
Abstract
Background:
Thyroglobulin (Tg) stored in thyroid follicles regulates follicular function in thyroid hormone (TH) synthesis by suppressing thyroid-specific gene expression in a concentration-dependent manner. Thus, Tg is an intrinsic negative-feedback regulator that can restrain the effect of thyrotropin (TSH) in the follicle. However, the underlying mechanisms by which Tg exerts its prominent autoregulatory effect following recognition by thyrocytes remains unclear.
Methods:
In order to identify potential proteins that recognize and interact with Tg, mass spectrometry was used to analyze immunoprecipitated Tg-bound proteins derived from Tg-treated rat thyroid FRTL-5 cells.
Results:
Flotillin 1 and flotillin 2, two homologs that are integral membrane proteins in lipid rafts, were identified as novel Tg-binding proteins with high confidence. Further studies revealed that flotillins physically interact with endocytosed Tg, and together these proteins redistribute from the cell membrane to cytoplasmic vesicles. Treatment with the lipid raft disrupter methyl-β-cyclodextrin abolished both the endocytosis and the negative-feedback effect of Tg on thyroid-specific gene expression. Meanwhile, siRNA-mediated knockdown of flotillin 1 or flotillin 2 also significantly inhibited Tg effects on gene expression.
Conclusion:
Together these results indicate that flotillin-containing lipid rafts are essential for follicular Tg to be recognized by thyrocytes and exert its negative-feedback effects in the thyroid.
Introduction
T
Thus, the function of thyrocytes in TH synthesis is likely delicately balanced between two opposing actions: stimulation by circulating TSH from the basal side, and negative-feedback regulation by follicular Tg from the apical side (follicular lumen). Although TSH signaling pathways are well established, the underlying molecular mechanism by which Tg exerts its negative-feedback effects remains largely unclear. To understand these mechanisms better, characterization of the interaction and specific recognition of Tg by thyrocytes is needed. Thus, this study aimed to identify proteins that can recognize and interact with Tg to mediate its negative-feedback effects in thyrocytes.
Materials and Methods
Cell culture and treatment
Rat thyroid FRTL-5 cells were grown in Coon's modified Ham's F-12 medium supplemented with 5% bovine serum (Invitrogen, Waltham, MA) and a six-hormone mixture (1 mIU/mL of TSH, 10 mg/mL of insulin, 10 ng/mL of somatostatin, 0.36 ng/mL of hydrocortisone, 5 mg/mL transferrin, and 2 ng/mL of glycyl-L-histidyl-L-lysine acetate) as previously described (8). Bovine Tg (Sigma-Aldrich, St. Louis, MO) that was previously confirmed to have essentially the same effect as highly purified 19S Tg (4,11,12) was dissolved in the culture medium and used at a final concentration of 5 mg/mL, which is within the lower range of Tg concentrations in the thyroid follicles (3,13). FRTL-5 cells were incubated with bovine Tg in a humidified incubator at 37°C with 5% CO2. To disrupt lipid rafts, cells were incubated with 5 mM of methyl-β-cyclodextrin (MβCD; Sigma-Aldrich) for 2 h prior to Tg treatment.
Protein preparation and immunoprecipitation
Cells incubated with bovine Tg for 48 h were lysed in a lysis buffer containing 50 mM of HEPES, 150 mM of NaCl, 5 mM of EDTA, 0.1% NP40, 20% glycerol, and a cOmplete Mini protease inhibitor cocktail tablet (Roche Diagnostics, Basel, Switzerland) for 1 h. The lysates were centrifuged at 4°C for 20 min to recover cell proteins. Protein concentration was determined using DC protein assay reagents (BIO-RAD, Hercules, CA) and a VMax Kinetic Microplate Reader (Molecular Devices, Sunnyvale, CA) according to the manufacturers' instructions. Tg-bound proteins were co-immunoprecipitated with a goat anti-Tg antibody (sc-7836; Santa Cruz Biotechnology, Dallas, TX) in the presence of Protein A Sepharose beads (GE Healthcare Life Sciences, Little Chalfont, United Kingdom). Briefly, protein extracts were incubated with Protein A Sepharose beads at 4°C for 2 h with gentle rotation and then centrifuged to yield supernatants with pre-cleared proteins. The pre-cleared proteins were incubated with the goat anti-Tg antibody at 4°C overnight with rotation, followed by another 2 h of incubation with Protein A Sepharose beads. The beads were pelleted by centrifugation, washed sequentially eight times with lysis buffer and three times with phosphate buffered saline (PBS), and Tg-bound proteins were recovered after centrifugation.
Mass spectrometry analysis
Immunoprecipitated Tg-bound proteins were digested with trypsin overnight at 37°C. The tryptic digests were desalted using C18 membrane filters, and then analyzed on a TripleTOF 5600 system (AB SCIEX, Foster City, CA) coupled to a DiNa-AP (KYA Technologies, Tokyo, Japan). ProteinPilot (AB Sciex, Framingham, MA) was used to process the acquired time-of-flight (TOF) mass spectra data for peptide and protein identification. This software uses the Paragon database search algorithm for peptide identification and the Protein Group algorithm for protein grouping to yield the ProteinPilot generic format.
Western blotting
Proteins were separated on NuPage 4–12% Bis-Tris gels (Invitrogen) and transferred to polyvinylidene fluoride (PVDF) membranes using Novex iBlot PVDF transfer stacks (Life Technologies, Waltham, MA). The membranes were washed with PBS containing 0.1% Tween 20 (PBST), blocked with PBST containing 5% nonfat milk for 1 h, and then incubated overnight at 4°C with a rabbit anti-flotillin 2 antibody (#3436; Cell Signaling Technology, Danvers, MA; 1:5000) or a rabbit anti-flotillin 1 antibody (ab41927; Abcam, Cambridge, United Kingdom; 1:10,000). After washing with PBST, membranes were incubated with a biotin-conjugated donkey anti-rabbit IgG antibody (GE Healthcare; 1:20,000) for 1 h, washed with PBST, and then incubated with streptavidin horseradish peroxidase (GE Healthcare; 1:20,000) for 1 h. Specific bands were visualized using Immunostar LD reagent (Wako Pure Chemical, Osaka, Japan) and captured with a C-DiGit blot scanner (LI-COR, Lincoln, NE) according to the manufacturer's instructions.
