
Abstract
Background:
Heterozygous mutations in the thyroid hormone receptor alpha (THRA) gene cause resistance to thyroid hormone alpha (RTHα), a disease characterized by variable manifestations reminiscent of untreated congenital hypothyroidism but a raised triiodothyronine/thyroxine ratio and normal thyrotropin levels. It was recently described that zebrafish embryos expressing a dominant negative (DN) form of thraa recapitulate the key features of RTHα, and that zebrafish and human receptors are functionally interchangeable.
Methods:
This study expressed several human thyroid hormone receptor alpha (hTRα) variants in zebrafish embryos and analyzed the resulting phenotypes.
Results:
All hTRα-injected embryos showed variable defects, including cerebral and cardiac edema likely caused by an aberrant looping during heart development, anemia, and an incomplete formation of the vascular network. Moreover, the hTRα-injected embryos presented severe defects of motorneurons and craniofacial development, thus affecting their autonomous feeding and swimming behaviors. Surprisingly, expression of all hTRα mutants had no detectable effect on thyrotropin beta and thyrotropin-releasing hormone transcripts, indicating that their DN action is limited on the thyroid hormone reception beta 2 targets at the hypothalamic/pituitary level in vivo. As previously described in vitro, treatment with high triiodothyronine doses can efficiently revert the observed defects only in embryos injected with missense hTRα variants.
Conclusion:
Injection of human THRA variants in zebrafish embryos causes tissue-specific defects recapitulating most of the RTHα clinical and biochemical manifestations. The described manipulation of zebrafish embryos represents a novel in vivo model to screen the functional consequences of THRA variants and the rescue potential of new therapeutic compounds.
Introduction
T
The molecular basis of RTHα has been investigated in animal models. The phenotypic alterations described in mouse models with artificially introduced TRα1 mutations are usually consistent with those reported in human cases. However, high TSH levels have been found in several murine models, in contrast to the findings reported in human patients with RTHα (12 –20).
The protein structure of the TRs is highly conserved across vertebrates, particularly in the DNA-binding (DBD) and ligand-binding (LBD) domains, reaching 90–95% of identity between human and zebrafish receptors. Therefore, in a previous study (21), embryos harboring endogenous DN mutations in the LBD of zebrafish TRα isoforms were generated by morpholino microinjection. Those morphant embryos recapitulated most of the key clinical and biochemical manifestations (e.g., growth and brain defects, delayed cartilage development, defective heart function, high T3/T4 ratio) described in RTHα patients. In these thraa morphants, increased pituitary dio2 and reduced dio3a and dio3b expression was observed, which explains the peculiar biochemical features found in RTHα. It was also reported that zebrafish and human TRs were functionally interchangeable, and that high T3 doses partially reverted the DN action of mutant hTRα in thraa morphants.
The present work developed a simplified model and evaluated the molecular and functional tissue consequences of several human THRA mutations (i.e., D211G (10), A263V (5), A382PfsX7 (3), E403X (1), and F397fs406X (2)) by directly forcing their expression in zebrafish embryos. The injected embryos were then subjected to a series of experiments in order to analyze the effects of hTRα mutations during zebrafish development, particularly focusing on tissues and functions that are affected in patients with RTHα. Finally, the ability of T3 treatment to rescue the disease phenotype of embryos injected with the different THRA variants was tested.
Materials and Methods
The amino acid alignment of the different zebrafish TR (zTR) isoforms and the human TRα1 (hTRα1) and hTRα2 proteins (including the localization of the THRA variants studied in this work) is reported in Supplementary Figure S1 (Supplementary Data are available online at
Fish line and maintenance
Zebrafish (Danio rerio) from wild-type (AB) and tg(gata1:dsRed)sd2; tg(kdrl:EGFP)S843 (22) adults were maintained in controlled conditions, and all procedures were performed in accordance with Italian and European regulations (D. Lgs no. 2014/26, implementation of the 2010/63/UE). As no experiments were foreseen beyond five days post fertilization (dpf), the Institutional Review Board approved the protocol. Zebrafish embryos, obtained from natural spawning, were raised and maintained according to established techniques (23) and staged according to morphological criteria (24). For whole mount in situ hybridization (WISH) and immunohistochemistry (IHC) experiments, from 24 hours post fertilization (hpf), the embryos were cultured in fish water containing 0.002% of 1-phenyl-2-thiourea (Sigma) that prevents embryonic pigmentation without side effects on TH synthesis (21).
mRNA synthesis, microinjection, and treatment of zebrafish embryos
The human THRA (ENST00000450525) mutant plasmids were linearized and in vitro transcribed using the mMESSAGE mMACHINET7 kit (Ambion), followed by RNA purification using the Megaclear kit (Ambion), according to the manufacturer's protocols. Each hTRα transcript was injected in zebrafish eggs at the one- to two-cell stage at a dose of 80 pg/embryo. Starting from 6 hpf, the injected embryos were treated with 20 μM of dimethyl-sulfoxide (DMSO; Sigma) or 20 μM of T3 (IBSA-Farmaceutici). Uninjected embryos treated with the same dose of DMSO were used as controls.
WISH and IHC
To study the cardiac and thyroid function, WISH was performed (25) using the following probes: cmlc2 (26), tg (27), tshba (28), dio2 (29), dio3a, and dio3b (30). The DIG-probes were detected using anti-DIG-alkaline phosphatase (AP) and NBT/BCIP or FastBlue staining, as previously described (27). For IHC, embryos were exposed to anti-mouse primary antibodies (1:200 of znp1, a4-1025, or zn5; ZIRC) (31) or rabbit anti-T4 (1:1000; ICN Biochemicals), and 1:200 of AlexaFluor 488 anti-mouse or anti-rabbit IgG (Invitrogen) as secondary antibody. After WISH and IHC, embryos were post-fixed in 4% paraformaldehyde (PFA), washed three times in phosphate-buffered saline (PBS) +0.1% Tween 20, rinsed in 85% glycerol/PBS, and then submitted to image acquisition.
RNA isolation and quantitative real-time polymerase chain reaction
Total RNA from pools of 50 embryos at different stages was extracted with Trizol Reagent (Invitrogen) and then reversed transcribed using the ImProm-II Reverse Transcription System (Promega) according to the manufacturers' instructions. Quantitative real-time polymerase chain reaction (qRT-PCR) was analyzed on an ABI7300 System (PerkinElmer Applied Biosystems) and SYBRGreen MasterMix (Invitrogen). The relative mRNA expression was measured and normalized against the housekeeping gene eef1α using the 2−ΔΔCt method. Values represent the mean ± standard deviation (SD) of three experiments, performed independently. The list of primers is available upon request.
Heart-rate counts of zebrafish embryos
Heart rates (beats per minute) were counted manually under the Leica MZFLIII stereomicroscope in injected embryos and uninjected controls at 2 dpf. Embryos were first anesthetized by adding 0.016% Tricaine (ethyl3-aminobenzoate methanesulfonate salt; Sigma), which did not have any effect on the zebrafish heart rate (32). Results are expressed as the mean ± SD of three experiments (15 embryos in each experiment).
