
Abstract
Background:
Graves' disease (GD) and Hashimoto's disease (HD) are autoimmune thyroid disorders distinguished by the presence or absence of antithyrotropin receptor (TSHR) antibodies (TRAb). TSHR gene polymorphisms determine the amount of TSHR expressed, which may in turn influence TRAb production. The FANTOM5 project identified six GD-associated single nucleotide polymorphisms (SNPs) within the enhancer regions of the TSHR and unknown genes. This study examined the association of 11 TSHR and unknown gene polymorphisms, five of which are located in TSHR enhancer regions, with the development and prognosis of GD and HD.
Methods:
SNPs of the TSHR and unknown genes were genotyped in 180 GD patients, including 62 patients with intractable GD and 48 patients with GD in remission; 151 HD patients, including 65 patients with severe HD and 40 patients with mild HD; and 111 healthy controls.
Results:
The rs4411444 GG genotype and G allele, the rs2300519 AA genotype, and the rs179247 AA genotype and A allele were more frequent in GD patients than they were in controls. These same genotypes and alleles, in addition to the rs2300519 A allele and rs4903961 GG genotype and G allele, were more frequent in patients with intractable GD than they were in controls and patients with GD in remission. Interestingly, the rs2300519 TT genotype and T allele, rs4903961 CC genotype and C allele, and rs179247 GG genotype, all of which are minor genotypes and alleles among the evaluated SNPs, were more frequent in HD patients than they were in controls, but there were no differences in the frequencies of these genotypes and alleles between patients with severe HD and mild HD. Among the evaluated SNPs, the rs4411444 GG genotype and the rs4903961 C allele in the enhancer regions of the TSHR gene were most strongly associated with the development of GD, especially intractable disease, and that of HD, respectively.
Conclusions:
Among the evaluated TSHR gene SNPs, the rs4411444 GG genotype and the rs4903961 C allele in the enhancer regions of the TSHR gene were most strongly associated with the development of GD, especially intractable disease, and that of HD, respectively.
Introduction
A
AITDs can be characterized by lymphocytic infiltration in the thyroid and the production of autoantibodies to thyroid-specific antigens, such as thyrotropin receptor (TSHR), thyroglobulin (Tg), and thyroid peroxidase (TPO) (13 –17). GD is diagnosed by the presence of an anti-TSHR antibody (TRAb), including thyroid-stimulating antibodies (TSAb), which stimulate thyroid epithelial cells to produce thyroid hormones and cause hyperthyroidism (18 –20). HD is diagnosed by the absence of TRAb and the presence of other thyroid autoantibodies, including anti-Tg antibodies (TgAb) and/or anti-TPO antibodies (TPOAb) (21). Sometimes HD transitions to GD, as determined by the appearance of TRAb, and GD remission with antithyroid drug therapy is associated with the disappearance of TRAb. Therefore, the propensity to develop GD is determined by poor immunological tolerance of the TSHR antigen and production of a TSHR autoantibody (22). A genome-wide association study identified strong associations of the TSHR and the major histocompatibility complex (MHC) class II variants with persistently positive TRAb and GD, and the authors suggested that shedding the A subunit of the TSHR may result in TSAb production (23,24).
Recently, the FANTOM5 project identified 43,011 enhancers within the majority of human cells and tissues (
This study aimed to clarify the association of genetic differences in the TSHR with the development and prognosis of AITDs by genotyping previously described putative GD-associated SNPs: six within the enhancer regions of the TSHR, unknown genes, and five SNPs in the intron 1 and intron 7 regions of the TSHR. To analyze the function of these polymorphisms, the expression level of TSHR mRNA and serum levels of TRAb were examined.
Materials and Methods
Subjects
The presence/absence of each polymorphism was determined in 151 HD patients, 180 GD patients, and 111 healthy volunteers (control subjects). HD patients were positive for TgAb and/or antithyroid microsomal antibodies (McAb). GD patients were positive for TRAb at diagnosis and had a clinical history of thyrotoxicosis during HD. Healthy volunteers were euthyroid and negative for thyroid autoantibodies.