Double immunofluorescence staining
Cells grown on poly-L-lysine coated culture coverslips (Matsunami Glass, Osaka, Japan) in a 24-well plate were fixed with 10% buffered formalin (Wako Pure Chemical) for 15 min, permeabilized with 0.3% Triton X-100 (Wako Pure Chemical) in PBS for 5 min, and blocked with 0.5% bovine serum albumin (BSA; Sigma Aldrich) in PBS for 1 h. Double immunofluorescence staining of flotillin 2 and Rab 5 was performed by incubating the coverslips with a mixture of rabbit anti-flotillin 2 antibody (#3436; Cell Signaling Technology; 1:500) and mouse anti-Rab 5 antibody (sc-46692; Santa Cruz Biotechnology; 1:200) at 4°C overnight. Coverslips were then incubated with a mixture of Alexa Fluor 594-conjugated chicken anti-rabbit IgG antibody (Life Technologies; 1:1000) and Alexa Fluor 488-conjugated goat anti-mouse IgG antibody (Life Technologies; 1:1000) for 1 h at room temperature. Double immunofluorescence staining of flotillin 2 and bovine Tg was performed by incubating the coverslips with a mixture of rabbit anti-flotillin 2 antibody (#3436; Cell Signaling Technology; 1:500) and mouse anti-bovine Tg antibody (AM20511PU-N; Acris Antibody, San Diego, CA; 1:500), which does not react with endogenous rat Tg, overnight at 4°C, then with a mixture of Alexa Fluor 488-conjugated goat anti-rabbit IgG antibody (Life Technologies; 1:1000) and Alexa Fluor 594-conjugated goat anti-mouse IgG antibody (Life Technologies; 1:1000) for 1 h. The nuclei were counterstained with Hoechst 33258 (Life Technologies; 1:1000). Cover slips were placed with fluorescence mounting medium (Dako, Tokyo, Japan). Immunofluorescence was visualized and the images were captured with an all-in-one BZ-9000 Generation II fluorescence microscope (KEYENCE, Osaka, Japan).
Knockdown of Flot1 or Flot2
Expression knockdown was performed by transfecting cells with predesigned siRNA to specifically target Flot1 (s134414; Life Technologies) or Flot2 (s136235; Life Technologies) and Lipofectamine RNAiMAX reagent (Life Technologies) according to the manufacturer's instructions. Control siRNA (#4390843; Life Technologies) was also used. Briefly, cells were grown in six-well plates to 70% confluence before the culture medium was changed with optical medium that lacked bovine serum or penicillin-streptomycin supplementation. For transfection, 2 μL of 5 nM siRNA with 5 μL of Lipofectamine RNAiMAX reagent was incubated in 250 μL of optical medium for 10 min at room temperature, and the mixture was gently pipetted into each plate well. Transfection was performed 24 h prior to Tg treatment.
Total RNA isolation and real-time polymerase chain reaction
Total RNA was purified using the RNeasy Plus Mini Kit (Qiagen, Hilden, Germany), and cDNA was synthesized using the High-Capacity cDNA Reverse Transcription Kit (Applied Biosystems, Foster City, CA) as previously described (10). Real-time polymerase chain reaction (PCR) was performed using Fast SYBR Green Master Mix (Applied Biosystems) and the StepOnePlus Real-Time PCR System (Applied Biosystems) according to the manufacturer's instructions. The sequences of PCR primers were as follows: Tg forward, 5′-GGC CTC CAC CTT CAC TCA AA-3′; Tg reverse, 5′-CAC TTG GTG GGA CAT CGG AA-3′; Slc5a5 forward, 5′-CTA CCG TGG GTG GTA TGA AGG-3′; Slc5a5 reverse, 5′-TGC CAC CCA CTA TGA AAG TCC-3′; Tpo forward, 5′-CAC GGC TTA CCA GGC TAC AA-3′; Tpo reverse, 5′-GCC TCC CAA CCA GAC ATC AA-3′; Duox2 forward, 5′-CAG CGC TAC GAC GGC TGG TTT A-3′; Duox2 reverse, 5′-CCC AAG CAC TGT GCG GTT GT-3′; Duoxa2 forward, 5′-TCA GCG TAC CGC TGC TCA TCG T-3′; Duoxa2 reverse, 5′-ACC AAC CAG AAC CAG CGC GAG T-3′; Nkx2-1 forward, 5′-ACC TTA CCA GGA CAC CAT GC-3′; Nkx2-1 reverse, 5′-TAC TTC TGC TGC TTG AAG CG-3′; Foxe1 forward, 5′-CCT TTA AGG AGG CGA AGC CA-3′; Foxe1 reverse, 5′-TGA AGA TAC AGC CCA AGC GG-3′; Pax8 forward, 5′-CAG GAC ACA CAC TGA TCC CC-3′; Pax8 reverse, 5′-GCA ATT CCT AGG AGC CCG TT-3′; Gapdh forward, 5′-ACA GCA ACA GGG TGG TGG AC-3′; and Gapdh reverse, 5′-TTT GAG GGT GCA GCG AAC TT-3′. The relative mRNA expression levels were normalized against corresponding Gapdh expression levels. Real-time PCR analysis was carried out at least in triplicate and repeated at least three times.
Statistical analysis
All experiments were repeated at least three times with different batches of cells, and the mean ± standard deviation of these experiments was calculated. The significance of the differences between experimental values was determined by an unpaired two-tailed t-test where p < 0.05 was significant.