Analysis of vascular development
To study the architecture of zebrafish vasculature and the circulating blood cells, tg(gata1:dsRed)sd2; tg(kdrl:EGFP)S843 double transgenic embryos were anesthetized with 0.016% Tricaine (Sigma) at 2 dpf and then submitted to image acquisition with a fluorescence or confocal microscope. The endogenous alkaline phosphatase (AP) activity was determined as previously described (33). Briefly, the embryos were fixed with 4% PFA at room temperature (RT) for 2 h, rinsed in PBS, and incubated overnight at 4°C. Embryos were then incubated in 100% MetOH for 2 h at RT, hydrated sequentially in PBS, and rinsed for 30 min in AP buffer (100 mM of Tris-HCl pH 9.5, 100 mM of NaCl, 50 mM of MgCl2, 1% Tween20), and finally stained with NBT/BCIP diluted in AP buffer.
Staining for hemoglobinized red cells were achieved by incubating embryos in a solution containing O-dianisidine (Sigma) as described (34).
Analysis of skeletal development
For cartilage and bone staining (35), larvae at 5 dpf were fixed in 4% PFA for 2 h at room temperature, washed three times in PBS for 5 min, and rinsed in 50% ethanol for 10 min. Embryos were stained overnight in the staining solution (0.02% Alcian blue, 0.5% Alizarine red, 40 mM of MgCl2 in 70% EtOH). The next day, larvae were washed in clearing solutions (20% glycerol and 0.25% KOH, 50% glycerol and 0.25% KOH), rinsed in 85% glycerol/PBS and then submitted to image acquisition.
TH quantification
The quantification of TH levels was performed as previously described (36,37). Approximately 100 larvae were homogenized in 0.1 mL of lysis buffer. Samples were disrupted for 10 min on ice by sonication and centrifuged (15,871 g) for 10 min at 4°C. The supernatants were collected, and the T3 and T4 contents (ng/g fish weight) were measured using specific enzyme-linked immunosorbent assay kits (Uscnlife) following the manufacturer's instructions.
Image acquisition and statistical analysis
Images were taken with a Leica MZFLIII epifluorescence stereomicroscope equipped with a DFC480-R2 digital camera and the LAS imaging software (Leica). Confocal microscopy was performed on a Nikon C2+ confocal microscope (Nikon), equipped with a 20× objective. Images were processed using Adobe Photoshop software. All data are shown as means ± SD or percentages. Student's t-test was used for comparison of differences between groups. A p-value of <0.05 was considered statistically significant. All the analyses were conducted with GraphPad Prism v4.0 (GraphPad).
Results
hTRα variants affect zebrafish embryonic development that can be variably overcome by high T3 doses
Wild-type zebrafish embryos at the one- to two-cell stage were injected with 80 pg/embryo of each hTRα mRNA. The injection of wild-type hTRα has no evident effects on zebrafish development, whereas the hTRα mutants D211G (10), A263V (5), A382PfsX7 (3), E403X (1), and F397fs406X (4) cause relevant defects. After 6 hpf, the injected embryos were treated with the control vehicle (DMSO) or a T3 dose that was previously found to overcome the diminished T3-binding activity of mutant receptors (20 μM in the harvest water) (21). At 2 dpf, all injected embryos showed a series of morphological defects when compared with uninjected controls, which were categorized into three major groups: G1 embryos (2% of total; Supplementary Fig. S2B and B′, E and E′, H and H′, K and K′, and N and N′) developing similarly to uninjected controls (A); G2 embryos (93% of total) with several developmental alterations, including short body length, small eyes, skeletal abnormalities, hydrocephalus, and cardiac edema (Supplementary Fig. S2C and C′, F and F′, I and I′, L and L′, and O and O′); and G3 (5%) embryos with dramatic morphological alterations leading to death by 4 dpf (D and D′, G and G′, J and J′, M and M′, and P and P′). Treatment with T3 could revert the abnormal phenotypes only in embryos injected with hTRα_D211G and hTRα_A263V transcripts, as shown by the significant increase in the rate of G1 embryos (+79% and +81% vs. controls, respectively; Supplementary Fig. S2B′–D′ and E′–G′). Conversely, no significant modifications were induced by T3 in embryos injected with the truncated hTRα_A382PfsX7 (H′–J′), hTRα_E403X (Fig. 1K′–M′), and hTRα_F397fs406X (N′–P′) mRNAs.
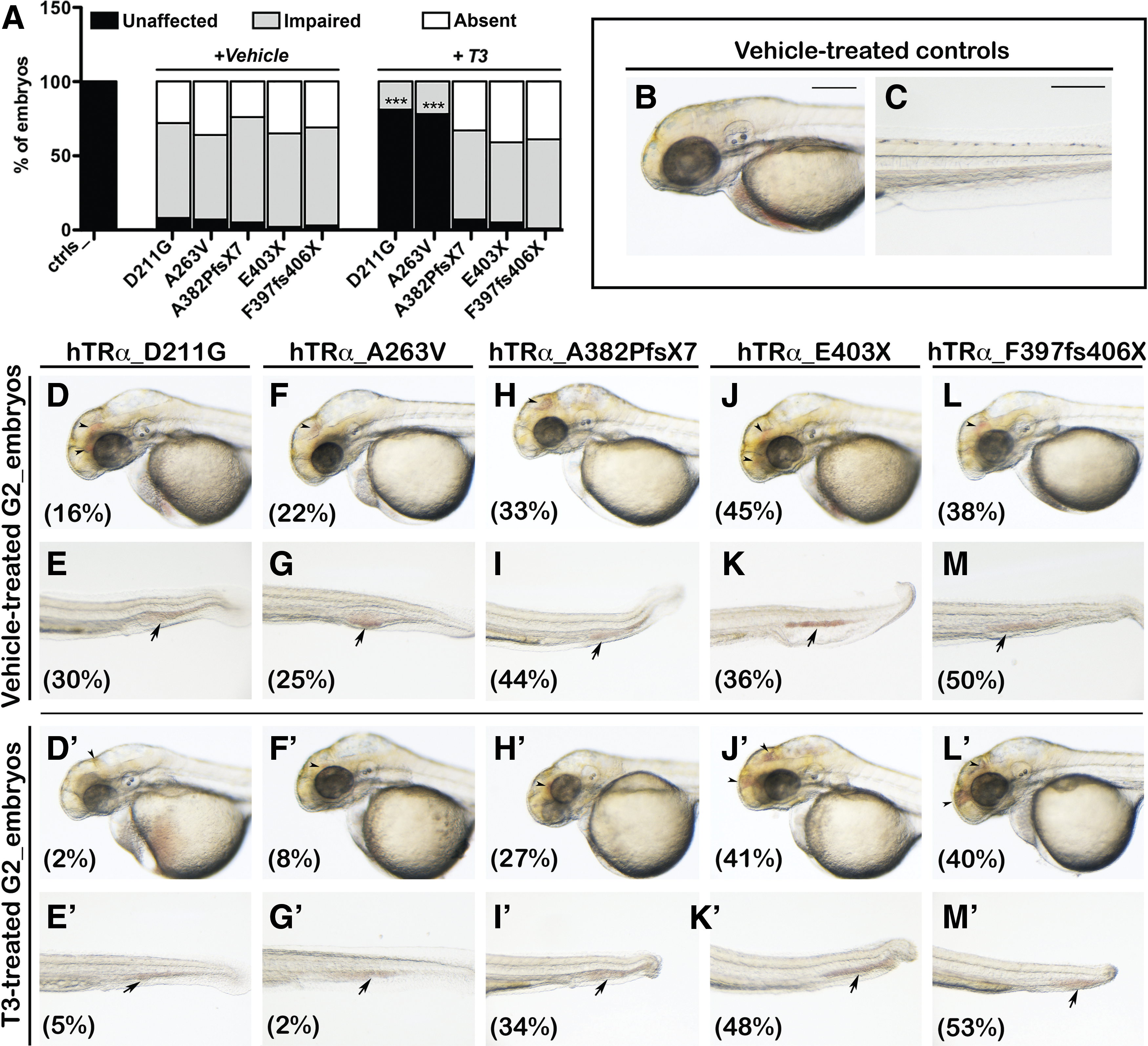
Analysis of blood circulation of human thyroid hormone receptor alpha (hTRα)-injected embryos. At 2 days post fertilization (dpf), the embryos injected with the hTRα variants displayed variable defects in blood circulation, including unaffected circulation, impaired circulation, or absent circulation. (
The mutant hTRα-injected (mut_hTRα) embryos exhibited blood circulatory defects with blood stasis in the head and the tail regions (Fig. 1). About 80–90% of vehicle-treated mut_hTRα embryos showed a severely defective blood circulation (Fig. 1A, D–M). Only the hTRα_D211G- and hTRα_A263V-injected embryos showed a significant recovery (+87% and +83% over the baseline, respectively) after T3 supplementation (Fig. 1A, D′–M′).