Among HD patients, 65 patients who developed moderate to severe hypothyroidism before 50 years of age and who were treated with thyroxine (severe HD) and 40 patients older than 50 years of age who were euthyroid and left untreated (mild HD) were genotyped. Among GD patients, 62 euthyroid patients who had been treated with methimazole for at least five years and who were still positive for TRAb (intractable GD) and 48 patients who had maintained a euthyroid state and who were negative for TRAb for more than two years without medication (GD in remission) were genotyped. Seventy GD patients and 46 HD patients who could not be categorized by the abovementioned parameters were defined as the “GD unclear group” and “HD unclear group,” respectively.
All patients with AITD and the control subjects were Japanese and were unrelated to each other. Genomic DNA was isolated from EDTA-treated peripheral blood mononuclear cells (PBMCs) using commercially available kits (QIAamp DNA Blood Mini Kit; Qiagen, Tokyo, Japan). Written informed consent was obtained from all subjects, and the study protocol was approved by the Ethics Committee of Osaka University, Osaka, Japan.
Genotyping GD-associated SNPs
The SNPs genotyped in this study are described in Table 1 and Figure 1. SNPs in enhancer regions were named SNP E1-6, and those in the intron 1 and intron 7 regions of the TSHR gene were named SNP1-5. The linkage of SNP E1-3 was analyzed using HaploView v4.2 software (Broad Institute, Cambridge, MA), and it was found that these SNPs were strongly linked (r 2 = 1.0). Therefore, only SNP E2 was genotyped. A polymerase chain reaction–restriction fragment length polymorphism (PCR-RFLP) method was used to genotype TSHR SNP E2, SNP E5, and SNP1. Briefly, the target sequence of each gene was amplified using PCR, and the PCR products were digested by restriction enzymes. A direct sequencing method was used for genotyping TSHR SNP E4, SNP2, SNP3, and SNP4. Briefly, the target sequence of each gene was amplified using PCR, and then the purified PCR products were sequenced using Big Dye thermosequenase (Bid Dye Terminator v1.1 cycle Sequencing Kit; Applied Biosystems, Carlsbad, CA). The dye-labeled purified PCR products were analyzed using a sequencer (Applied BioSystems 3100 Genetic Analyzer; Applied Biosystems). An amplification refractory mutation system-PCR (ARMS-PCR) method was used to genotype TSHR SNP5. Briefly, the PCR products containing only each target allele were amplified using four sequence-specific PCR primers. The sequences of PCR primer pairs and sequence primers, PCR conditions, and restriction enzymes used in this study are summarized in Table 2. TaqMan SNP genotyping assays (Applied Biosystems, Tokyo, Japan) were used to genotype SNP E6.
