
Abstract
Background:
High mobility group box-1 (HMGB1), a non-histone protein, plays an important role in autoimmune diseases. However, the significance of HMGB1 in the pathogenesis of autoimmune thyroiditis has not been reported. The purpose of this study was to explore whether HMGB1 participates in the pathogenesis of autoimmune thyroiditis, and whether glycyrrhizin (GL), a direct inhibitor of HMGB1, attenuates the severity of thyroid inflammatory infiltration in a murine model of autoimmune thyroiditis.
Methods:
A total of 80 male NOD.H-2h4 mice were randomly divided into a control or iodine supplement (NaI) group at four weeks of age, and the control group was fed with regular water, whereas the NaI group was supplied with 0.005% sodium iodine water. Another 24 male NOD.H-2h4 mice were also randomized into three groups (eight mice per group) as follows: control, NaI, and GL treatment after iodine supplementation (NaI + GL). The NOD.H-2h4 mice were fed with 0.005% sodium iodide water for eight weeks to enhance autoimmune thyroiditis. After iodine treatment, the mice received intraperitoneal injections of GL for four weeks. The severity of lymphocytic infiltration in the thyroid gland was measured by histopathological studies. The serum levels of HMGB1, tumor necrosis factor alpha, interleukin (IL)-6, IL-1β, and thyroglobulin antibody titers were measured using an enzyme-linked immunosorbent assay. HMGB1 expression was measured by immunohistochemical staining and real-time polymerase chain reaction. TLR2, HMGB1, MyD88, and nuclear transcription factor κB were measured by Western blot.
Results:
The mRNA expression of HMGB1 was significantly higher at 8 and 16 weeks in the NaI group than it was in the control group. Serum levels of thyroglobulin antibodies, HMGB1, tumor necrosis factor alpha, IL-6, and IL-1β were significantly increased in the NaI group, but they were dramatically attenuated with GL injection. The prevalence of thyroiditis and the infiltration of lymphocytes were significantly decreased in the NaI + GL group. GL administration also significantly reduced the protein expression of TLR2, MyD88, HMGB1 and nuclear transcription factor κB in the thyroid gland and attenuated the severity of thyroiditis.
Conclusion:
HMGB1 may play a crucial role in autoimmune thyroiditis by causing inflammatory infiltration, thus increasing the severity of autoimmune thyroiditis. GL effectively attenuated thyroiditis in the iodine-induced NOD.H-2h4 mice via a molecular mechanism related to the inhibition of TLR2-HMGB1 signaling.
Introduction
A
High mobility group (HMG) proteins are non-histone DNA binding proteins that are common in eukaryotic cells and exert a variety of biological functions (4). HMGB1, a member of the HMG family, is mainly located in the nucleus and interacts with DNA, modulates chromosomal architecture, and can be transferred outside of the cell in some circumstances, such as apoptosis and necrosis. When translocated outside of the cell, HMGB1 can act as a damage-associated molecular pattern (DAMP) or alarmin (5). Extracellular HMGB1 is released by active secretion from innate immune cells or derived passively from injured cells (6). Recent research has shown that extracellular HMGB1 may play a role in the pathogenesis of inflammatory disease. Extracellular HMGB1 stimulates the secretion of cytokines and pro-inflammatory mediators by macrophages (7). In return, cytokines promote HMGB1 secretion, which forms a positive feedback loop and plays an important role in maintaining the inflammatory response. HMGB1 can also bind with pattern-recognition receptors, such as Toll-like receptors (TLR), to activate TLR signaling pathways, inducing the production of cytokines and the proliferation of immune cell (8). The role of HMGB1 may be dose dependent. That is, a medium concentration of HMGB1 may be beneficial to the host response by limiting inflammation or tissue damage and promoting injury repair and tissue regeneration (9). However, a high concentration of HMGB1 may lead to an uncontrolled inflammatory response, make tissue injury worse, and result in extensive tissue damage and organ failure (10).
Increasing evidence suggests that HMGB1 plays a crucial role in the development of many autoimmune diseases. Increased expression of HMGB1 has been found in the serum and local lesions of patients with autoimmune diseases, such as systemic lupus erythematosus (SLE) (11) and rheumatoid arthritis (RA) (12). HMGB1 may also play an important role in the maturation of dendritic cells and can be released during cell apoptosis or activation. Previous studies in SLE patients and mouse models of SLE suggested that increasing levels of serum HMGB1 may be an indicator of abnormal immune cells in SLE, and that high levels of serum HMGB1 may induce systemic inflammatory reactions and immune responses (13,14). New research has found that HMGB1 is upregulated in the serum and peripheral blood mononuclear cells of SLE patients compared with healthy controls. Furthermore, in SLE patients, the concentration of HMGB1 was positively correlated with the activity index and negatively correlated with C3 and C4. These studies provide further proof that HMGB1 is involved in the inflammatory process in SLE (15).
HMGB1 may be activated through the receptor for advanced glycation end products (RAGE) or through TLR2, TLR4, and TLR9, thus leading to the activation of nuclear transcription factor κB (NF-κB), which elevates the production of pro-inflammatory cytokines and results in an inflammatory response. Glycyrrhizin (GL) is a traditional medicine extracted from licorice root, which has been used in Japan and China for several decades. GL has been reported to have anti-inflammatory activity and to inhibit HMGB1 by binding to both HMG boxes (16). A recent study has reported that GL protects against tissue damage in an ischemia–reperfusion animal model (17). However, to the best of the authors' knowledge, there are no reports regarding the protective role of GL via the HMGB1 pathway in AIT. Therefore, the aim of this study was to investigate the effect of GL, a known inhibitor of HMGB1, in a murine model of autoimmune thyroiditis.
Methods
Mice and groups
NOD.H-2h4 mice were purchased from Jackson Laboratory (Bar Harbor, ME). Animals were maintained in the animal house of China Medical University under specific pathogen-controlled temperature, humidity, and light/dark cycles, along with regular disinfection. All animal experimental procedures were performed in accordance with the rules approved by Animal Ethics Committee of China Medical University.