hTRα variants perturb development and function of zebrafish heart
The in vivo analysis of circulation defects observed in all mut_hTRα embryos prompted the investigation of their heart morphology by a molecular approach, through WISH with the cardiac-specific marker cardiac myosin light chain 2 (cmlc2; Fig. 2). At 2 dpf, the control fish had a normal S-shaped heart, with the ventricle positioned on the right of the atrium, indicating a correct D-looping process (Fig. 2A). Heart morphology was unaffected in a minority of the mut_hTRα embryos (Fig. 2B and F), but most of these embryos displayed a moderate phenotype with impaired looping (Fig. 2C), or absence of looping with a completely linear heart tube (Fig. 2D), or a reversed heart looping with the ventricle on the left of the atrium (Fig. 2E). Additionally, all mut_hTRα embryos exhibited a 20–25% reduction in heart rate compared with controls (Fig. 2G). Only the hTRα_D211G and hTRα_A263V variants positively responded to T3 supplementation, with restoration of normal cardiac development and contractility (Fig. 2F and G).
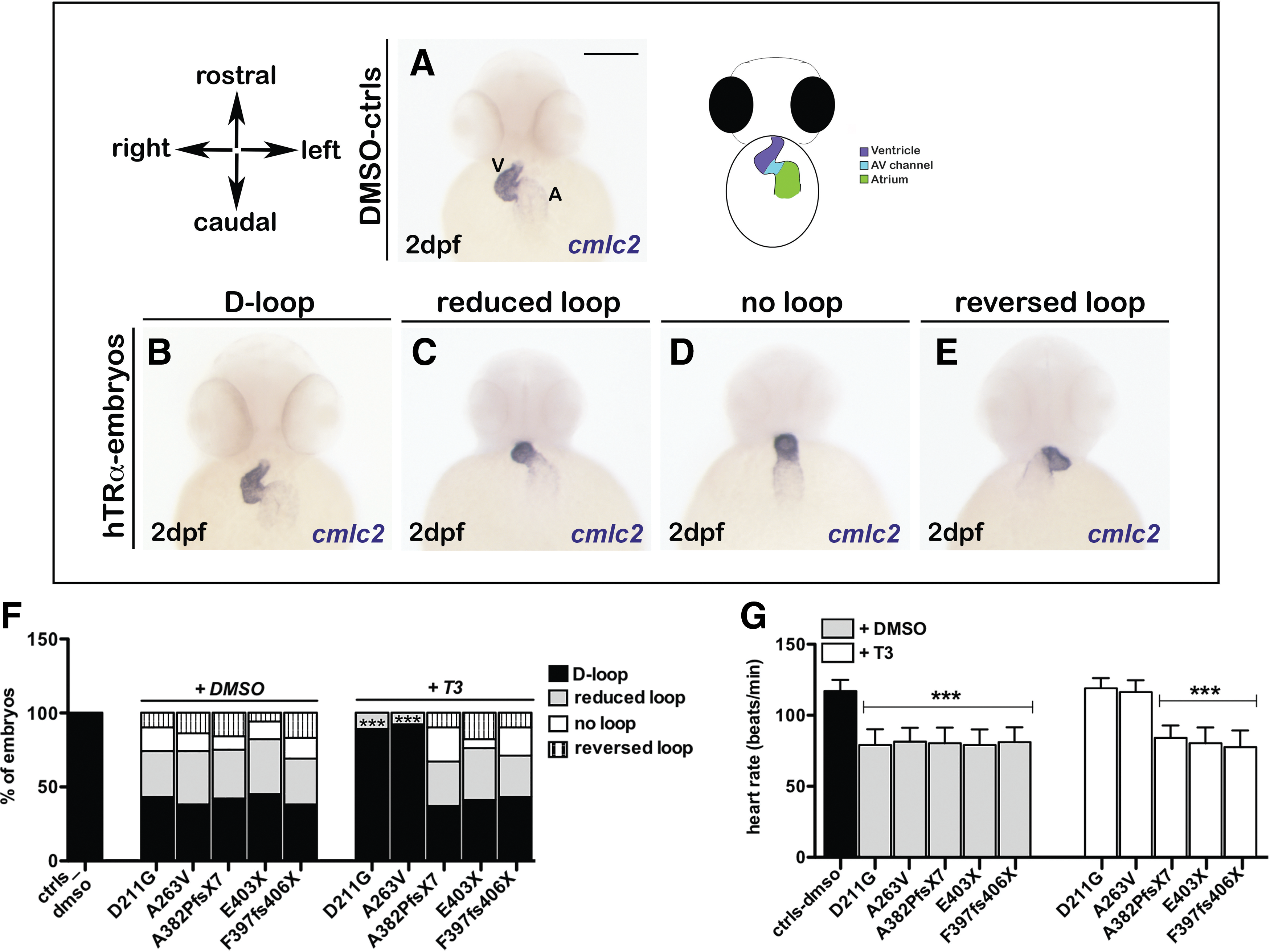
Analysis of heart looping in hTRα-injected embryos. Analysis of cmlc2 expression by whole mount in situ hybridization (WISH) was performed on controls and hTRα-injected embryos followed by treatment with vehicle or triiodothyronine (T3) at 2 dpf. Scale bar in (
hTRα variants alter vascular development and erythropoiesis of zebrafish embryos
The tg(gata1:dsRed)sd2; tg(kdrl:EGFP)S843 double transgenic line (22), in which endothelial cells are labeled in green and erythrocytes in red, was used to study vascular development. At 2 dpf, the mut_hTRα embryos had moderate to severe defects (Fig. 3). The mut_hTRα embryos with a moderate vascular phenotype were characterized by the reduction of the cardinal vein (CV) plexus and presence of all intersomitic vessels (ISVs) along the trunk and tail, but with a partially altered patterning (Fig. 3K and L vs. Fig. 3C and D). The ISVs of these injected embryos reached the dorsal longitudinal anastomotic vessel (DLAV), but they appeared thin and straight, not following the normal shape of somites (Fig. 3L). In injected embryos with a severe vascular phenotype, the posterior ISVs displayed aberrant branching, and the prematurely truncated vessels did not connect to the DLAVs (Fig. 3O and P). Notably, moderate or severe defects in ISVs were observed in mut_hTRα embryos with both normal (Fig. 3I and K) and curved tails (Fig. 3M and O), suggesting that vascular alterations were likely caused by abnormalities of angiogenesis. The rate of embryos with the illustrated defects is reported in the histogram (Fig. 3Q). Consistent with previous findings after T3 treatment, the vast majority of the D211G- and A263V-injected embryos recovered normal development of the trunk and tail vasculature (Fig. 3F and G) with well-defined and intact ISVs (Fig. 3H). Additionally, attention was focused on the development of the subintestinal vein (SIV), which delivers blood to the digestive tract of the larva (38,39). The SIV plexus, at around 3 dpf, appeared as a basket of predominantly venous angiogenic vessels, extended on the large surface of the yolk ball. The vast majority of the mut_hTRα embryos had an impaired (Supplementary Fig. S3G–I) or absent (J–L) formation of both the left and right SIV plexus compared with controls (Supplementary Fig. S3A–C). The fish injected with the missense variants D211G and A263V (M) showed significant normalization in the development of the SIV plexus (D–F) after T3 treatment. Taken together, these results indicate a central role of hTRα during angiogenic blood-vessel formation.