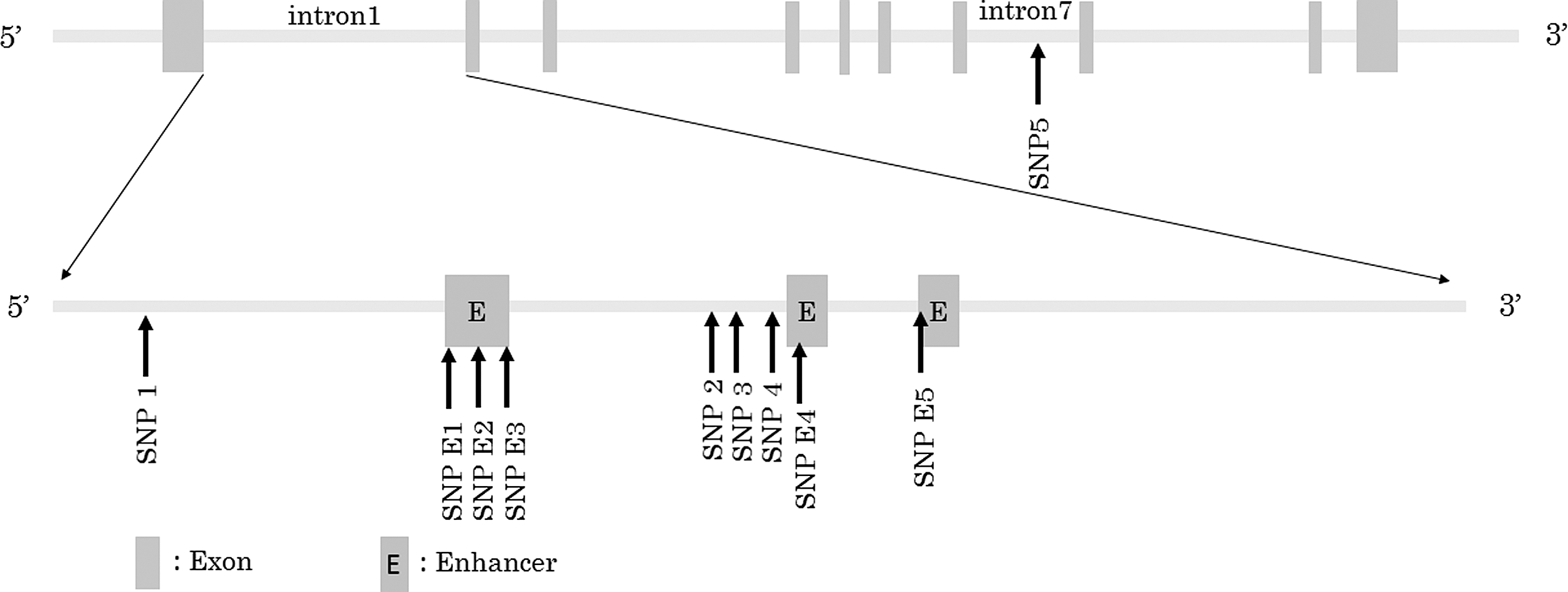
Locations of TSHR gene polymorphisms.
Asterisk indicates sequence primer.
PCR polymerase chain reaction; PCR-RFLP, PCR–restriction fragment length polymorphism; ARMS-PCR, amplification refractory mutation system-PCR.
mRNA extraction from PBMCs
The TSHR mRNA expression levels in PBMCs from 30 control subjects were analyzed to exclude the effect of medical treatment. Briefly, peripheral blood samples were collected in EDTA-treated tubes and PBMCs were isolated by density gradient centrifugation with Lymphoprep (Axis-Shield PoC As, Oslo, Norway) at 400 g for 30 min at room temperature. PBMCs were washed in sterile phosphate-buffered saline (PBS) and preserved in RNAlater solution (Ambion, Austin, TX) at –80°C until analysis. Total RNA was extracted from PBMCs with the mirVana™ PARIS™ KIT (Ambion), according to the manufacturer's protocol.
Quantitative real-time PCR
cDNAs were generated using the High Capacity cDNA Reverse Transcription Kit (Life Technologies, Tokyo, Japan). Real-time PCR was performed with TaqMan Gene Expression Assays (Life Technologies). The expression levels of TSHR mRNA were normalized to GAPDH. All reactions were performed in triplicate. The relative expression level of each mRNA was calculated using the ΔΔCt method and shown as the relative value to a particular sample.
Statistical analysis
A chi-square test was used to evaluate the significance of differences in the frequencies of genotypes and alleles among the groups. The Mann–Whitney U-test was used to analyze the difference between the serum levels of TRAb. Data were analyzed with JMP11 software (SAS Institute, Inc., Tokyo, Japan). Probability values of <0.05 were considered significant.
Results
Genotypes of TSHR gene polymorphisms
TSHR rs4411444 G/A polymorphism (SNP E2)
The frequencies of the GG genotype and G allele were significantly higher in GD patients than they were in controls (p < 0.0001 and p = 0.0008, respectively; Table 3). These frequencies were also significantly higher in GD patients than they were in HD patients (p < 0.0001 and p < 0.0001, respectively; Table 3). The GG genotype and G allele were significantly more frequent in intractable GD patients compared with controls (p < 0.0001 and p = 0.0003, respectively; Table 4) and were also significantly more frequent in intractable GD patients compared with GD patients in remission (p = 0.0227 and p = 0.0427, respectively; Table 4).
AITD, autoimmune thyroid disease; GD, Graves' disease; HD, Hashimoto's thyroiditis; NS, not significant.