Experiment 1
A total of 80 NOD.H-2h4 mice were randomized into two groups at four weeks of age: a NaI supplement group and a control group. Week 0 was used as an initial time point in the experiment. The NaI group was given drinking water containing 0.005% NaI for 2, 4, 8, and 16 weeks. The control group was given regular water for the same time period, without being fed sodium iodide. Mice from the NaI and control groups were anesthetized with 10% ethyl carbamate (Sigma–Aldrich, St. Louis, MO) and then killed at 2, 4, 8, and 16 weeks after the initiation of the experiment. Thyroid tissues were removed, washed with cold saline, dried on a pad of filter paper, and weighed on an electronic balance (BS210S; Sartorius, Göttingen, Germany).
Experiment 2 (GL treatment)
NOD.H-2h4 mice were fed in the animal house until four weeks of age. A total of 24 male mice were then randomly separated into three different groups (eight per group). Mice in the control group were given sterile water without supplement. Mice in the NaI group were given 0.005% NaI in the drinking water. Mice in the NaI + GL group were treated with 50 mg/kg of GL (Sigma–Aldrich) once a day for four weeks, administered via intraperitoneal injection after eight weeks of iodine supplementation. Thyroid tissues were removed surgically under anesthesia, washed with cold saline, blotted on filter paper, and weighed using an electronic balance (BS210S; Sartorius). The thyroid gland tissues were then stored at −80°C until use.
Assessment of autoimmune thyroiditis
One thyroid lobe from each mouse at each experimental time point was immediately fixed in 4% paraformaldehyde for at least 24 h, embedded in paraffin, and then sliced into 5-μm-thick sections. Sections were prepared and stained with hematoxylin and eosin (H&E). Histological changes in thyroid tissue were evaluated under light microscopy (BX51/BX52; Olympus, Tokyo, Japan). The extent of lymphocytic infiltration was assessed, as previously described (18). Briefly, H&E-stained thyroid sections were graded using the following scale, according to the area of lymphocytic infiltration: 0, normal; 1+, 1–10%; 2+, 10–30%; 3+, 30–50%; and 4+, >50%. The thyroiditis scores shown are the means of at least three non-continuous sections of thyroid gland.
Measurement of serum TgAb by enzyme-linked immunosorbent assay
Frozen thyroid tissues were homogenized and centrifuged. The supernatant was then collected and murine Tg (mTg) obtained, as previously described by Imaizumi et al. (19). Blood was collected from the orbital venous system, stored at room temperature for at least 2 h, separated by centrifugation at 1500 g for 20 min, and stored at −80°C until use. Mouse serum was diluted 1:100 and then assessed using 96-well plates, which had been coated with 1 μg/mL of mTg, and stored overnight at 4°C. The plates were then washed and blocked with 1% bovine serum albumin (BSA). Peroxidase-labeled rabbit anti-mouse immunoglobulin G (1:250 dilution; Sigma–Aldrich) and tetramethylbenzidine (T0440; Sigma–Aldrich) were used as the secondary antibody. Absorbance was measured at 450 nm using in a microtiter plate reader (Model 680; Bio-Rad Laboratories, Inc., Hercules, CA).
Determination of serum levels of tumor necrosis factor alpha, interleukin-6, interleukin-1β, and HMGB1
The serum levels of tumor necrosis factor alpha (TNF-α), interleukin (IL)-6, IL-1β, and HMGB1 were measured via a quantitative enzyme immunoassay technique, using enzyme-linked immunosorbent assay kits (TNF-α, IL-6, IL-1β: R&D Systems, Minneapolis, MN; HMGB1: Shino-Test Corp, Sagamihara, Japan), according to the manufacturers' instructions. Absorbance was measured at a wavelength of 450 nm, using a microplate reader, to analyze the intensity of color development in each well.
Immunohistochemistry analysis
Thyroids from mice were embedded in paraffin and sectioned coronally into 5 μm sections using a microtome. Sections were dewaxed, rehydrated, and treated for endogenous peroxidase using 3% methanol-hydrogen peroxide for 10 min. All sections were incubated with the primary antibody (HMGB1, 1:250; Abcam, Cambridge, MA) at 4°C overnight and were then incubated for 40 min at room temperature. The tissue sections were incubated in biotin-conjugated secondary antibodies (Zhongshan Biotechnology, Beijing, China) for 10 min and then with a streptavidin-peroxidase complex (Zhongshan Biotechnology) for 10 min. Sections were treated with a solution of 3,3′-diaminobenzidine (DAB; Zhongshan Biotechnology) for 1–3 min, depending upon the staining of the DAB reaction product observed under light microscopy. Finally, the sections were counterstained with hematoxylin, dehydrated, rinsed, and mounted in neutral gum (China National Medicines, Shanghai, China). All thyroid sections were analyzed at 400 × . The integral optical density (IOD) values, which indicate protein expression levels, were measured using Image-Pro Plus 5.0 software (Media Cybernetics, Silver Spring, MD).
Western blot analysis
Western blot was used to detect proteins in samples from five animals from each group. After dissection, the thyroid lobes were quickly frozen in liquid nitrogen and stored at −80°C. Frozen lobes were homogenized in phosphate-buffered saline (pH 7.4) supplemented with 0.05% Triton X-100 and a protease inhibitor cocktail (Sigma–Aldrich). The protein concentration was determined by the Bradford method using BSA as the standard. SDS-PAGE was performed using 30% acrylamide (Sigma–Aldrich). After electrophoresis, gels were equilibrated for 20 min in transfer buffer (25 mM of Tris, 190 mM of glycine, and 20% methanol). Proteins were transferred onto polyvinylidene difluoride membranes (0.5 h, 30 V), which were then incubated with a blocking solution consisting of 5% dried skim milk in 100 mM of Tris (pH 7.5) with 140 mM of NaCl and 0.01% Tween 20 for a minimum of 1.5 h. The blots were then incubated overnight at 4°C, either with a mouse monoclonal anti-HMGB1 antibody (1:800; Abcam), a polyclonal rabbit anti-TLR2 antibody (1:800; Abcam), a polyclonal rabbit anti-MyD88 antibody (1:800; Abcam), a mouse monoclonal anti-NF-κB antibody (1:800; Abcam), or a polyclonal rabbit anti-β-actin (1:1000; Abcam) on the same membrane. The blots were washed three times with the blocking solution and incubated with diluted horseradish peroxidase-conjugated secondary antibodies (1:1000; Bio-Rad) for 1.0 h at room temperature. Blots were washed extensively and developed using an enhanced chemiluminescence kit (Amersham Pharmacia Biotech, Piscataway, NJ). Western immunoblot bands were quantified by means of a Bio-Rad calibrated densitometer (GS-800) using the vendor's software (Bio-Rad Laboratories, Inc.), and β-actin was used as an internal control for all analyses.