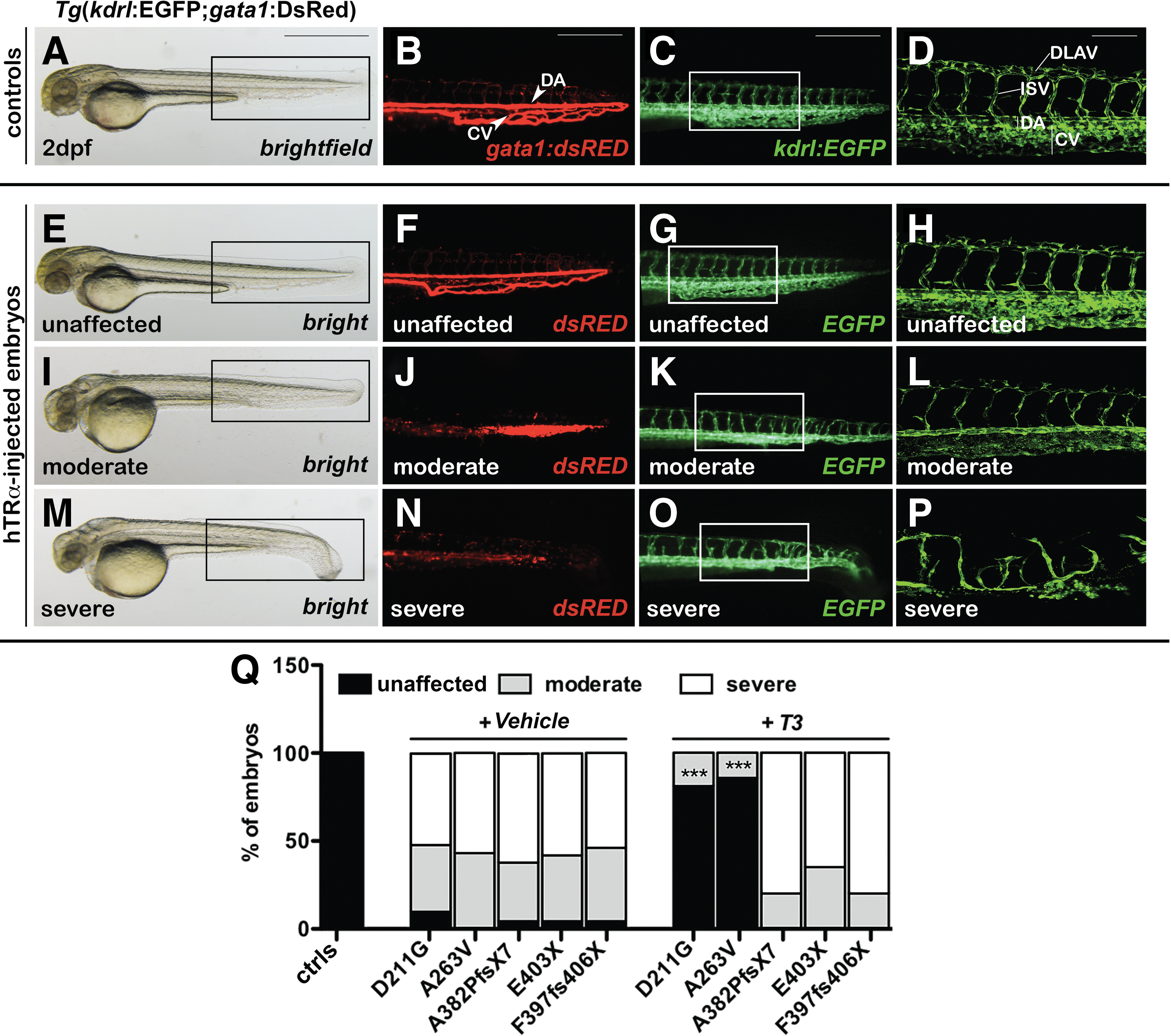
Analysis of vasculature development in hTRα-injected embryos. The double transgenic line tg(gata1:dsRed)sd2; tg(kdrl:EGFP)S843 was used to visualize the vascular architecture and the blood circulation at 2 dpf. (
Interestingly, a large number (>80%) of the vehicle-treated mut_hTRα embryos also showed blood stasis at the level of the caudal vein plexus and a diminished red staining consistent with a reduced amount of circulating erythrocytes at 2 dpf (Figs. 1 and 3J and N).
Defects of erythrocyte numbers observed in vivo were further investigated by the staining of hemoglobin with o-dianisidine (40) at 2 and 5 dpf (Fig. 4). Hypochromia was evident at 2 dpf, when >90% of the mut_hTRα embryos exhibited severe reductions in circulating erythrocytes (Fig. 4E and F, G and H), that were instead detectable in the heart (H), the sinus venosus (SV), the main axial vessels (DA and PCV), and in the CV plexus in control embryos (Fig. 4A and B). At 5 dpf, o-dianisidine clearly stained the erythrocytes localized in the jugular veins (JV), in the aortic arches (AA), and in the heart of controls (Fig. 4J), whereas the injection of the hTRα mutant transcripts resulted in a dramatic decline in mature erythrocytes (Fig. 4K, L, and M). In 15–30% of these cases, the remaining erythrocytes appeared concentrated in the head (Fig. 4L) or tail regions (Fig. 4M). Again, only the embryos injected with the missense hTRα variants showed an almost complete rescue of the hematological phenotype upon T3 treatment (Fig. 4I and N).