Results
Identification of flotillins as novel Tg-binding proteins by mass spectrometry
To identify proteins that could recognize and interact with follicular Tg, Tg-bound proteins derived from bovine Tg-treated FRTL-5 cells were first immunoprecipitated using an anti-Tg antibody, which has been shown to react effectively with endocytosed bovine Tg in thyrocytes (10). The immunoprecipitated proteins were then subjected to mass spectrometry (MS) analysis. Immunoprecipitates from cells that were not incubated with Tg were used as a negative control. In total, 10 proteins were predicted to be Tg-binding proteins based on the presence of at least three high-confidence peptides (Table 1). Among these proteins, flotillin 1 (isoform 1) and flotillin 2, two homologous integral membrane proteins, ranked high in the total data analysis (Table 1).
Proteins predicted with at least three high-quality peptides are listed.
A measure of the total amount of evidence for a detected protein calculated from the peptide confidence of all detected peptides that were used to predict the protein.
Number of high-quality peptides detected with a confidence ≥95.
Percentage of matching amino acids from identified peptides with confidence ≥95 divided by the total number of amino acids in the protein sequence.
Consistent with the protein prediction, large amounts of flotillin 1 and flotillin 2 were detected in the immunoprecipitated Tg-bound proteins derived from Tg-treated cells (Fig. 1, lane 2) relative to proteins recovered from cells without Tg treatment (Fig. 1, lane 1) or those without Tg immunoprecipitation (Fig. 1, lane 4). Although the binding affinity between flotillins and Tg should be evaluated by solid phase binding assays, flotillins were identified as an abundant Tg-binding protein in FRTL-5 cells. Flotillin 1 and flotillin 2 were co-immunoprecipitated in Tg-bound protein complexes, which is in agreement with the notion that flotillins exist as homo- and hetero-oligomers in specialized cholesterol-enriched membrane microdomains termed lipid rafts (14 –16). These flotillin-containing lipid rafts are important in an endocytic mechanism that is distinct from the clathrin-mediated pathway. Moreover, flotillin-enriched lipid rafts are thought to participate in intracellular traffic and signal transduction in fibroblasts (17 –19), adipocytes (20,21), hepatocytes (22), neurons (23 –25), and T lymphocytes (26,27), but their functions in thyrocytes are unclear. Therefore, this study sought to elucidate a potential role for flotillin-containing lipid rafts in Tg endocytosis and in the negative-feedback effect of Tg using rat thyroid FRTL-5 cells.

Endogenous flotillins bind with endocytosed bovine thyroglobulin (Tg) in FRTL-5 cells. Whole cellular protein extracts were prepared from FRTL-5 cells incubated with (+) or without (–) 5 mg/mL of bovine Tg for 48 h. Immunoprecipitation (IP) was performed by incubating protein extracts with or without an anti-bovine Tg antibody in the presence of Protein A Sepharose beads, which were collected after IP and used directly in Western blotting for flotillin 1 or flotillin 2 as described in the Materials and Methods. Whole cell protein extracts were used as an input control.
Flotillin redistributes from cell membranes to endosomes together with endocytosed Tg
Compatible with their putative roles in mediating endocytosis, flotillin-containing lipid rafts were shown to have a dynamic distribution upon internalization of certain “cargoes,” in that they bud into cells from the cell membrane and then translocate to various cytoplasmic vesicular compartments, for example endosomes, lysosomes, phagosomes, and exosomes (15,19,28 –30). To detect such a redistribution of flotillin-containing lipid rafts that may occur during Tg endocytosis, double immunofluorescence staining for flotillin 2 and Rab 5, a protein marker for early endosomes (31), was performed. In agreement with its cell membrane localization, before Tg addition FRTL-5 cells were rimmed with flotillin 2 (Fig. 2A) without associating Rab 5 (Fig. 2B and C). After 6 h of Tg treatment, flotillin 2 staining on the cell membrane decreased while granular staining of the cytoplasm increased (Fig. 2D), which is consistent with a redistribution of flotillin 2 to endosomes. Moreover, much of the granular staining overlapped with Rab 5 staining (Fig. 2E and F). This granular co-localization of flotillin 2 and Rab 5 was observed even at high magnification (Fig. 2F, inset), indicating that the redistribution of flotillin 2 is closely related to the formation and trafficking of endosomes during Tg endocytosis.