Quantitative real-time polymerase chain reaction analysis
The mRNA expression of HMGB1 was measured using TaqMan real-time polymerase chain reaction (PCR). Total RNA was extracted from thyroid gland tissue from each experimental time point using TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Waltham, MA) following the standard protocol instructions. The Nano Drop 2000C (Thermo Fisher Scientific) was used to determine RNA purity and concentration. The OD 260/280 absorbance ratio was between 1.8 and 2.0 in each sample. Total RNA was reverse transcribed with primers using a reverse transcriptase kit (PrimeScript TM RT Reagent Kit; TaKaRa Biotechnology Co. Ltd., Dalian, China). PCR was carried out with an ABI 7500 sequence detection system (Applied Biosystems; Thermo Fisher Scientific). All PCR reactions were performed in a total volume of 20 μL, and single transcripts of gene expression were determined with the SYBR® Premix Ex Taq TM II kit (TaKaRa Biotechnology Co. Ltd.), according to the manufacturer's instructions. Reactions began with a 10 s hot-start activation of the Taq polymerase at 95°C, followed by 40–45 cycles of amplification in three steps (denaturation at 95°C for 5 s, followed by a 30 s annealing at 60°C and a 30 s extension at 72°C). The reactions were carried out in a total volume of 20 μL in a 0.2 mL flat cap PCR tube (Axygen, Tewksbury, MA). The primers for PCR were as follows: HMGB1: 5′-TTA GTC CCA GCG AAG GCT AT-3′ (forward), 5′-CAA GTT TCC TGA GCA ATC CA-3′ (reverse); GAPDH: 5′-GCA AGT TCA ACG GCA CAG-3′ (forward), 5′-CGC CAG TAG ACT CCA CGA C-3′ (reverse).
Statistical analysis
All statistical analyses were performed using IBM SPSS Statistics for Windows v20.0 (IBM Corp., Armonk, NY). Results are displayed as means ± standard deviation. One-way analysis of variance was used to compare the effect of GL treatments in the three different groups. The Bonferroni test was used for pairwise comparisons. Graphpad Prism v5 was used to analyze study graphs. p-Values of <0.05 were considered statistically significant.
Results
HMGB1 mRNA expression in thyroid tissues at different time points
The dynamic changes of HMGB1 mRNA levels in the thyroids of NOD.H-2h4 mice were measured by PCR (Fig. 1). There were no significant differences between the control group and NaI group after two or four weeks of iodine treatment. However, HMGB1 mRNA expression was significantly higher in the NaI group compared with the control group after 8 and 16 weeks of iodine supplementation (p < 0.05).
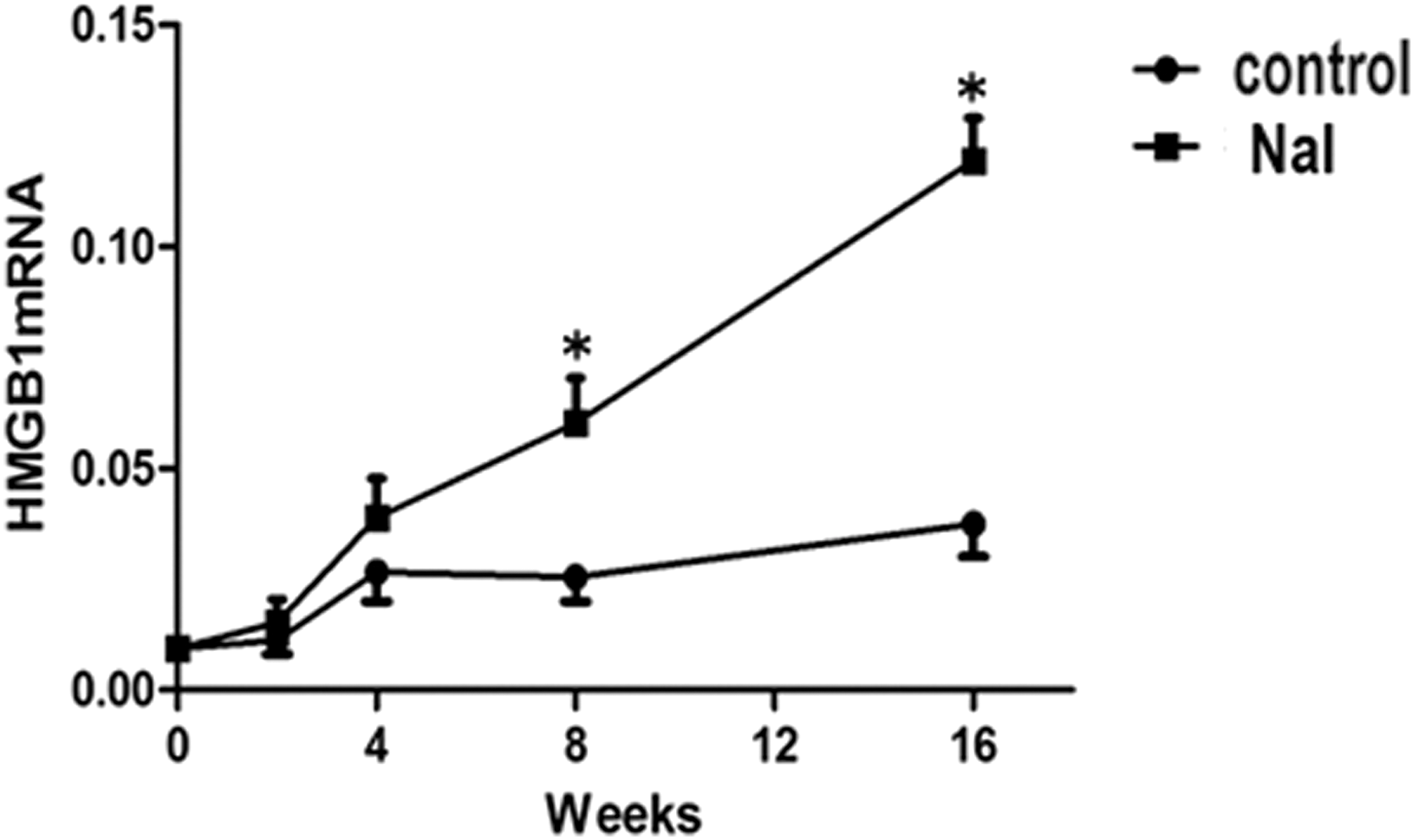
Messenger RNA expression of HMGB1 in thyroid glands of iodine (NaI)-treated mice and controls at different time points. NaI group: NOD.H-2h4 mice were fed with 0.005% sodium iodine water; control group: NOD.H-2h4 mice were fed with regular water. Values are shown as mean ± standard deviation (SD). Statistical analysis was conducted using a t-test. *p < 0.05 versus control at the same time point.