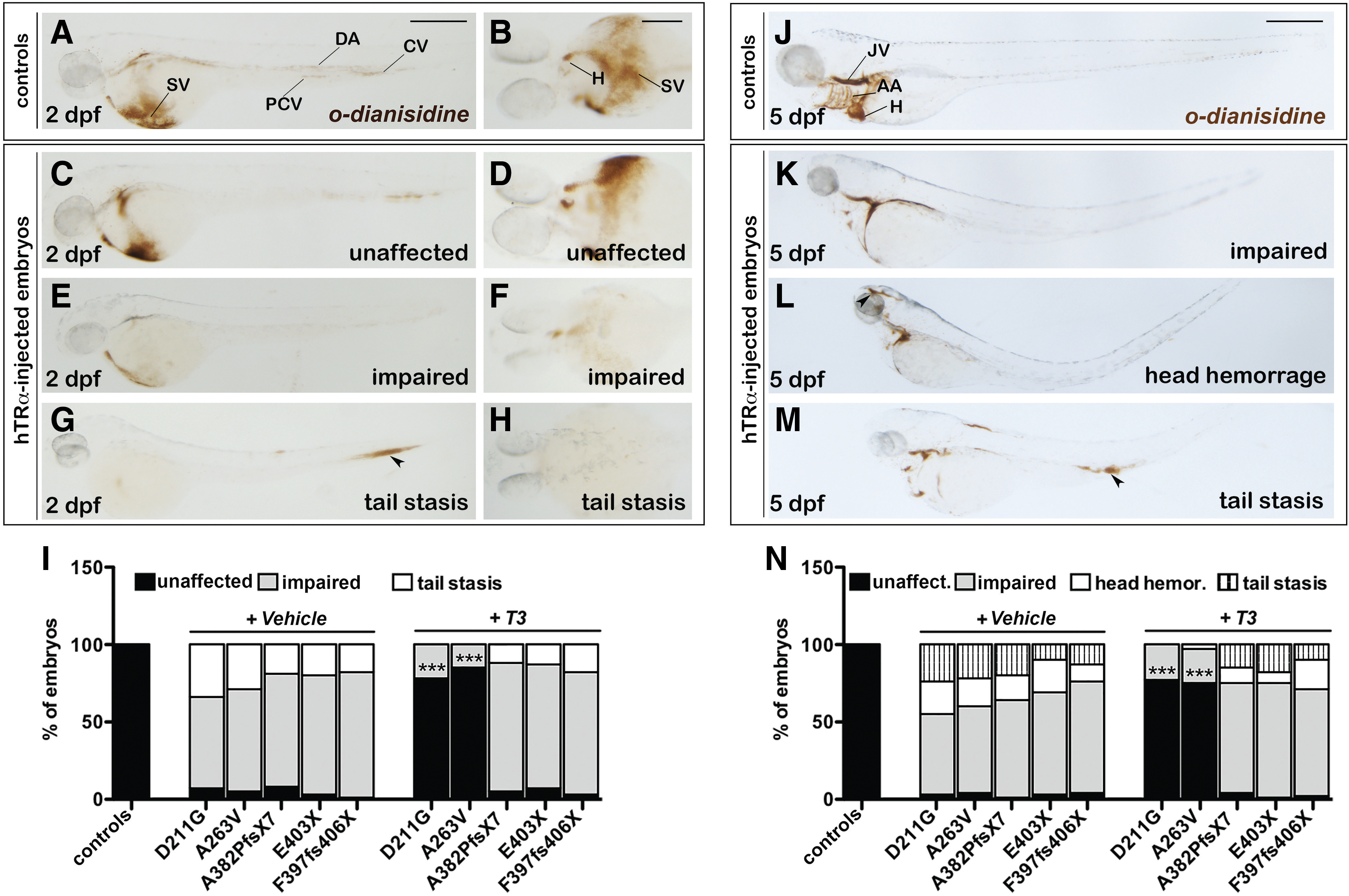
Analysis of erythropoiesis in hTRα-injected embryos. (
hTRα mutations affect motorneuron development of zebrafish embryos
It was observed that all injected embryos showed progressive motility defects, starting with reduced spontaneous movements at 1 dpf and leading to a severe or complete inability of autonomous swimming in later stages. The zebrafish spinal cord has both early-developing primary motorneurons, which arise during the first day of zebrafish development, and secondary motorneurons that develop later on. The latter axons sprout several hours after the primary motor growth cones have extended to the periphery (41,42). At 28 hpf, znp1 labeled the primary motorneurons (PMN) and their axonal projections (AP) that innervated specific parts of the adjacent myotomes in control fish (Fig. 5A and A′). In contrast, axonal pathfinding was impaired in mut_hTRα embryos with a moderate to severe disorganization of motorneurons that appear misorientated, or truncated (Fig. 5B–D), but this was accompanied by a normal development of the muscle fibers (Fig. 5B′–D′). At 52 hpf, by performing IHC with a specific antibody for secondary motorneurons (zn-5), it was observed that secondary motorneuron axons were also affected in mut_hTRα embryos (Fig. 5E–H). In >85% of these embryos, axons of the secondary motorneurons (SMN) stalled at the distal point of the common path, failing to reach the horizontal myoseptum (HMS; Fig. 5F) or were totally absent (Fig. 5G and H). Consistent with previous data, T3 treatment significantly improved motorneuron development only in embryos injected with the missense hTRα_D211G and _A263V transcripts (I and J).
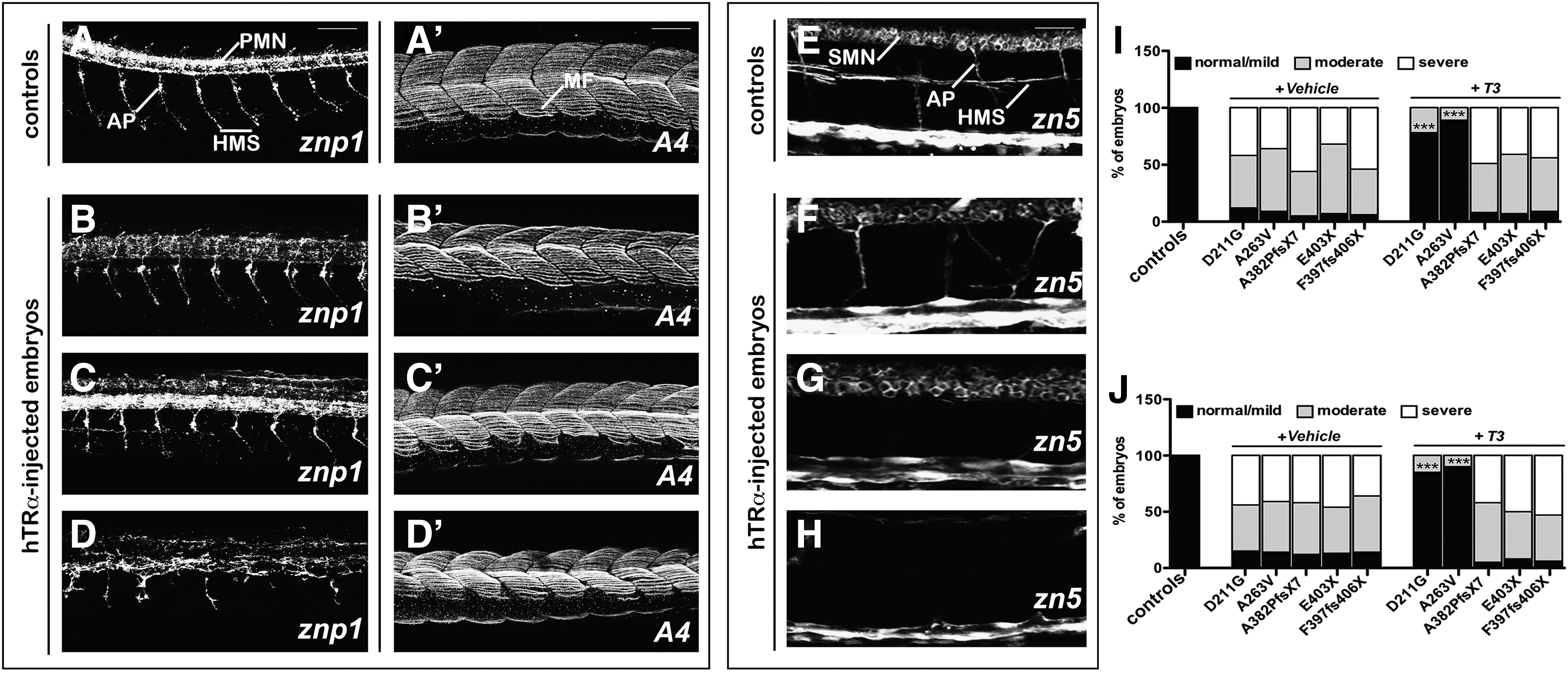
Analysis of development of motorneurons in hTRα-injected embryos. Left: Staining of controls of the znp1 monoclonal antibody at 28 hours post fertilization (hpf). Zpn1 marks the primary motorneurons (PMN), visible at the level of the spinal cord, and their axonal projections (AP) that extend ventrally and reach the horizontal myoseptum (HMS) (
hTRα variants perturb the development of cartilage and bones of zebrafish larvae
In mut_hTRα embryos, most of the pharyngeal and neurocranium cartilage elements were malformed (Fig. 6G–L vs. 6A–C). Indeed, the vast majority of the vehicle-treated embryos presented with moderate to severe phenotypes (Fig. 6M). In mut_hTRα embryos with a moderate phenotype, the first mandibular arch (Meckel's cartilage) appeared slightly malformed, whereas the second arch (ceratohyal cartilage) was unevenly formed (Fig. 6G–H), and the five ceratobranchial arches were largely lacking (Fig. 6H). In all severely affected embryos, all cartilaginous components were completely absent (Fig. 6J–L). Severe morphological phenotypes were also observed in bone mineralization. Under basal conditions, most of the mut_hTRα embryos showed an impaired or absent development of the brachiostegal rays and operculum (Fig. 6G and H and J and K) in comparison with controls (Fig. 6A and B). In contrast, a strong reduction or absence of mineralization of the cleithrum and notochord was seen only in the mut_hTRα embryos with a severe phenotype (Fig. 6J and L). Consistent with previous results, T3 treatment effectively reverted the pathological skeletal phenotype only in the embryos injected with the hTRα_D211G and hTRα_A263V transcripts (Fig. 6D–F and M).
