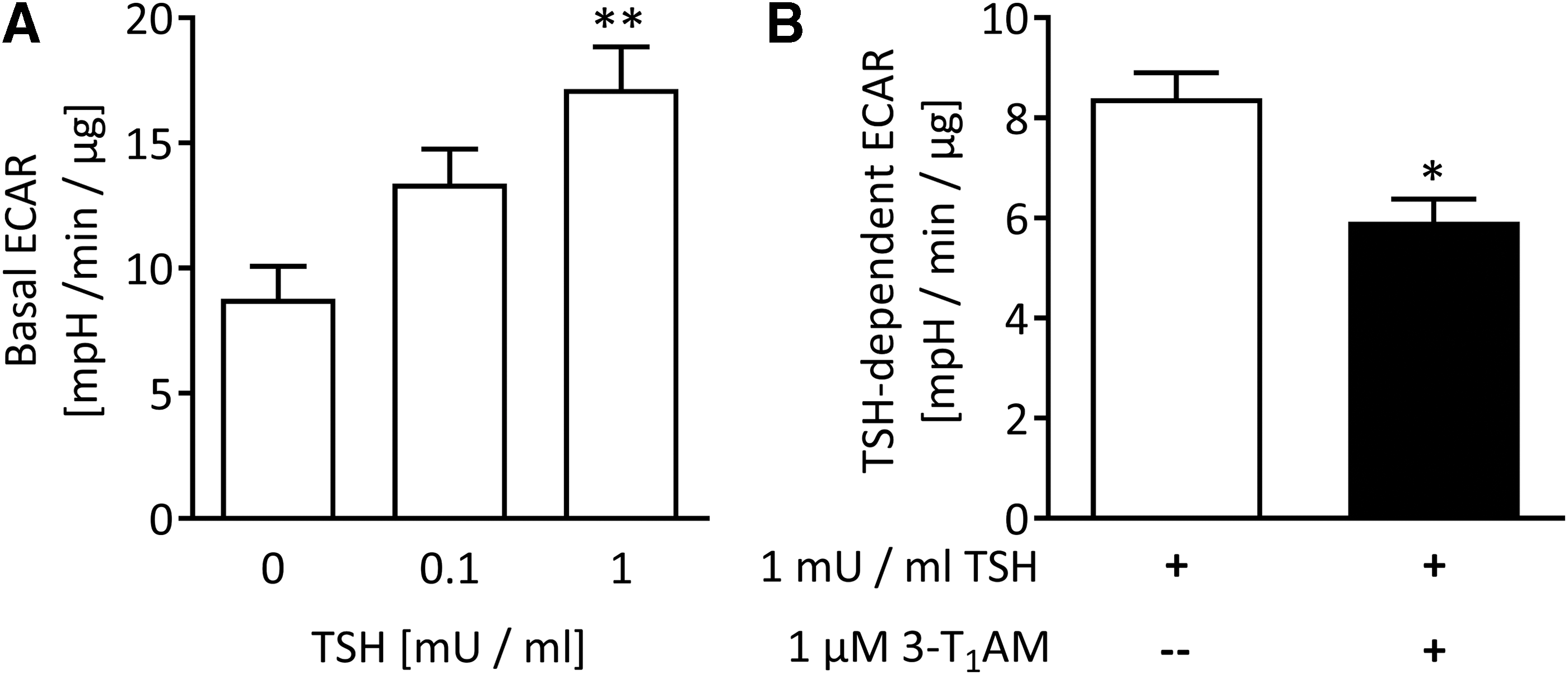
Basal ECAR measured in PCCL3 cells with the Seahorse Analyzer XFe96. TSH increased basal ECAR in PCCL3 cells within 60 min, n = 4; **p < 0.01, one-way ANOVA followed by Dunnett's post-test (
3-T1AM increases cytosolic free Ca2+ in PCCL3 cells
To elucidate the mechanism mediating the thyrocyte-specific effects of 3-T1AM, first both TSH-dependent signaling cascades were analyzed. Basal or TSH-dependent cAMP signaling was not modified by 3-T1AM in this experimental setting (Supplementary Fig. S4). No evidence for an effect of 3-T1AM on inositol 1,4,5-trisphosphate (IP3) formation was obtained. As 3-T1AM is a strong modulator of intracellular Ca2+ signaling via the transient receptor potential channel melastatin 8 (TRPM8, menthol receptor) in different cell types (38,45), the thyrocyte-specific calcium response to 3-T1AM was tested by using the calcium indicator fura-2 and fluorescence imaging. The application of 1 μM of 3-T1AM on fura-2-loaded PCCL3 cells led to a significant and quick increase in cytosolic free Ca2+ (Fig. 6A). Specifically, the f340nm/f380nm ratio increased from 1.201 ± 0.004 to 1.243 ± 0.005 after application of 1 μM of 3-T1AM (250 sec; p < 0.01; Fig. 6A and C), whereas the TRPM8 channel blocker BCTC (20 μM) clearly suppressed this effect (1.206 ± 0.005; p < 0.005; Fig. 6B and C). This indicates that the 3-T1AM-induced Ca2+ increase is due to Ca2+ channel activation such as TRPM8.
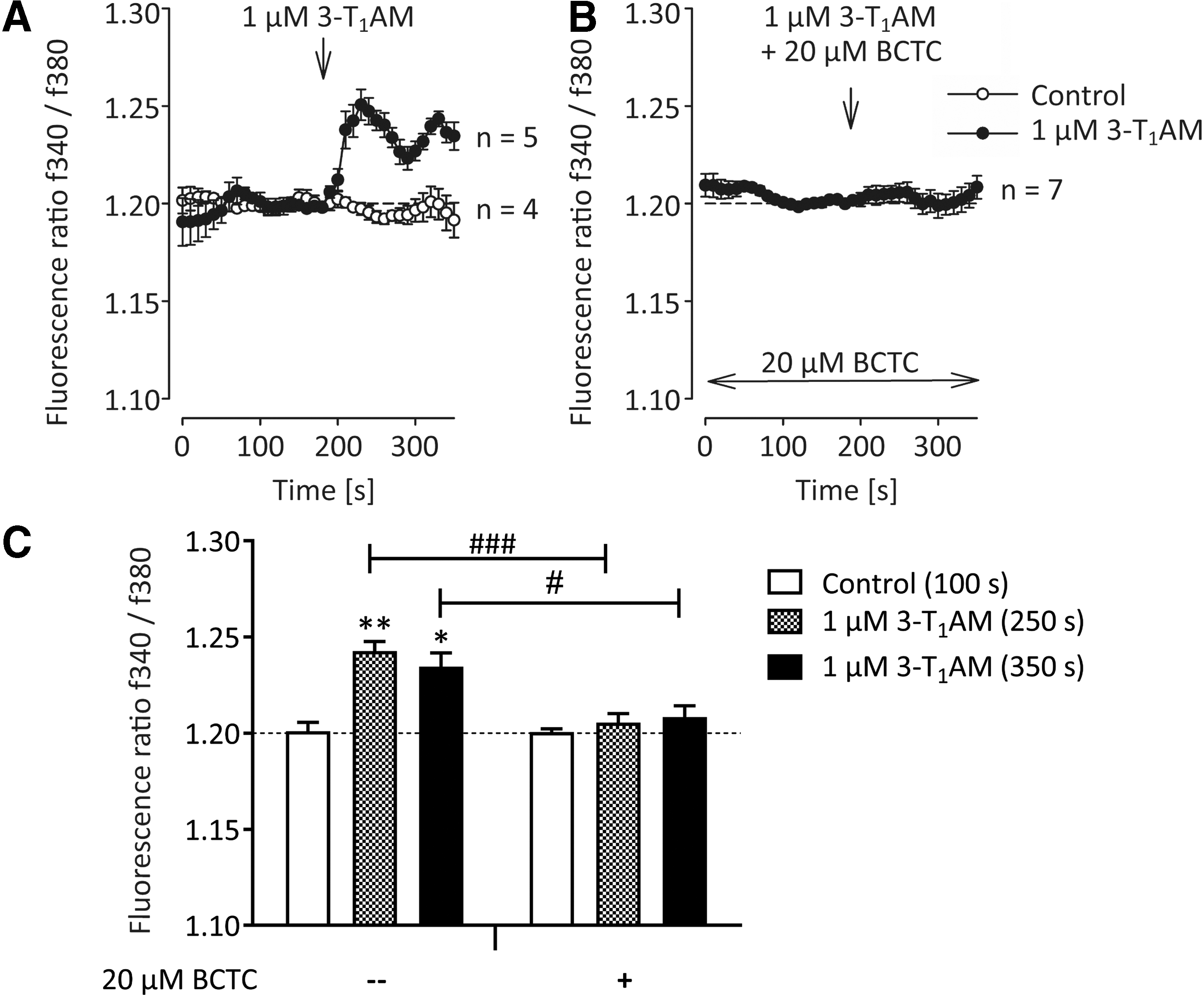
Effect of 3-T1AM on cytosolic free calcium in PCCL3 cells. The traces show intracellular Ca2+ measurements with 3-T1AM (n = 5; filled circles) and without 3-T1AM (n = 4; open circles), demonstrating a control Ca2+ baseline (
Discussion