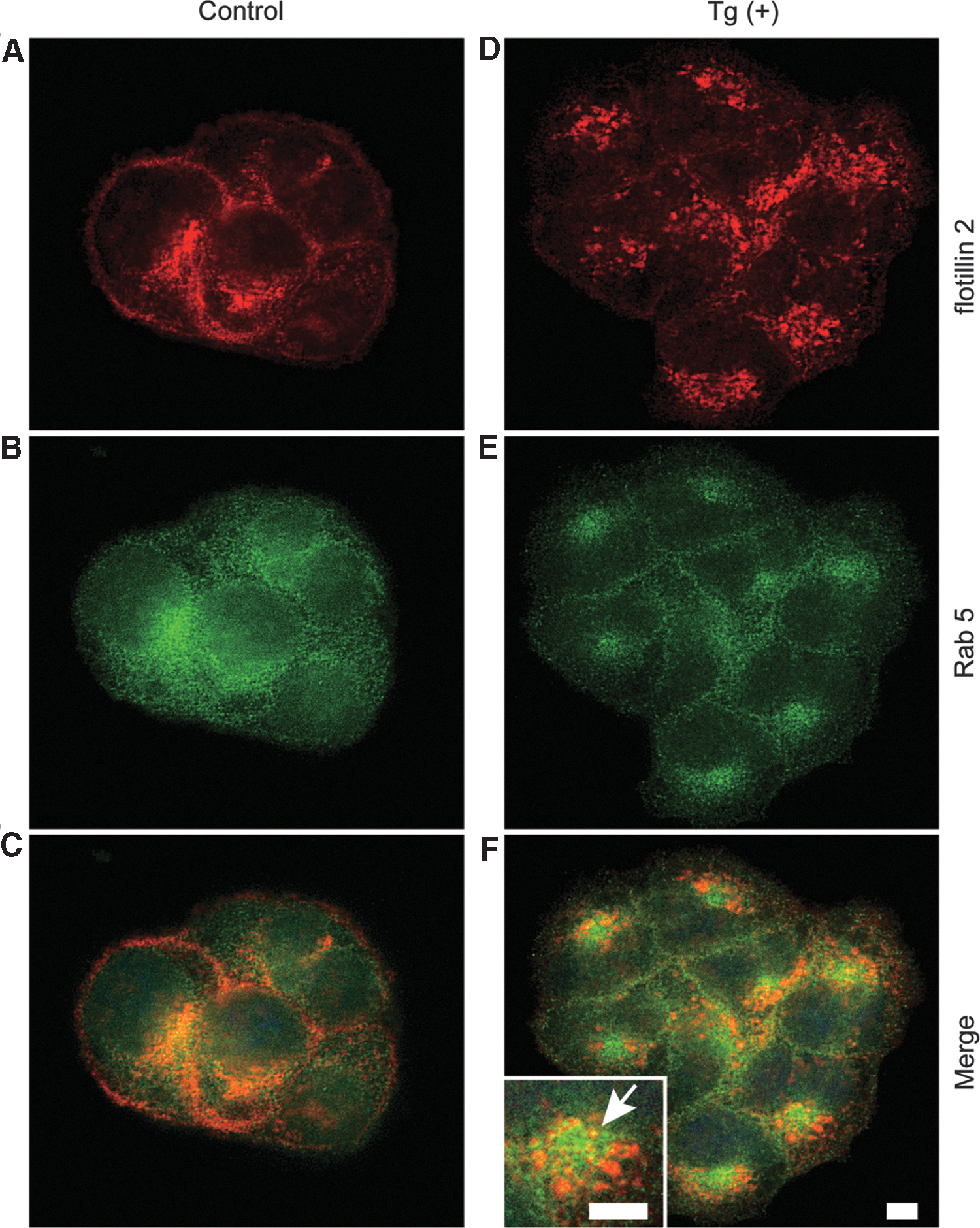
Tg induces redistribution of flotillin 2 from cell membranes to the endosomes. Control cells or cells incubated with 5 mg/mL of Tg for 6 h were subjected to double immunofluorescence staining for flotillin 2 (red) and Rab 5 (green). The arrow indicates typical co-localization of flotillin 2 and Rab 5 observed at high magnification (yellow). Bars: 10 μm.
Double immunofluorescence staining was performed using an anti-flotillin 2 antibody and an anti-bovine Tg antibody that does not cross react with endogenous rat Tg (Supplementary Fig. S1; Supplementary Data are available online at
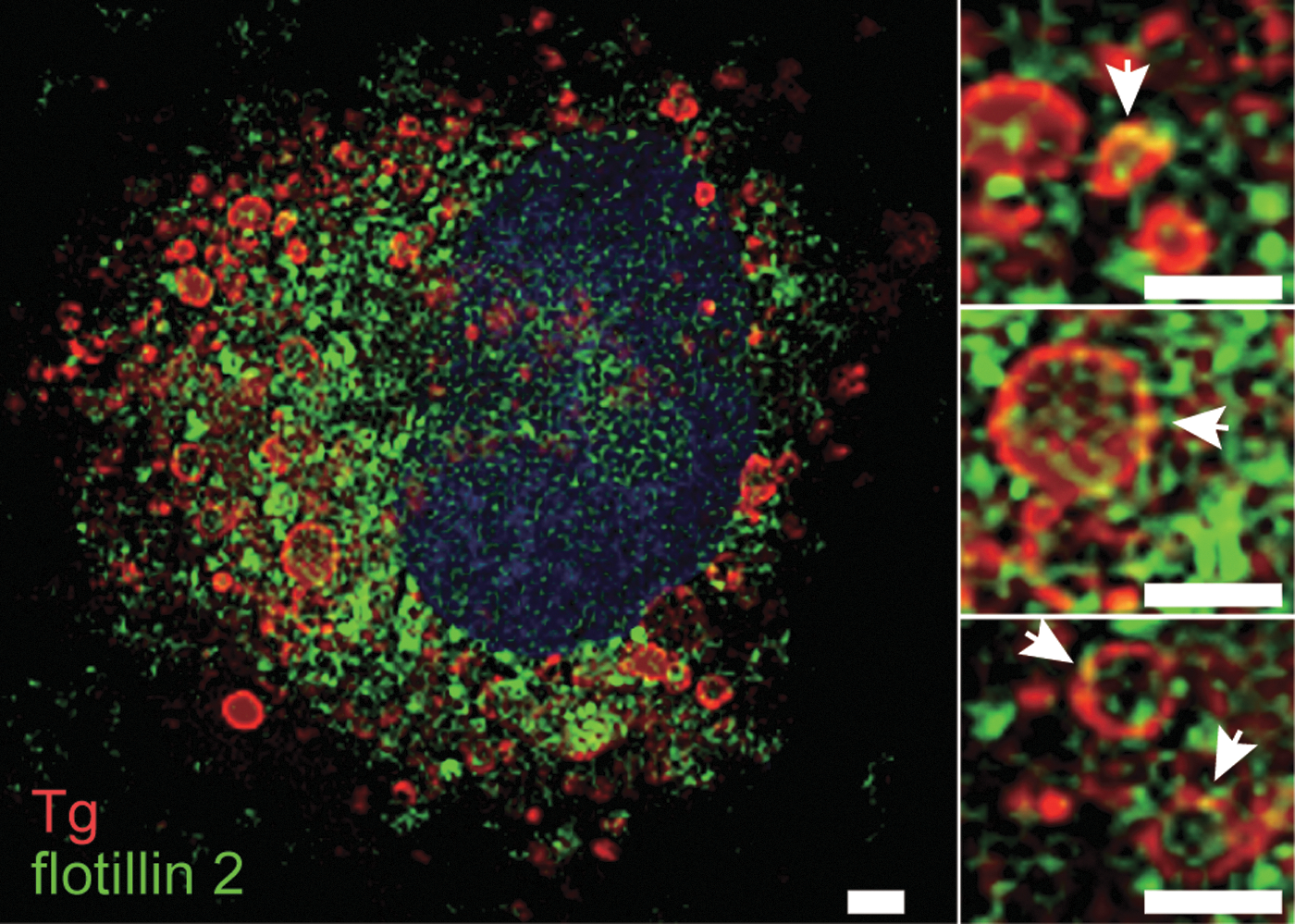
Endocytosed bovine Tg and endogenous flotillin 2 co-localize in FRTL-5 cells. Cells incubated with 5 mg/mL of Tg for 24 h were subjected to double immunofluorescence staining for bovine Tg (red) and flotillin 2 (green). The nuclei were counterstained (blue). Arrows indicate typical co-localization of endocytosed Tg with flotillin 2 (yellow). Bars: 5 μm.