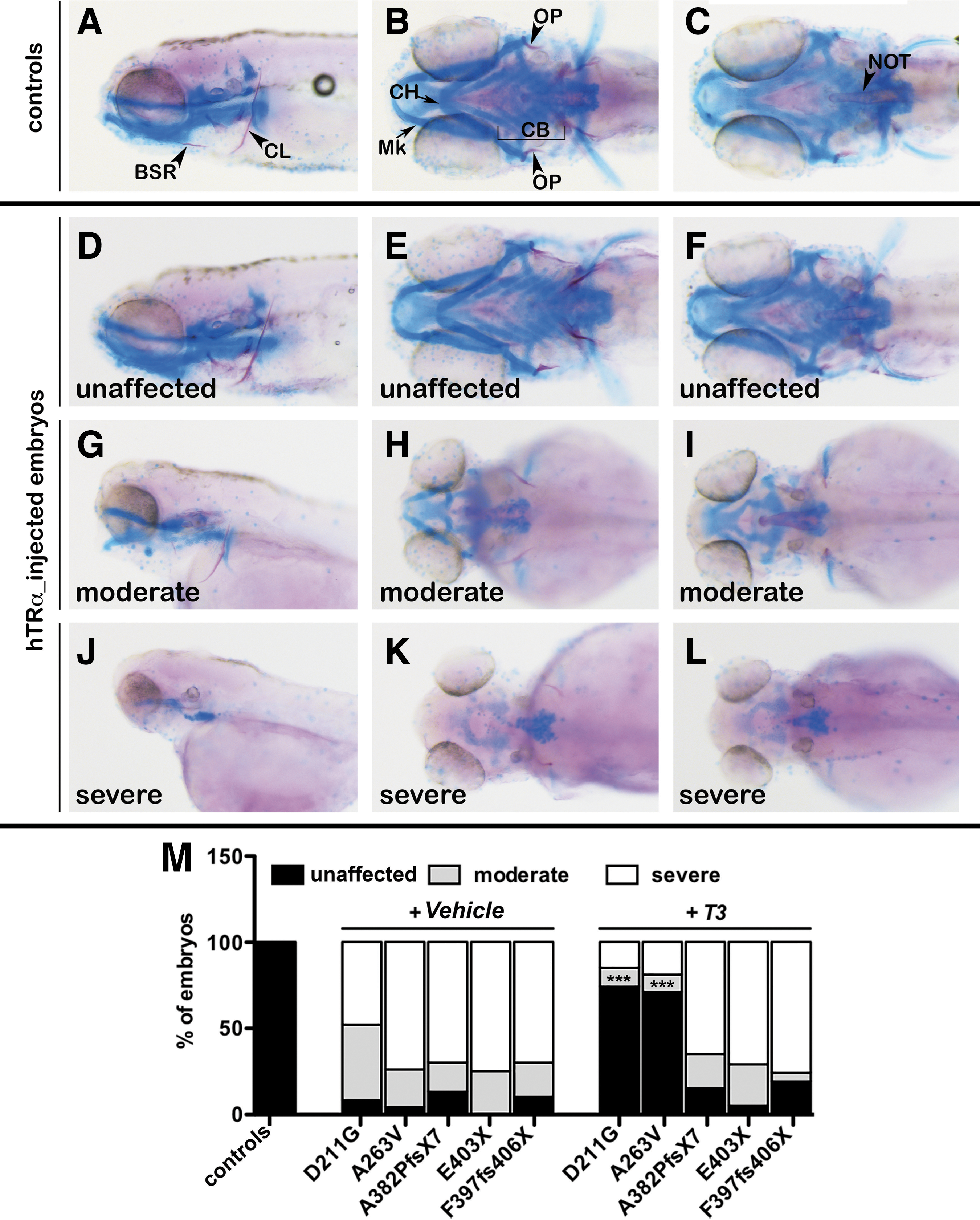
Analysis of cartilage and bone development in hTRα-injected larvae. At 5 dpf, all embryos were stained with Alician Blue (in blue) and Alizarine (in red) to visualize the formation of the head cartilage and the mineralization of bones, respectively. (
Injection of the hTRα variants recapitulates the biochemical features of RTHα
The TH sensitivity at 5 dpf—the stage in which the zebrafish gland produces T4, and the development of the HPT axis is complete and responds to endogenous TH levels (43)—was explored. Under basal conditions, the mut_hTRα larvae presented normal levels of the hypothalamic trh (Fig. 7D, G, J, M, and P) and pituitary tshba (Fig. 7E, H, K, N, and Q), but displayed a low number of T4-producting follicles (Fig. 7F, I, L, O, and R) when compared with controls (Fig. 7A–C). The administration of 20 μM of T3 led to the suppression of trh and tshba transcripts (Fig. 7D′, G′, J′, M′, and P′) together with an expected further reduction of T4-positive follicles (Fig. 7F′, I′, L′, O′, and R′). The measurement of the T4 content gave consistent results (Fig. 7S). Conversely, T3 levels were significantly increased in hTRα-injected larvae in both basal (p < 0.01) and T3 conditions (p < 0.001). Importantly, the changes observed in TH levels of mut_hTRα larvae after T3 treatment (p < 0.01 vs. vehicle) together with the suppression of trh, tshba, and T4 reflected the conservation of the TH sensitivity of the HPT axis.