3-T1AM is an endogenous decarboxylated TH metabolite that exerts effects partially antagonistic to TH and interferes with the HPT axis in rodents after single-dose application (16,18,23). In vivo evidence suggests extrathyroidal generation of 3-T1AM from T4 (12), an observation that does not exclude thyroidal contribution to circulating serum 3-T1AM (46). The recently observed formation of 3-T1AM from T4 supports the theory of alternate TH metabolism by peripheral tissues to ensure TH homeostasis and local fine-tuning of TH action. The aim of this study was to investigate whether repeated treatment with a moderate dose of 3-T1AM might interfere with thyroid function and the HPT axis in mice. Additionally, thyrocyte specific effects of 3-T1AM and its metabolites were analyzed in vitro using the PCCL3 cell line.
In vivo study
As the interference of a single dose of 50 mg/kg of 3-T1AM was shown to suppress the HPT axis in rats (23), the 10-fold lower dose of 5 mg/kg of 3-T1AM was chosen in the repeated treatment setup. The mice were injected with this amount of 3-T1AM or vehicle daily for seven days. Measurements with an in-house chemiluminescence immunoassay revealed that the 3-T1AM treatment procedure significantly increased 3-T1AM serum concentrations in the treated mice compared with the endogenous levels in the sham group (sham 17.4 ± 1.4 nM, n = 5; 3-T1AM 67.7 ± 7.3 nM, n = 6; p = 0.0043). This is only a fourfold increase. Therefore, the applied dose was considered physiological rather than pharmacological. While the expression of the Tshr and several other thyroidal transcripts were unaffected 24 hours after the last injection, mRNA expression of Nis, Tg, and Pds were downregulated by the repeated 3-T1AM treatment. NIS is the basolateral iodide symporter of thyrocytes, and accumulates iodide in the thyroid gland, which is the prerequisite for TH