Thyroid weights
As shown in Figure 2A, the thyroid weights were significantly higher in the NaI groups compared with the control group (p < 0.05). The thyroid weights were remarkably lower in the NaI + GL group compared with the NaI group (p < 0.05).
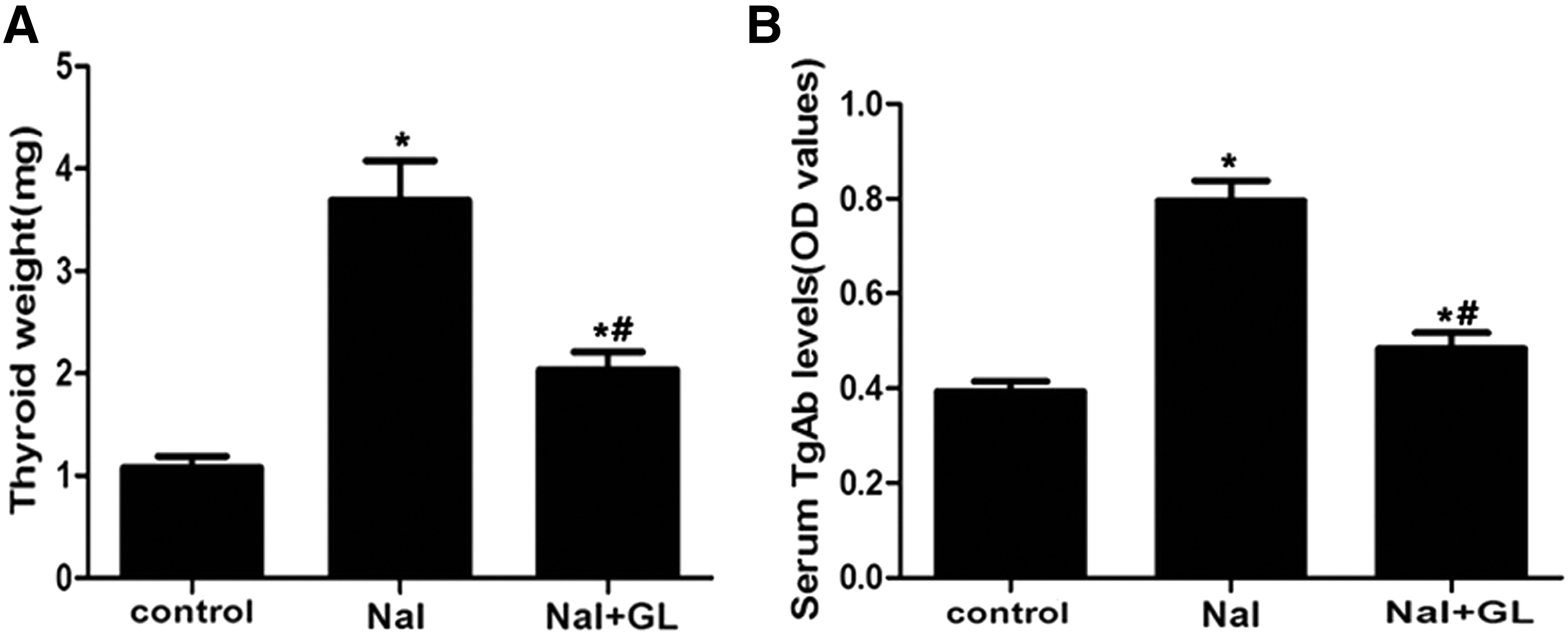
Thyroid weight and serum antithyroglobulin antibodies (TgAb) levels in different groups of NOD.H-2h4 mice. NaI group: NOD.H-2h4 mice were fed with 0.005% sodium iodine water; control group: NOD.H-2h4 mice were fed with regular water. GL, glycyrrhizin. (
Serum TgAb titers
Serum TgAb titers were remarkably higher in the NaI group (0.78 ± 0.09 OD NaI) compared with the control group (0.39 ± 0.03 OD; p < 0.05; Fig. 2B, control). Interestingly, the serum TgAb levels significantly decreased after supplementation with GL (0.51 ± 0.05 OD; p < 0.05; Fig. 2B, NaI + GL). The TgAb concentration was not significantly different between the control and GL treatment groups.
Thyroid histopathology
The thyroid follicles were relatively uniform and moderately sized in the center and larger in the periphery in the control group (Fig. 3A). Moreover, the follicular epithelial cells formed a monolayer with round or elliptical nuclei in the centers of the cells, and the thyroiditis scores ranged from 0 to 1+ (Fig. 3D, control). In the NaI group, the follicular cavities were enlarged, and the follicular epithelial cells were flat in shape. Lymphocytic infiltration was observed in the thyroids in almost 100% of NOD.H-2h4 mice given 0.005% NaI in water for eight weeks (Fig. 3B), and the thyroiditis scores ranged from 1+ to 4+ (Fig. 3D, NaI). In NaI + GL group, the follicular cavities were smaller, and the thyroiditis scores ranged from 0 to 2+ (Fig. 3D, NaI + GL). The severity of lymphocytic infiltration in the thyroid was lower in the NaI + GL compared with the NaI group (Fig. 3C). The thyroiditis scores in the NaI + GL and control groups were significantly lower compared with the NaI group (p < 0.05; Fig. 3D).