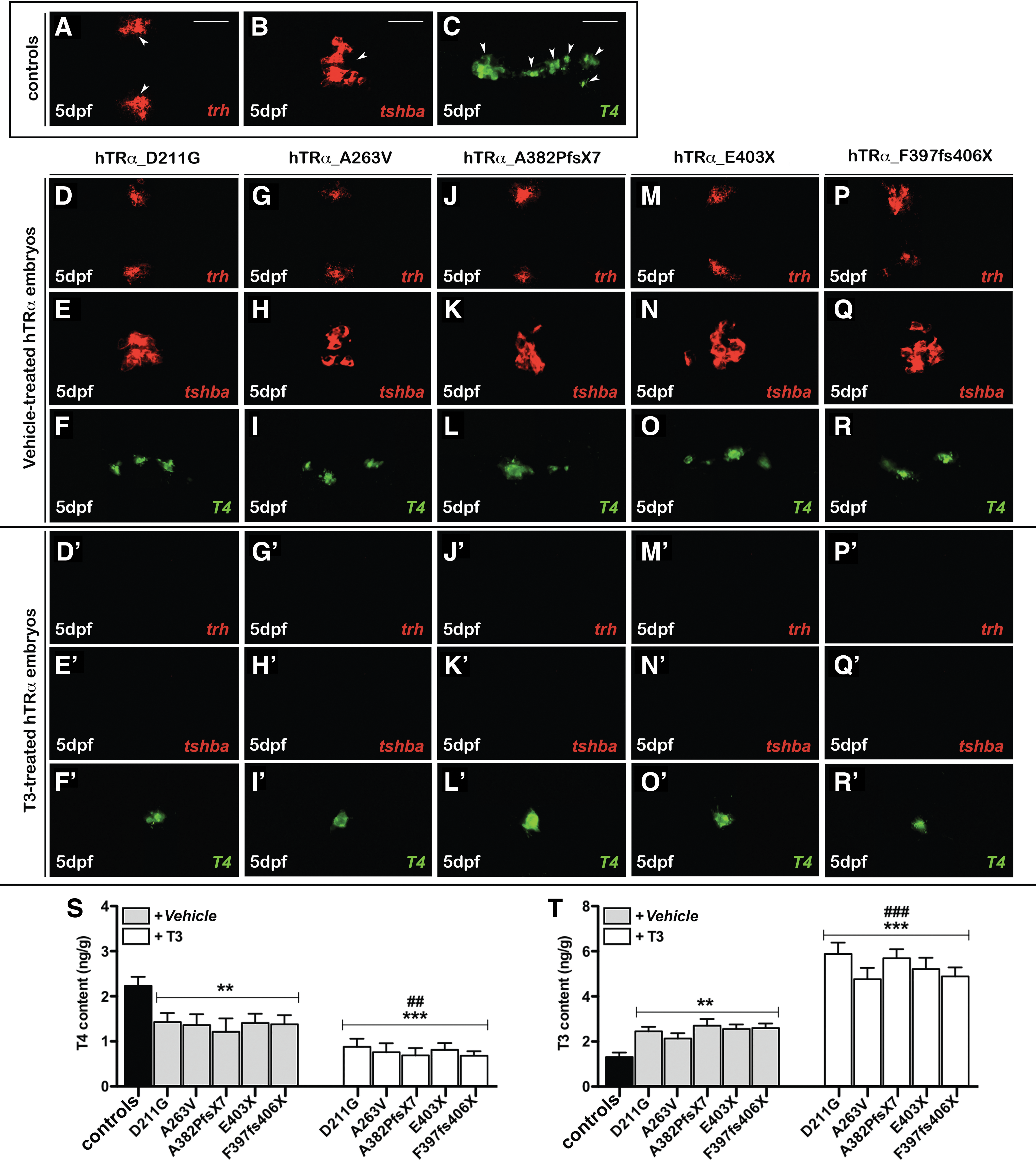
Evaluation of hypothalamic–pituitary–thryroid (HPT) axis function and thyroid hormone levels of hTRα-injected larvae. WISH of trh and tshba, and immunohistochemistry (IHC) of thyroxine-producing follicles followed by confocal microscopy in controls acquired in ventral view, head to the left (
Therefore, the ability of human D211G and E403X variants on the activity of zebrafish receptors was tested using a published T3-dependent activation assay (1) (Supplementary Fig. S4). As expected, the hTRα_D211G and _E403X plasmids did not activate the TH-responsive gene, either in the absence or in the presence of 10 nM of T3 (A). Furthermore, when co-expressed, both variants strongly inhibited the T3 transcriptional activation of the wild-type human and zebrafish TRs in a dominant negative manner (Supplementary Fig. S4B). Interestingly, both D211G (–30%) and E403X (–27%) exhibited a less pronounced interference on wild-type activity when co-transfected with the zTRβ2 plasmid (Supplementary Fig. S4B). Since TRβ2 is one of the major contributors in the regulation of the HPT axis, particularly on TSHβ expression (44), these data may explain the conserved response of tshβ and trh transcripts observed in mut_hTRα embryos in vivo.
The high T3/T4 ratio observed in the mut_hTRα embryos may be explained by an altered TH metabolism, as previously described (21). As reported in Supplementary Fig. S5, at 2 dpf, the expression of pituitary dio2 was increased in both vehicle and T3-treated mut_hTRα embryos (D and D′, G and G′, J and J′, M and M′, and P and P′ vs. control A and A′). At 1 dpf, all vehicle-treated mut_hTRα embryos presented a reduced expression of both dio3a in the brain region (E, H, K, N, and Q) and dio3b in the pronephros (F, I, L, O, and R). In responses to T3 treatment, only the embryos injected with hTRα_D211G and hTRα_A263V transcripts recovered a normal expression of dio3a and dio3b (E′ and F′ and H′ and I′). These results have been also confirmed by qRT-PCR. Remarkably, changes in TH metabolism observed at 2 dpf (for dio2) and at 1 dpf (for dio3a and dio3b) persisted in the mut_hTRα larvae at 5 dpf (S–V).
Discussion
The present work took advantage of the previously described interchangeability between the human and zebrafish TRα proteins (21) to generate a simplified “tailor-made” in vivo model to study the pathological consequences of human THRA variants at the various tissue levels. The phenotypic consequences induced by several human THRA variants were analyzed, including two missense (D211G (10), A263V (5)) and three truncated (A382PfsX7 (3), E403X (1), and F397fs406X (2)) hTRα forms, forcing their expression by direct mRNA microinjection in zebrafish one- to two-cell embryos, as recently done with the human mutant transcripts found in patients with dynaminopathies (45). The embryos injected with hTRα mutants underwent profound morphological, neurological, cardiovascular, hematological, skeletal, and biochemical alterations reminiscent of RTHα in humans. An efficient rescue of the disease phenotypes was always seen upon the addition of a high T3 concentration in the harvesting water only in embryos injected with the two missense variants, which are partially resistant to T3, but not with the completely refractory truncation mutations (1 –10,21). Therefore, these findings support the view that the observed fish phenotypes are specifically caused by the DN activity exerted by hTRα mutants on zTRs, and that this may constitute a simplified model to study the functional consequences of hTRα mutants in vivo.