synthesis (47). The significance of iodide uptake into the thyroid gland is illustrated by the occurrence of thyroid disorders due to inadequate iodide supply (48). The TH synthesis and storage protein TG represents the major component of the follicular colloid. It provides tyrosine residues that are iodinated and form the TH precursors monoiodotyrosine (MIT) and diiodotyronine (DIT), and is therefore an important part of the TH synthesis system (49). PDS, the protein encoded by the Pendred syndrome gene, is an apical iodide transporter candidate in thyrocytes (50,51). Its importance is reflected by the goiter development in Pendred syndrome that is most likely caused by defective iodine organification due to reduced iodide efflux into the follicular lumen (52). Therefore, an inhibitory effect on the expression of Nis, Tg, and Pds as caused by 3-T1AM in this study may have the potential to interfere with TH homeostasis. The expression of T3-responsive genes in the pituitary that are important for the regulation of the HPT feedback axis (Tshb, Trhr, Trhde, Dio2, Thrb), as well as T3-responsive Dio1 and Thrsp (Spot 14) in the liver, were not altered by 3-T1AM treatment. These findings indicate that the HPT axis was not affected in the model, and provide evidence for a direct effect of 3-T1AM on the TH synthesis machinery in thyrocytes. Nevertheless, it cannot entirely be excluded that a compensatory adaptation of the HPT axis has already occurred after the treatment period of seven days. Despite the inhibited gene expression of Nis, Pds, and Tg, total TH serum concentrations were not altered after the seven-day treatment with 3-T1AM. A likely explanation might be that the duration of seven days of 3-T1AM treatment was not sufficient to deplete thyroidal Tg and TH stores (53,54).