MβCD inhibits Tg endocytosis in FRTL-5 cells
In addition to flotillin-containing lipid rafts, caveolae are also known as a membrane lipid raft micro-domain (32 –34). Both flotillin-containing lipid rafts and caveolae have similar lipid compositions that are enriched in cholesterol, but they are distinct in their membrane-coated proteins, with flotillin (16) and caveolin (33), respectively, predominating. However, caveolin mRNA could not be detected by real-time PCR or in a previously performed DNA microarray (data not shown), which is in accordance with previous reports (35,36). Thus, flotillin-containing lipid rafts, rather than caveolae, appear to be the dominant lipid raft variety on the FRTL-5 cell membrane. To test whether flotillin-containing lipid rafts are required for Tg endocytosis, MβCD was used to disrupt lipid raft structures in FRTL-5 cells (37). In control cells, intracellular vesicles could be observed by phase-contrast microscopy following Tg addition, and these vesicles gradually developed over time (Fig. 4A, left panels). However, incubation with 5 mM of MβCD for 2 h prior to Tg addition abolished the formation and development of such intracellular vesicles in Tg-treated cells (Fig. 4A, right panels). Immunofluorescence staining for bovine Tg in control cells demonstrated that by 6 h, there was accumulation of internalized Tg, which became more abundant 24 h after adding bovine Tg to the culture medium (Fig. 4B, left panels). In contrast, endocytosed Tg was barely detectable in MβCD-pretreated cells, even 24 h after Tg treatment (Fig. 4B, right panels). These results suggest an indispensable role for flotillin-containing lipid rafts in Tg endocytosis.
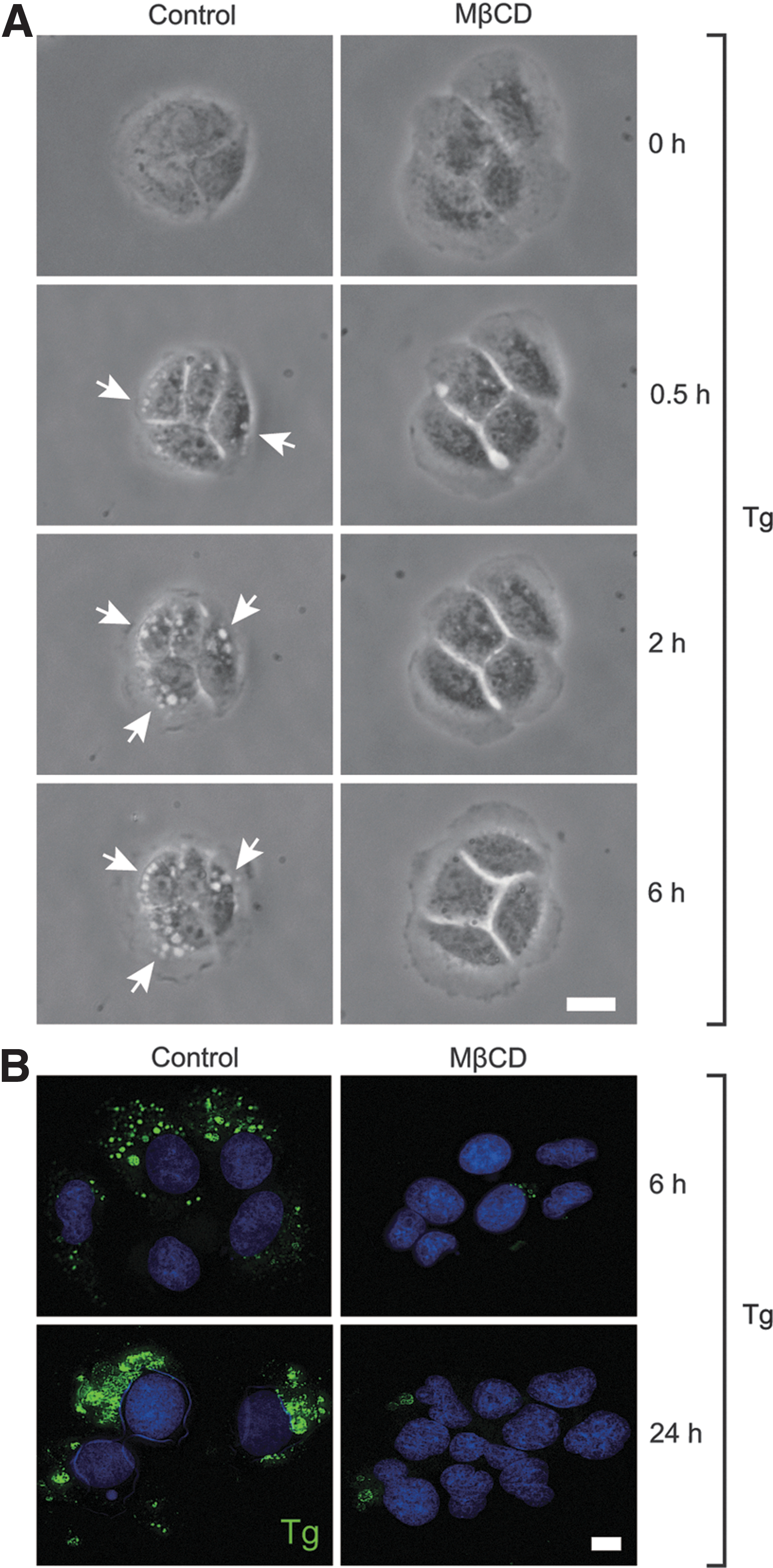
Lipid raft disrupter methyl-β-cyclodextrin (MβCD) inhibits Tg endocytosis in FRTL-5 cells. Control cells or cells pre-treated with 5 mM of MβCD for 2 h were incubated with 5 mg/mL of Tg. Inverted phase-contrast microscope images were taken at the indicated time after Tg addition (
MβCD treatment or flotillin knockdown abolished the negative-feedback effect of Tg on thyroid-specific gene expression
Next, the study whether flotillin-containing lipid rafts are also required for Tg to exert its negative-feedback suppression of thyroid-specific gene expression. First, the effect of Tg on thyroid-specific gene expression in the presence or absence of MβCD treatment was tested. As verified earlier (2,4 –8,10), in the absence of MβCD treatment, Tg at follicular concentrations strongly suppressed the expression of thyroid-specific genes, including Tg, Slc5a5, Tpo, Duox2, Duoxa2, Nkx2-1, Foxe1, and Pax8, as demonstrated by real-time PCR analysis (Fig. 5, open bars). However, this suppressive regulation of thyroid-specific gene expression by Tg was abolished in cells pre-incubated with MβCD (Fig. 5, solid bars).