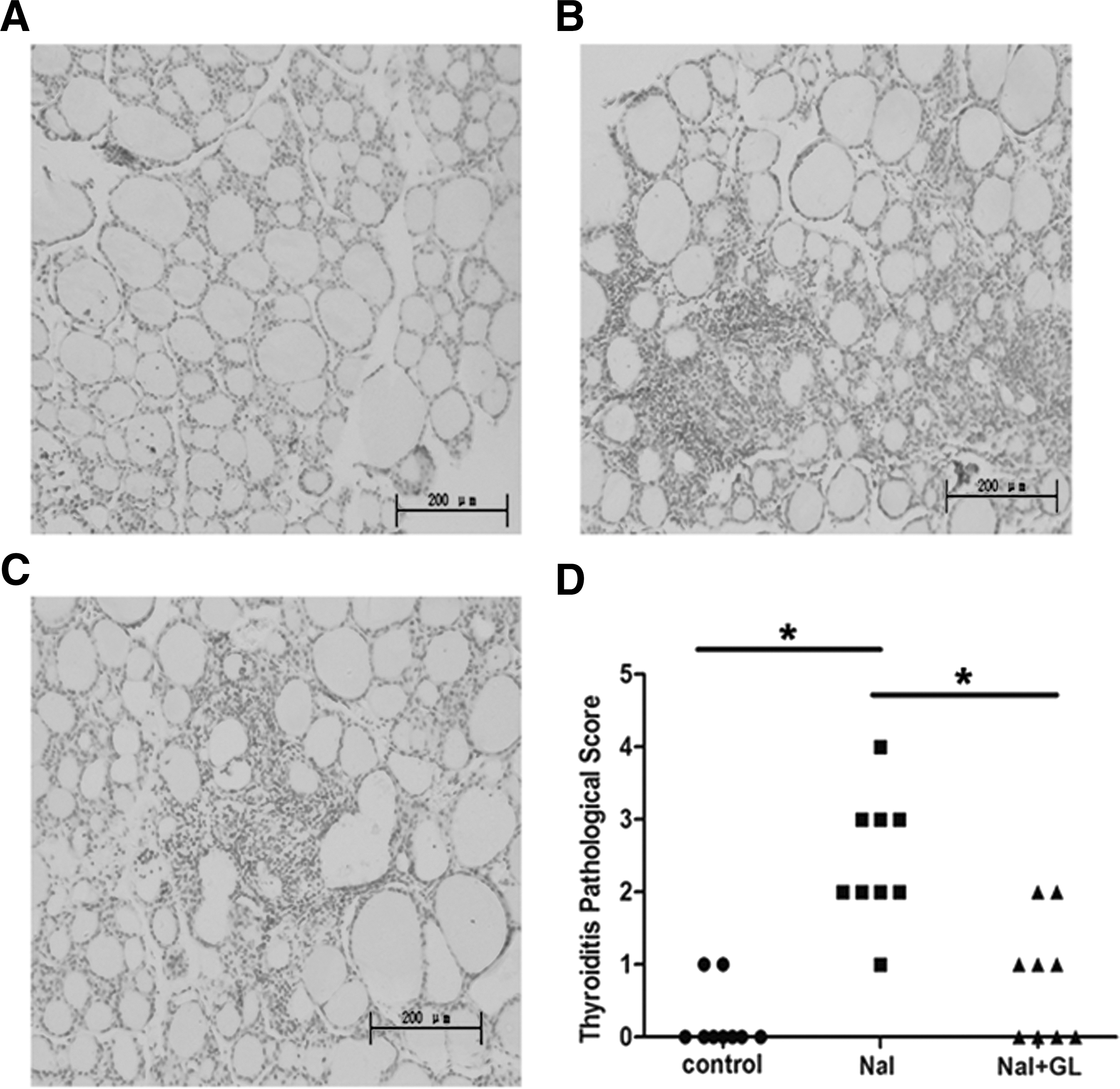
The effects of GL on thyroid histology of NOD.H-2h4 mice. Representative histology section of hematoxylin and eosin (H&E)-stained thyroid tissues from different groups of NOD.H-2h4 mice (H&E staining, magnification × 200). NaI group: NOD.H-2h4 mice were fed with 0.005% sodium iodine water; control group: NOD.H-2h4 mice were fed with regular water. (
Serum HMGB1 levels
HMGB1 serum levels in the NaI group (137.52 ± 15.92 pg/mL; Fig. 4, NaI) were significantly higher compared with those in the control group (22.37 ± 8.11 pg/mL; p < 0.05; Fig.4, control). Indeed, serum HMGB1 expression decreased remarkably in the NaI + GL group (74.88 ± 27.78 pg/mL; Fig. 4, NaI + GL) compared with the NaI group after four weeks of GL treatment (p < 0.05). These results suggest that GL treatment might be involved in the inhibition of HMGB1 expression during the development of autoimmune thyroiditis.