In vitro studies
In order to comply with the 3R principle of animal ethics and before engaging in prolonged in vivo animal studies, advantage was taken of a more simplified in vitro thyrocyte model. To dissect the direct effects of 3-T1AM on thyrocytes in an isolated system at the molecular level, the PCCL3 thyrocyte cell line was chosen as an established in vitro model. These cells are highly differentiated and retain a non-transformed phenotype very close to in vivo thyrocytes (27). 3-T1AM is detectable in serum (18) and was shown to be bound to apolipoprotein B-100 (55). For this reason, PCCL3 cells were deprived from FBS before the 3-T1AM stimulation in order to start with a well-defined 3-T1AM concentration in the experimental setup. A decrease in the TSH-dependent mRNA and protein expression of NIS was observed after co-stimulation with 3-T1AM, which is partly in line with the in vivo data. This supports the concept of a direct inhibitory action of 3-T1AM on thyrocytes at doses that do not yet interfere with serum TH concentration or HPT feedback adaptation.
The functional TSH-dependent iodide uptake in PCCL3 cells was also decreased by 3-T1AM. Notably, this decrease only occurred after the cells had undergone a prolonged depletion protocol in which FBS and TSH depletion was prolonged for another day compared with the standard depletion protocol (3 days TSH free, 18 h FBS free). The in vitro setup revealed 3-T1AM metabolism to T0AM and 3-TA1 by Dio1 and Mao A, respectively. Both 3-T1AM-derived metabolites were shown to possess biological activity themselves in previous studies by others (16,23,56). In several single-dose application studies in rodents, T0AM was shown to exert similar but partially weaker effects than were exerted by 3-T1AM, for example in terms of cardiac output, body temperature, and plasma TH levels (16,23). No data on long-term effects of T0AM have been published yet. In contrast, 3-TA1 does not provoke cardiac or thermoregulatory changes in mice after acute or repeated treatment over seven days (57). The biological action of 3-TA1 known so far mainly comprises neurological effects via interaction with the histaminergic system (56,58). It was hypothesized that one of the 3-T1AM metabolites might actually contribute to the observed decrease in iodide uptake in PCCL3 cells. Furthermore, the different depletion protocols that were applied might have altered the extent of the 3-T1AM metabolism to T0AM or 3-TA1. While 3-TA1 was inactive regarding modulation of iodide uptake, a strong decrease in TSH-dependent iodide uptake by T0AM treatment was observed. This occurred regardless of the preceding depletion protocol, and indicates that T0AM (at least partially) mediates the observed effects of 3-T1AM incubation. This assumption is supported by the trend toward higher Dio1 activity in TSH ±3-T1AM-treated cells following the prolonged depletion compared with the standard protocol. The direct quantification of increased T0AM formation corresponding to the Dio1 activity can hardly be assessed in this experiment. This is probably due to further metabolism of T0AM as well as 3-TA1 to iodine-free thyroacetic acid TA0 (41). The source of TA0 cannot easily be determined yet by the currently available LC-MS/MS method.