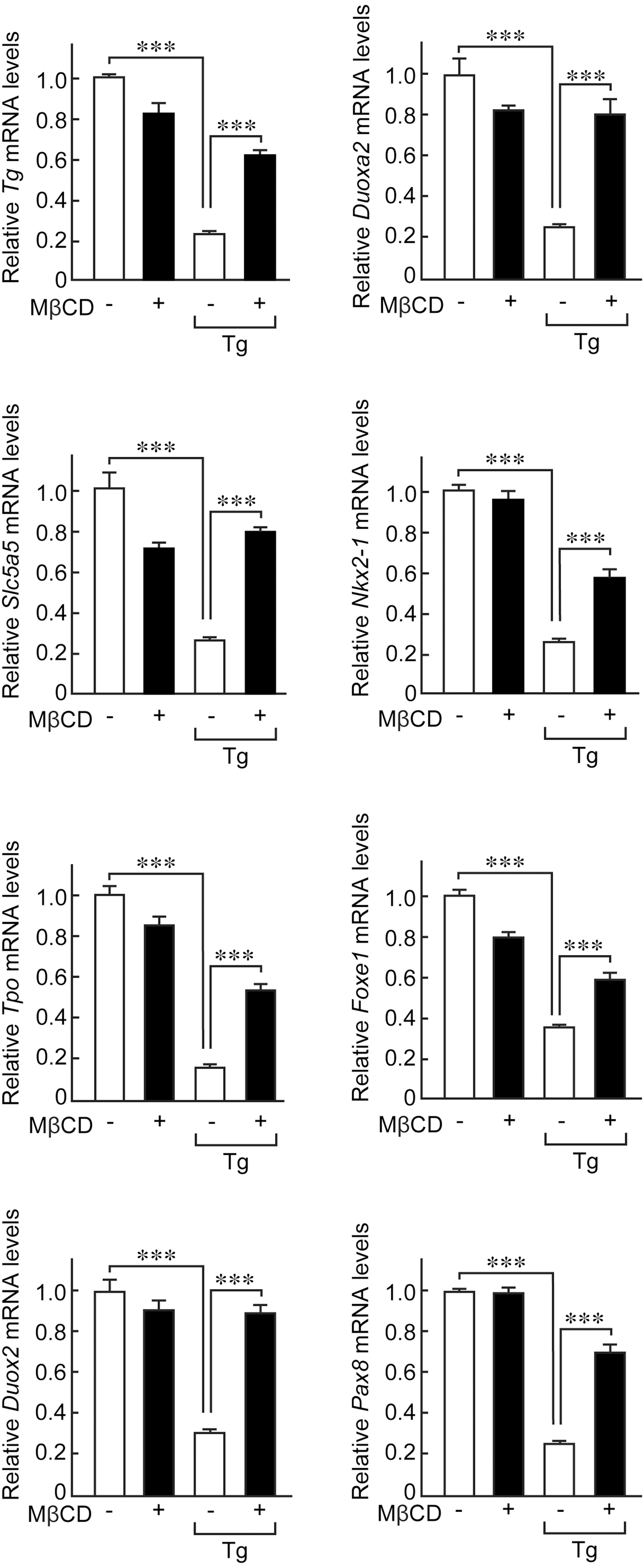
MβCD abolishes negative-feedback suppression of thyroid-specific gene expression by Tg. Control cells or cells pre-treated with 5 mM MβCD for 2 h were incubated with 5 mg/mL of Tg for 24 h. Total RNA was extracted and mRNA levels of thyroid-specific genes Tg, Slc5a5, Tpo, Duox2, Duoxa2, Nkx2-1, Foxe1, and Pax8 were evaluated using real-time polymerase chain reaction (PCR). mRNA levels of each gene were normalized against those of Gadph and the values shown are relative to the control. ***p < 0.001.
Furthermore, the essential role of flotillins in the Tg effect was confirmed through knockdown of flotillin 1 or flotillin 2 gene expression by specific siRNAs. The negative-feedback suppression of the thyroid-specific genes Tg, Slc5a5, Tpo, Duox2, Duoxa2, Nkx2-1, Foxe1, and Pax8 by Tg was significantly inhibited in flotillin 1- or flotillin 2-knockdown cells (Fig. 6). These results indicate that flotillin binding with Tg that was identified by MS analysis was not due to a non-specific interaction, and suggest that flotillin-containing lipid rafts play an important role in the recognition of follicular Tg and the negative-feedback effect of Tg on thyroid-specific gene expression in thyrocytes.