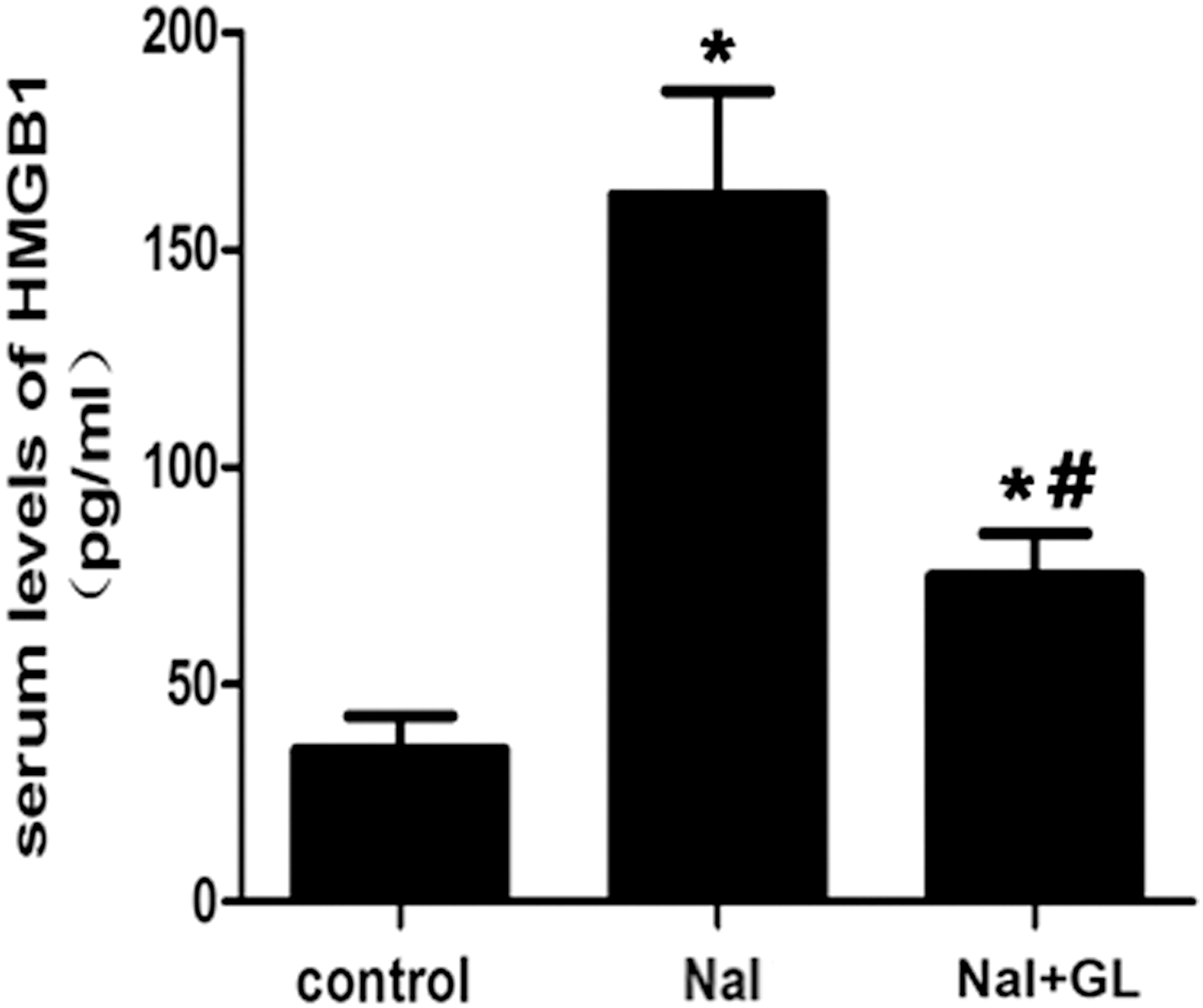
Serum HMGB1 levels in different groups of NOD.H-2h4 mice (pg/mL). NaI group: NOD.H-2h4 mice were fed with 0.005% sodium iodine water; control group: NOD.H-2h4 mice were fed with regular water. One-way ANOVA was used to compare the differences in multiple groups. *p < 0.05 versus control at the same time point. # p < 0.05 versus NaI at the same time point.
Serum cytokine assays
The serum levels of IL-6, IL-1β, and TNF-α were significantly higher in the NaI group compared with the control group (p < 0.05; Fig. 5A, B, and C, respectively). However, these three pro-inflammatory cytokines dramatically decreased after GL treatment (p < 0.05; Fig. 5A, B, and C, respectively). These findings suggest that GL administration might play a key role in the inhibition of the inflammatory response during the development of autoimmune thyroiditis.
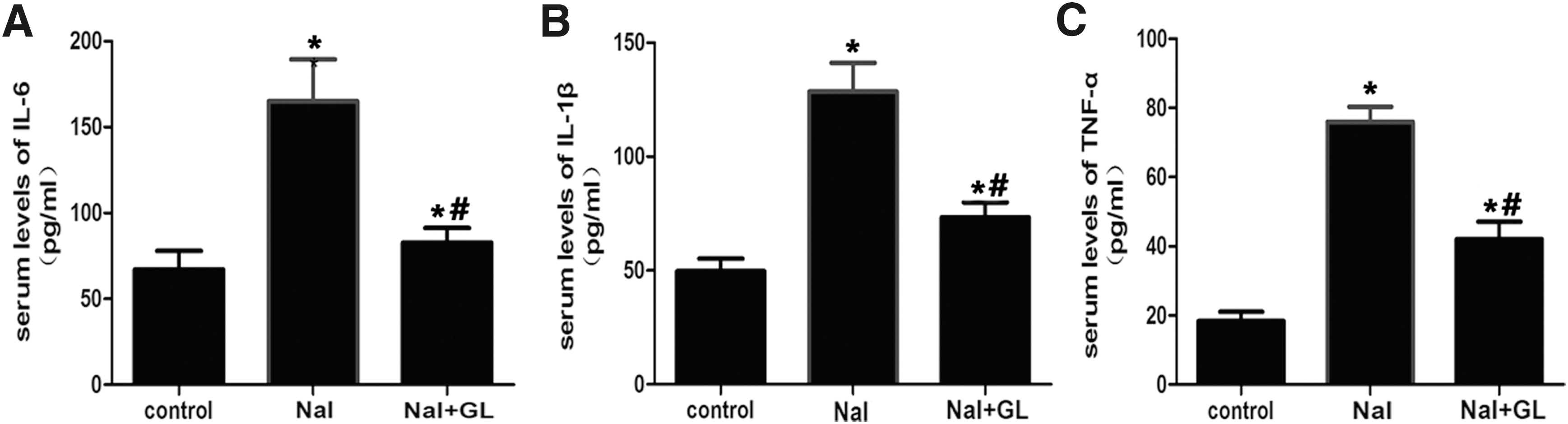
Serum levels of interleukin (IL)-6, IL-1β, tumor necrosis factor alpha (TNF-α) in different groups of NOD.H-2h4 mice. NaI group: NOD.H-2h4 mice were fed with 0.005% sodium iodine water; control group: NOD.H-2h4 mice were fed with regular water. (
HMGB1 expression in thyroid gland with GL treatment
HMGB1 was mildly expressed in follicular epithelial cells and lymphocyte cells of thyroid tissues in the control group (Fig. 6A) and significantly increased in the NaI group (Fig. 6B) compared with the control group. Surprisingly, HMGB1 expression was significantly reduced after GL treatment (Fig. 6C). Furthermore, HMGB1 expression was remarkably higher in the NaI group than it was in both the control and the NaI + GL groups, as found by Western blot experiments (p < 0.05; Fig. 7A and B). Similarly, real-time PCR analysis demonstrated that HMGB1 expression was significantly decreased in the NaI + GL group compared with the NaI group (p < 0.05; Fig. 7C). These results suggest that GL treatment decreases HMGB1 expression in thyroid tissue in this model of autoimmune thyroiditis.