To analyze whether 3-T1AM has effects on the energy metabolism of thyrocytes, mitochondrial respiration (OCR) and extracellular acidification (ECAR) were measured with the Seahorse Bioanalyzer. The TSH-dependent increase in glucose breakdown measured as ECAR was attenuated by acute 3-T1AM incubation. This is in line with the previously reported trend of high doses of 3-T1AM to decrease glucose utilization (20). It was shown that the pentose phosphate cycle plays an important role in the energy metabolism of thyrocytes and that it is TSH dependent (59). Further studies will be needed to clarify whether the measured ECAR is due to glycolysis or metabolism in the pentose phosphate pathway in the PCCL3 model. A reduced uptake of glucose is a possible explanation for the observed decline in ECAR by 3-T1AM stimulation. In the related FRTL5 thyrocyte model, treatment with 100 μM of 3-T1AM was shown to decrease glucose uptake within 60 min (41).
In order to elucidate the mechanism underlying the 3-T1AM-induced effects on thyrocytes, this study first tested whether 3-T1AM modifies signaling pathways downstream of the TSH receptor. TSH, the major regulator of thyroid function, is known to increase concentrations of the second messengers cAMP and IP3 in thyrocytes (60,61). So far, there is no evidence for an effect of 3-T1AM on IP3 formation in PCCL3 cells. Additionally, the TSH-dependent activation of the cAMP signaling pathway was not altered by 3-T1AM under conditions comparable to those resulting in functional and metabolic effects. Although it cannot be excluded that the site of action of 3-T1AM is further downstream from that of the TSH-mediated increase in the second messenger concentration, the present data support the concept that the 3-T1AM effects described above are independent of TSH signaling. Interestingly, a 3-T1AM-dependent BCTC-sensitive Ca2+ increase in PCCL3 was observed, which is in line with previous publications in ocular cells (38,45). This result suggests an involvement of Ca2+ channels such as TRPM8 in 3-T1AM-induced Ca2+ rise, since BCTC is known to be a TRPM8 blocker (62,63). Several lines of evidence could connect the increase in Ca2+ to the present results, and might be tested in future studies to explore Ca2+-induced signaling mechanisms in more detail. First, there is a subgroup of PDE that is activated by Ca2+ (64,65). PDE activation by 3-T1AM via Ca2+ could decrease intracellular cAMP. Of note, this would not be recorded by the cAMP assay in this study because it was performed in the presence of the PDE inhibitor IBMX. Second, the activation of protein kinase C, which is a downstream target of increasing Ca2+ concentration, is known to inhibit iodide metabolism in thyrocytes (66). Third, a complex cross-talk between calcium and cAMP signaling has been reported for thyrocytes (66,67). Therefore, the modulation of Ca2+ signals might be a promising hypothesis to be tested to explain the 3-T1AM effects in thyrocytes.
Taken together, the in vivo and in vitro data suggest that 3-T1AM can directly act on thyrocytes to inhibit iodide transport and the TH synthesis machinery without involvement of the HPT feedback axis. A non-classical Ca2+ signal-associated mechanism might contribute to the observed effects.
Footnotes
Acknowledgments
We thank Dr. Eva Wirth for helpful discussions and advice, Dr. Keith Richards and Dr. Daniel Rathmann for operating the LC-MS/MS, as well as Dr. Kostja Renko and Janine Martitz for sharing their expertise on Sandell-Kolthoff based assays. The technical assistance of Kristin Fischer, Ray Monk, and Anja Fischbach is highly appreciated. This work was funded by the Deutsche Forschungsgemeinschaft DFG (KO922-16/1-2# to J.K. and S.M., ME1706/14-1# to S.M., BI893/5-2# and BI893/6-3 to H.B., HO5096/1-1 and HO5096/2-1# to C.S.H., and MI1242/2-1# to J.M.), the Charité (J.K.), and the Swedish Research Council (J.M.) (#funded by the DFG funded Priority Programme SPP1629 THYROID TRANS ACT).
Author Disclosure Statement
No competing financial interests exist.