TSHR rs2300519 A/ T polymorphism (SNP E4)
The frequency of the AA genotype was significantly higher in GD patients than it was in controls (p = 0.0228; Table 3). The proportions of the AA genotype and A allele were also significantly increased in GD patients compared with those in HD patients (p = 0.0001 and p < 0.0001, respectively; Table 3). On the other hand, the TT genotype and T allele were significantly more frequent in HD patients than they were in controls (p = 0.0132 and p = 0.0366, respectively; Table 3). The frequencies of the AA genotype and A allele were significantly higher in intractable GD patients than they were in controls (p = 0.0032 and 0.0173, respectively; Table 4) and were also significantly higher in intractable GD patients compared with GD patients in remission (p = 0.0263 and p = 0.0243, respectively; Table 4).
TSHR rs4903961 G/C polymorphism (SNP E5)
The frequencies of the CC genotype and C allele were significantly higher in HD patients than they were in controls (p = 0.0126 and p = 0.0042, respectively; Table 3). The GG genotype and G allele were significantly more frequent in GD patients than they were in HD patients (p < 0.0001 and p < 0.0001, respectively; Table 3). The proportions of the GG genotype and G allele were significantly increased in intractable GD patients compared with those in controls (p = 0.0127 and p = 0.0214, respectively; Table 4) and were also significantly increased in intractable GD patients compared with GD patients in remission (p = 0.0147 and p = 0.0166, respectively; Table 4).
TSHR rs179247 A/G polymorphism (SNP1)
The frequencies of the AA genotype and A allele were significantly higher in GD patients than they were in controls (p = 0.0052 and p = 0.0158, respectively; Table 3), and were also significantly higher in GD patients than they were in HD patients (p = 0.0001 and p < 0.0001, respectively; Table 3). On the other hand, the GG genotype was significantly more frequent in HD patients than it was in controls (p = 0.0274; Table 3). The frequencies of the AA genotype and A allele were significantly higher in intractable GD patients compared with those in controls (p = 0.0003 and p = 0.0027, respectively; Table 4), and were also significantly higher in intractable GD patients compared with GD patients in remission (p = 0.0090 and p = 0.0027, respectively; Table 4).
TSHR rs541680762 C/G polymorphism (SNP2), rs137877366 A/G polymorphism (SNP3), and rs201206147 A/G polymorphism (SNP4)
No minor allele was found in genotyped subjects (data not shown).
TSHR rs2268475 T/C polymorphism (SNP5)
The frequency of the TT + TC genotype and T allele were significantly higher in GD patients than they were in controls (p = 0.0052 and p = 0.0068, respectively; Table 3). Furthermore, the population of the T allele was significantly increased in HD patients compared with those in controls (p = 0.0355; Table 3). No associations were found between this polymorphism and the intractability or severity of AITDs.
Genotypes of chromosome 6 polymorphism
rs6941355G/A polymorphism (SNP E6)
No difference was found in the genotype and allele frequencies of this polymorphism between each group of patients and controls (Tables 5 and 6).
mRNA expression levels of TSHR in PBMCs
The expression of TSHR mRNA was not detected in PBMCs, and the association between the TSHR mRNA expression levels and the examined SNPs could not be analyzed.
Serum levels of TRAb and genotyped polymorphisms
No associations were found between each of the genotyped SNPs and the serum levels of TRAb at the onset of the disease in GD patients (data not shown).
Discussion
This is the first report that has comprehensively examined the association of previously described putative GD-associated SNPs, including those within the enhancer regions of the TSHR gene and unknown genes, with the development and prognosis of GD and HD. It has been clarified for the first time that SNPs within the enhancer regions of the TSHR gene are most strongly associated with the development of GD, especially intractable disease, and HD.
In the case of SNP1, it was found that the frequencies of the AA genotype and the A allele were significantly higher in GD patients than they were in controls (Tables 3 and 4), as shown in previous studies (26,30). It was also found that those were significantly higher in intractable GD patients than they were in patients with GD in remission, and that the frequency of the GG genotype was higher in HD patients than it was in controls (Tables 3 and 4). These results are consistent with those of a previous study (31). Expression of ST4, which is a TSHR mRNA splice variant, was higher in subjects with the AA genotype than it was in those with the GG genotype (26), and increased ST4 expression may result in a greater level of shedding the A subunit and thus increased production of TSAb (23). Therefore, SNP1 may be associated with GD development and prognosis based on differences in the TSAb levels caused by changes in ST4 expression.