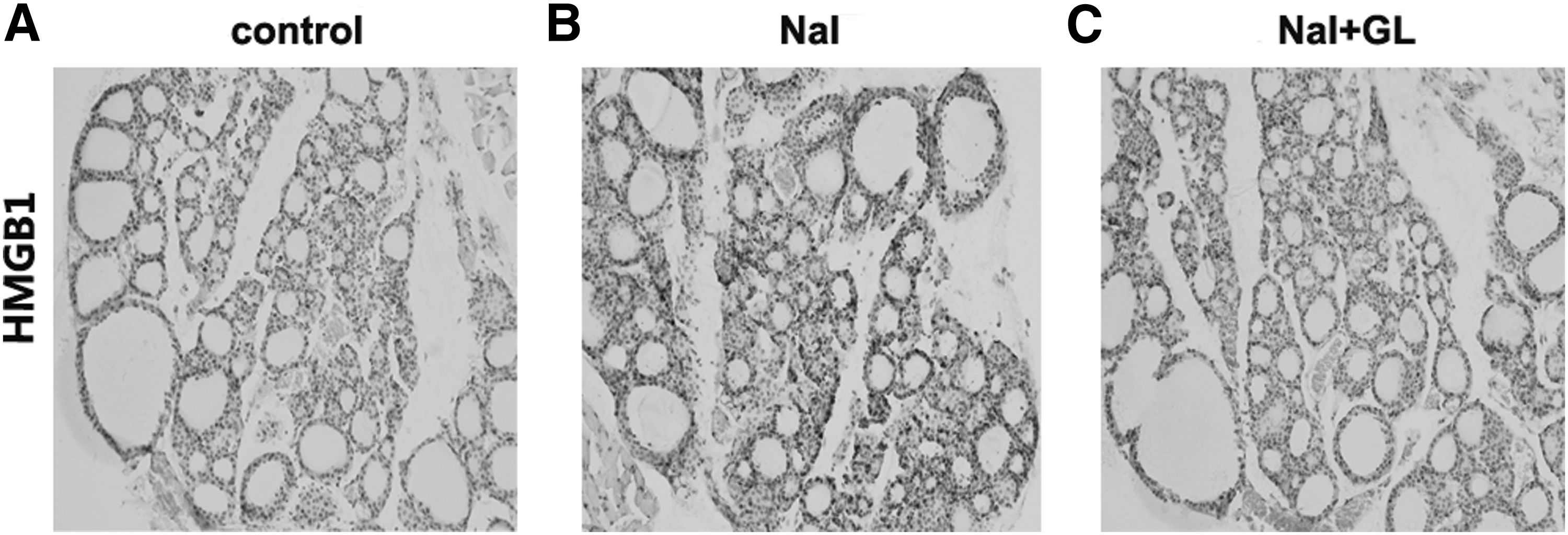
HMGB1 protein expression in thyroid tissue of NOD.H-2h4 mice in response to NaI and GL. (
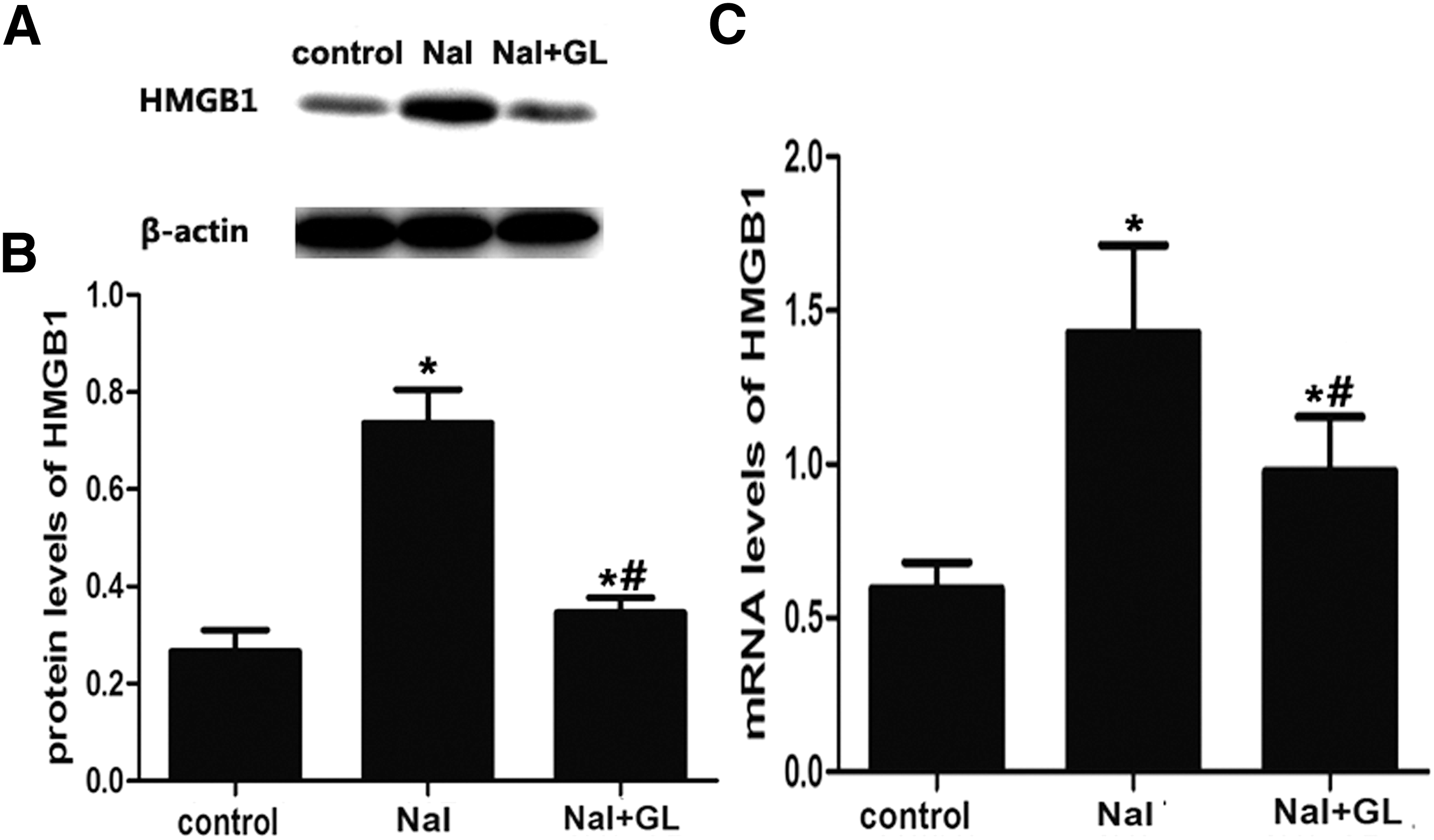
HMGB1 mRNA and protein expression in thyroid tissue of NOD.H-2h4 mice to determine the effect of GL. (
Pathway and molecular mechanism under Western blotting
The protein expression of TLR2, MyD88, and NF-κB was significantly elevated in the NaI group. The expression of TLR2 was significantly lower in the NaI + GL group compared with the NaI group (p < 0.05; Fig.8A and B). MyD88 expression in the NaI + GL and control groups was significantly lower compared with the NaI group (p < 0.05; Fig. 8C and D). The expression of NF-κB also decreased after GL treatment (p < 0.05; Fig.8E and F). There was no difference in the protein expression levels of TLR2, MyD88, and NF-κB between the NaI + GL and control groups.