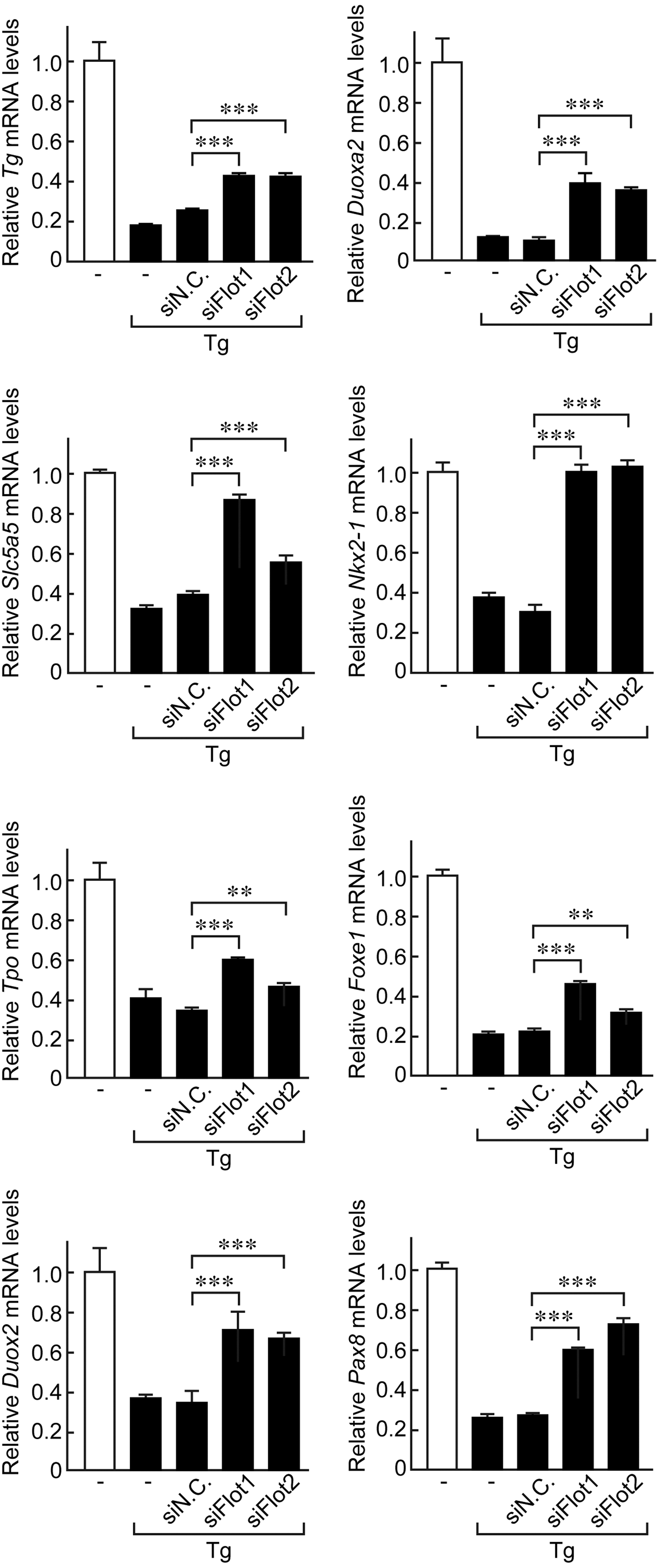
Knockdown of flotillin 1 or flotillin 2 by specific siRNAs significantly inhibits negative-feedback suppression of thyroid-specific gene expression by Tg. Control cells or cells transfected with siRNA against flotillin 1 or flotillin 2, or negative control siRNA were incubated with 5 mg/mL of Tg protein for 24 h. Total RNA was extracted and mRNA levels of thyroid functional genes, including Tg, Slc5a5, Tpo, Duox2, Duoxa2, Nkx2-1, Foxe1, and Pax8 were evaluated using real-time PCR. mRNA levels of each gene were normalized against those of Gadph, and values shown are relative to the control. **p < 0.05; ***p < 0.001.
Discussion
This study first identified flotillin 1 and flotillin 2, two characteristic lipid raft proteins, as novel Tg-binding proteins in FRTL-5 cells through MS analysis. It further confirmed that flotillins physically interact with endocytosed Tg and redistribute from the cell membrane to endosomes during the internalization of exogenous Tg. Disruption of lipid rafts by MβCD treatment or knockdown of flotillin 1 or flotillin 2 by specific siRNAs significantly abolished the negative-feedback effect of Tg on thyroid-specific gene expression levels. Thus, these results suggest that flotillin-containing lipid rafts play an essential role in the negative-feedback effect of accumulated follicular Tg to suppress gene expression that is necessary for TH synthesis.
Among the Tg-binding proteins identified in the MS analysis, flotillins were chosen for further examination, particularly because flotillin-containing lipid rafts have been shown to participate in various signaling pathways, for example insulin signaling (20), epithelial growth factor (EGF) signaling (15,17,19), p38 mitogen-activated protein kinase (MAPK) signaling (18), T-cell receptor signaling (26), and signal transduction by neurotransmitters (23 –25). Moreover, their function appears to be substantialized largely by the interaction of flotillins with their “cargoes,” as flotillin knockdown impairs relevant signal transduction pathways (15,18,21 –23,25,26,28,30,38,39). Co-assembly of flotillins has been shown to be sufficient to generate de novo invaginated membrane microdomains, distinct from caveolae, that are dynamic and can bud into the cell (29). In addition to these previously reported signaling pathways, this study reports for the first time a novel role for flotillin-containing lipid rafts in the recognition of Tg in the follicular colloid and its signal transduction in thyrocytes.