Interestingly, this model recapitulates the biochemical abnormalities described in RTHα cases, confirming that the disproportionately raised T3/T4 ratio is the consequence of the combination of reduced levels of dio3, whose expression is known to be regulated by TRα1 (46), and raised dio2 expression and activity. Both these modifications represent suitable responses to the impaired TH action at the tissue level (21). Consistent with data reported in patients with RTHα, and in contrast to the high TSH levels found in several murine models (12 –20), the expression of the different hTRα variants caused modifications of TH levels in presence of normal expression of trh, tshba, and tg. Furthermore, the sensitivity of the HPT axis of the injected embryos was well conserved, since they properly responded to exogenous administration of T3, thus allowing the presented model to recapitulate human RTHα also at the biochemical level. The lack of an increased resistance to T3 in the hypothalamus and pituitary despite the unrestricted expression of hTRα may be due to intrinsic properties of the endogenous TRα and TRβ, as previously described in knock-in mice (47). Consistently, the activity of the zTRβ2 is only partially disturbed by both missense and truncated hTRα variants also in vitro, suggesting a minor interaction between these two isoforms. Nevertheless, only the TRβs (as homodimers or heterodimers with RXR) appear able to affect the transcription of the specific repertoire of target genes involved in TH feedback mechanism effectively.
Regarding the morphological and phenotypical consequences of the injected embryos, this study shows that the expression of all hTRα mutants resulted in variable defective embryonic development, confirming that the human mutant transcripts are able to interact with the functional zebrafish receptor in vivo (21). The hTRα-injected embryos had hydrocephalus and cardiac edema associated with brain hemorrhages and blood stasis likely as a result of defective development or function of the cardiovascular system. The expression analysis of the specific cardiac marker cmlc2 by WISH revealed that the majority of the injected embryos at 2 dpf presented several defects in heart looping, as a likely consequence of an altered heart tube bending associated with low heart rate at 2 dpf. These “altered looping” and bradycardia phenotypes indicate that the expression of genes involved in the differentiation of cardiac precursors, in the lateralization of the cardiac tube, or in the pacemaker functions (48 –51) are also T3-TRα dependent in zebrafish. It is well documented that T3 plays a central role in the control of heart development and function, and that TRα1 is expressed in the myocardium, the atria, and the proximal conduction system (PVCS including the sino-atrial node and the atrio-ventricular node) in mice (52). Homozygous TRα1–/– mice present an average heart rate that is 20% lower than that of control animals, and prolonged QRS and QTend duration, suggesting that TRα1 tightly controls cardiac pacemaking and ventricular repolarization (20). Moreover, mice heterozygous for a mutant TRα1+/m exhibit bradycardia, reduced contractility, and delayed relaxation time of isolated cardiomyocytes (53).
Most of the injected embryos presented vascular defects, including a reduction of the CV plexus, frequently in combination with misguided branches and premature ending of the intersomitic vessels at 2 dpf. At 3 dpf, the embryos also exhibited a reduced or absent development of the subintestinal vessels, a complex network of vessels that delivers blood to the developing digestive tract and which is a good indicator of angiogenesis processes (39,54). The proangiogenic effect of TH has been studied in rats, which displayed substantial coronary angiogenesis concomitant with the upregulation of the basic fibroblast growth factor (Fgf2) upon T4 treatment (55). Furthermore, it has been reported that TH have a relevant impact on brain vasculature development (56). Hypothyroid rats have decreased brain angiogenesis with a reduced complexity and density of microvessels, in parallel with a reduced expression of Fgf2 and vascular endothelial growth factor A; all these functions could be rescued by T3 treatment (56). Interestingly, physiologic concentrations of TH stimulate the proliferation of brain-derived endothelial cells in vitro, accompanied by the downregulation of the apoptosis-related genes Bcl2 and Bad (56). Therefore, the present results confirm a central role for TH in angiogenesis and strongly indicate that these angiogenic effects are mediated by TRα.
The injection of hTRα mutants in zebrafish embryos produced marked blood stasis at the level of the CV plexus and a low amount of circulating erythrocytes at 2 dpf. Analysis of the hemoglobin content by o-dianisidine staining up to 5 dpf confirmed that anemia was evident in >90% of embryos exhibiting a dramatic reduction of circulating erythrocytes. Anemia in RTHα patients correlates with documented abnormal erythropoiesis and a reduced hematocrit in TRα–/– or TRαPV/+ mutant mice (57,58), although the mechanisms of defective proliferation or differentiation of erythroid progenitors remain to be elucidated. In vitro and in vivo studies demonstrated that the T3-dependent TRα induction can affect the signal transduction pathways implicated in proliferation of hematopoietic progenitors for the erythroid lineage (59 –62).
It was also observed that the expression of the hTRα variants affects motorneuron development, resulting in reduced spontaneous movements of embryos at 1 dpf and in an almost complete swimming inability of the developing larvae at 3 and 5 dpf. Most of the hTRα-injected embryos presented with moderate to severe alterations of primary and secondary motorneurons, but in the presence of normal development of the muscle fibers, suggesting that hTRα is implicated in the establishment of muscle innervation. Consistently, RTHα patients present a reduced muscle tone with normal power but with impairment in gross and fine motor coordination, resulting in slow and clumsy movements (11). Moreover, neonatal hypothyroidism induced in rats by propylthiouracil treatment during gestation was associated with a reduced innervation of the muscle fibers and delayed movements, whereas these processes were accelerated in hyperthyroid conditions (63). Interestingly, locomotor dysfunctions have also been described in TRα1-R384C mice, which suffer from reduced grip strength, poor limb coordination, and an abnormal gait, findings that correlate with an aberrant development of GABAergic interneurons (64).
Finally, the profound alterations in cartilage and bone development could represent the zebrafish correlate of the delayed bone development and skeletal dysplasia described in RTHα patients (11) and in genetically modified mice (16 –18). The skeleton is a target tissue that is highly sensitive to TH, and T3 exerts anabolic actions during growth and catabolic effects in adult bone (65). Both hyperthyroid and hypothyroid conditions are associated with secondary osteoporosis and osteoarthritis or delayed bone maturation and skeletal dysplasia, respectively (65). Moreover, the similar skeletal phenotypes of TRα–/– and TRα–/– TRβ–/– mice suggested that TRβ is not essential for the normal postnatal maturation of bone, and that TRβ cannot substitute for TRα in this process (66). The present results thus further confirm the relevant role exerted by TRα during bone development in vertebrates.
Conclusions
In conclusion, the injection of human THRA mutant transcripts in zebrafish (one- to two-cell stage) embryos can cause tissue-specific interference in TH action that recapitulates the biochemical and clinical features of RTHα patients. This study reveals for the first time the involvement of TRα in the angiogenic processes.
Since high doses of T3 can overcome the binding defect and negative action of human TRα mutants, this “tailor-made” zebrafish model can be useful to shed light on the molecular mechanisms and functional consequences of the newly discovered human THRA variants, and to test new compounds that may be designed to overcome the specific deleterious effects of TRα mutations, perhaps avoiding the thyrotoxic effects on TRβ-expressing tissues.
Footnotes
Acknowledgments
The authors wish to thank Prof. Krishna Chatterjee and Dr. Maura Agostini (Cambridge, United Kingdom), and Prof. Robin Peeters (Rotterdam, NL) for the supply of the mutant hTRα plasmids.
This research was supported by funding from the Italian Ministry of Health, Rome, Italy (grant RF-2010-2309484), and the Ricerca Corrente of IRCCS Istituto Auxologico Italiano, Milan, Italy (Zebratir grant: 05C102_2011).
Author Disclosure Statement
The authors have nothing to disclose.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.