In the case of SNP E1–5 in the TSHR gene enhancer region, it was found for the first time that the frequencies of both major genotypes and alleles of these SNPs were higher in GD patients, especially in intractable GD patients, than they were in controls, and that the frequencies of both minor genotypes and alleles were higher in HD patients than they were in controls (Tables 3 and 4). Because these SNPs are near SNP1, their linkage was analyzed using HaploView v4.2 software, and it was found that SNP E2 is strongly linked to SNP1 (r 2 = 0.902). Among all of the genotyped SNPs in intron 1 of the TSHR (SNP1, E2, E4, and E5), SNP E2 was most strongly associated with the pathogenesis of GD. Thus, SNP E2 may directly affect the expression of the TSHR, ST4, and TSAb by altering transcription factor binding. GD is mainly an autoimmune thyroid disease in which most patients are positive for TgAb and TPOAb, in conjunction with the appearance of TRAb, including TSAb. GD remission can be achieved with antithyroid drug therapy in association with the disappearance of TRAb. Therefore, major genotypes and alleles of SNP E1–5, which were more frequent in GD patients, especially with intractable GD, than they are in controls, may induce higher TSHR gene expression and higher production of TSHR antigen for presentation to TSHR-specific T helper cells and B cells. Thus, TSAb would be produced more efficiently or abundantly, making it easier to break immunological tolerance of the TSHR antigen. By contrast, minor genotypes and alleles of these SNPs, which were more frequent in HD patients than they were in controls, may induce lower TSHR gene expression and lower production of the TSHR antigen for a weaker presentation to TSHR-specific cells and a lower impairment of immunological tolerance. The pathological condition of HD is usually characterized by an absent break of immunological tolerance to the TSHR antigen, and therefore the minor genotypes and alleles of these SNPs were considered to be more frequent in HD.
To test this hypothesis, transcription factor binding to TSHR enhancer regions, including SNP E1-6, was simulated using P-match software (
It has been suggested that TSHR mRNA is expressed not only on thyroid epithelial cells but also in PBMCs (32). Because normal thyroid tissue could not be obtained, the TSHR mRNA levels in PBMCs were examined from controls to clarify the effect of polymorphisms in TSHR expression. However, TSHR mRNA could not be detected in PBMCs. This is consistent with another study, which was unable to find relevant TSHR mRNA expression in PBMCs (26). These findings indicate that TSHR mRNA expression is limited in PBMCs, and for this reason, differences in TSHR expression among genotypes of TSHR SNPs could not be clarified.
Next, the association between SNPs in this study and the serum levels of TRAb at the onset of GD were analyzed. However, no associations were found between each of these SNPs and TRAb expression levels. In a previous study, the TSAb levels were increased, but the TRAb levels were not changed in experimental hyperthyroid mice (23). Therefore, it is supposed that some association may be found between the examined SNPs and TSAb levels.
Contrary to expectation, in the case of SNP5 in the intron 7 region, the major T allele was more prevalent not only in GD patients but also in HD patients compared with controls. Although SNP5 in GD has already been reported (29), this is the first study indicating the association between this SNP and susceptibility for HD. In other words, this SNP was associated with the development of AITD, including both GD and HD, through some unknown mechanism. This SNP is also located in a different gene, LOC101928462, whose function has not yet been characterized. SNP5 may affect the expression of this gene, and one could speculate that it could be involved in the development of AITD.
In conclusion, the major genotype and allele of each SNP (SNP1 and SNP E1-5) in the TSHR gene is associated with the development of GD, especially intractable GD. Among the SNPs, the rs441144 GG genotype (SNP E2) shows the most significant association. The minor genotype and allele of these polymorphisms is associated with the development of HD. Among them, the rs4903961 C allele (SNP E5) shows the most significant association. SNP5 in the TSHR gene is also associated with the development of GD and HD.
Footnotes
Acknowledgments
This work was supported by JSPS KAKENHI grant number 26293128.
Author Disclosure Statement
The authors declare that they have no conflict of interest.