The discovery of this autoregulatory effect of follicular Tg on thyroid function has provided significant new findings and insights that may allow the conventional understanding of how follicular function is regulated to be revised, especially given that TSH was believed to be an unparalleled master regulator of thyroid follicles. Remarkably, this newly recognized negative-feedback effect of follicular Tg is sufficiently significant to counteract and even reverse TSH action in thyrocytes (3 –5,7,9), which thus forms a more “symmetrical” model: TH biosynthesis in thyrocytes is not only stimulated by TSH signals from the basolateral side that is essentially uniform among all follicles, but is also restrained by follicular Tg signals arising from the apical side depending on the concentration of Tg that accumulates in each follicle.
Naturally, there is the question of whether follicular Tg signals are mediated through Tg receptors on the apical membrane of thyrocytes. The asialoglycoprotein receptor is thought to serve as an apical receptor for endocytosis of colloidal Tg (40). Megalin/gp330 is well established as an apical receptor that mediates Tg transcytosis to prevent hormone-poor Tg from competing with hormone-rich Tg in the lysosome pathway, thus involved in the regulation of TH secretion (41 –45). However, previously described Tg receptors (40,45 –47) are not particularly specific for Tg, and none of them seem to commit strongly to the transduction of Tg signals, as the ligands for these receptors did not reproduce the effects of Tg seen here (3,48). On the other hand, high-affinity receptors presumably are not needed for Tg endocytosis, since Tg is the most abundant product in the follicular lumen. The observation that Tg uptake in FRTL-5 cells is not a saturable process (43,49) supports the notion that nonspecific fluid-phase pinocytosis is the prevailing form of Tg endocytosis in thyrocytes.
The presence of lipid rafts in thyrocytes was first suggested in 1988 (before the concept of lipid rafts was formally developed) when Barriere et al. demonstrated that cholesterol contents are heterogeneously distributed in thyrocytes in vivo, and that cholesterol is highly abundant in the apical membrane (50). Small apical vesicles and colloid droplets generally have a high cholesterol concentration in their membranes (50), suggesting that some cholesterol-enriched membrane micro-domains are involved in colloid reabsorption. In 2004 Costa et al. reported that human thyrocytes have caveolae in the apical membrane, but not the basolateral membrane (36). Intriguingly, relative to wild-type mice, caveolae-deficient mice have significantly increased expression levels of Tg, TPO, and DUOX (51). This phenotype is unlikely to be due to dysfunctional TSH signaling (36), but instead could be explained by the potential abrogation of the negative-feedback effect of Tg. Here, flotillin-containing lipid rafts, rather than caveolae, were shown to be the dominant type of lipid raft in FRTL-5 cells. This divergence may result from difference between the monolayer culture and non-polarity of rat thyroid FRTL-5 cells and the mouse thyroid in vivo.
Some previously reported Tg receptors, including megalin, were not detected in the current MS analysis. This outcome might be due to the inclusion of EDTA in the protein lysis buffer that can inhibit the binding of megalin with Tg (52). Nevertheless, flotillins were identified, which are characteristic components of lipid rafts, as novel Tg-binding proteins. The inhibition of the negative-feedback effect of Tg on thyroid-specific gene expression that arises following disruption of lipid rafts by MβCD treatment (Fig. 5), which also almost completely abrogates Tg endocytosis (Fig. 4), suggests that Tg signals likely require an actual internalization of Tg molecules into thyrocytes. Thus, the endocytosed Tg and/or its degraded fragments should be crucial for signal transduction to occur. The inhibitory effect of MβCD on the negative-feedback effect of Tg differed among the thyroid-specific functional genes that were examined. The negative feedback effect of follicular Tg on thyroid-specific gene expression also differed somewhat between genes (7). This difference may reflect the complex transcriptional regulation of thyroid-specific functional gene expression by multiple transcription factors and signaling pathways. As such, further investigation will be needed to determine the mechanisms responsible for the Tg effect. The significant inhibition of the effect of Tg by knockdown of flotillin 1 or flotillin 2 (Fig. 6) obviously can be explained by a failure of flotillin-containing lipid raft formation. Moreover, recent studies have begun to address the role of flotillins beyond endocytosis, for example flotillins have been conclusively shown to be novel signaling molecules in the MAPK signal pathway (53,54). Thus, whether flotillins per se serve as signaling molecules in Tg signal pathways remains an open question. It was previously reported that in the absence of TSH, insulin, and serum, Tg significantly induces FRTL-5 cell growth without activating the cyclic AMP/protein kinase A (PKA) pathway, but instead activates the MAPK pathway (12,55). However, the signaling pathway that mediates the negative feedback suppression of Tg on the expression of functional genes in the thyroid in the presence of TSH remains to be determined.
This study identified flotillins as novel Tg-binding proteins, and shows that flotillin-containing lipid rafts play an important role in the negative feedback regulation of thyroid-specific gene expression by Tg in FRTL-5 cells. In addition to flotillins, several other proteins were also detected with high scores in MS analysis, including the 78 kDa glucose-regulated protein (Table 1). These proteins are also candidate Tg receptors that will be investigated in the future. Further studies are also needed to clarify the possible role of flotillins as direct signaling molecules that mediate Tg action, or other hitherto unrecognized downstream molecules that are essential for Tg signal transduction. Nevertheless, identification of the first step of Tg recognition, that is, the proteins that mediate initial contact with the thyrocytes, may further the understanding of the action of Tg in the context of TH synthesis. Such an understanding will also contribute to the elucidation of certain thyroid disorders that may be caused by abnormalities in Tg regulation, and to the development of therapeutic modalities to modulate signaling pathways that are initiated by follicular Tg.
Footnotes
Acknowledgments
This work was supported in part by a JSPS KAKENHI Grant (#15K09444 to K.S.). The authors thank the generous help from Keyence Co. (Osaka, Japan) for the use of the all-in-one fluorescence microscope BZ-9000 Generation II.
Author Disclosure Statement
The authors have nothing to disclose.