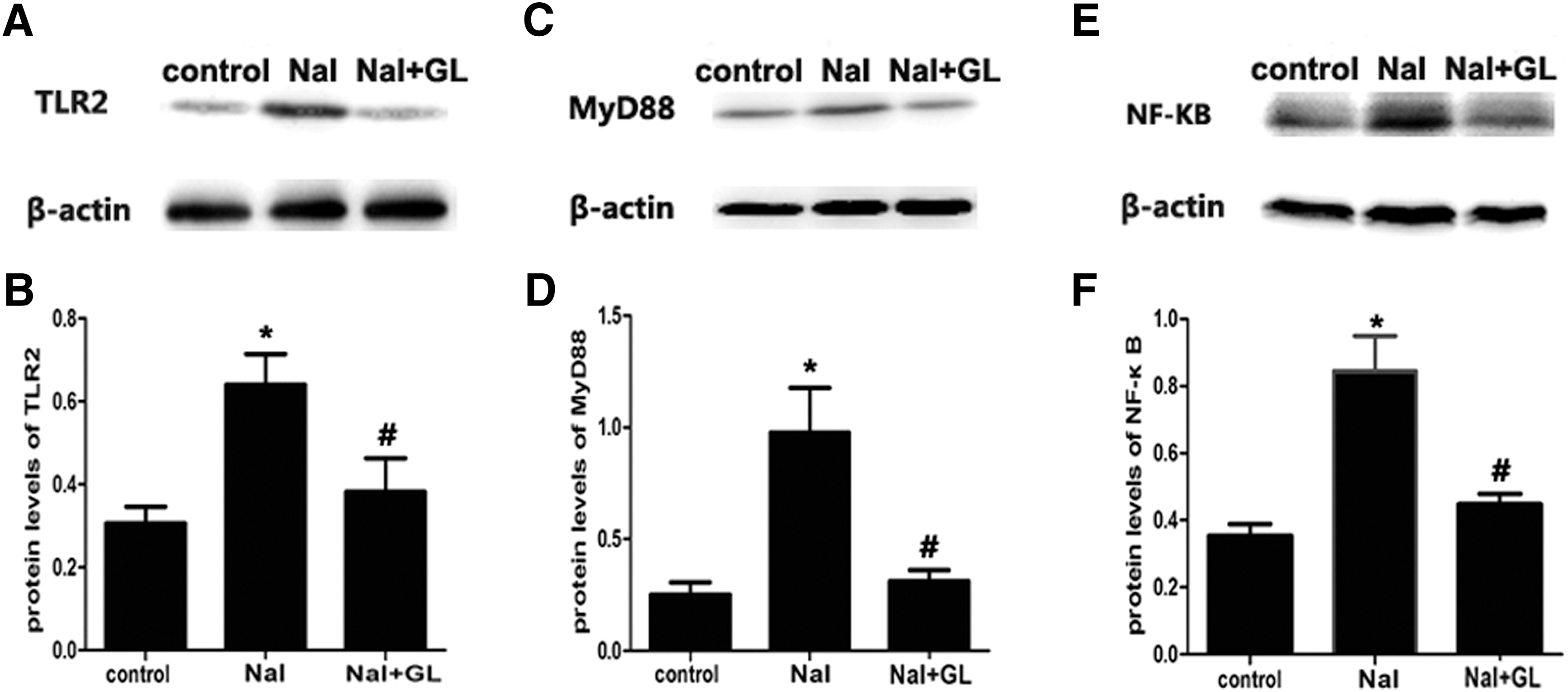
Protein expression in the Toll-like receptor 2 (TLR2) signaling pathway by Western blotting. (
Discussion
In this study, HMGB1 expression was significantly increased in the thyroid tissue and serum of NOD.H-2h4 mice with iodine-induced autoimmune thyroiditis. The study also demonstrated that the inhibition of HMGB1 via injection of GL decreased the lymphocytic infiltration of the thyroid and suppressed the expression of pro-inflammatory cytokines, such as TNF-α, IL-1β, IL-6, and downstream proteins including TLR2, MyD88, and NF-κB via the TLR2-HMGB1 signaling pathway. These findings suggest that HMGB1 may be involved in the pathogenesis of autoimmune thyroiditis.
AITD results from the loss of tolerance to self-antigens of the thyroid. This indicates that several factors participate in the development of thyroiditis, such as the environment and nutritional iodine intake. Iodine is a crucial element in the development of AIT, and both high and low iodine levels can increase the risk of AIT (20). NOD.H-2h4 mice, a mouse strain with a NOD genetic background, are prone to developing AIT and anti-mouse thyroglobulin (mTg) autoantibodies. Previous studies have shown that NaI, when given in excessive amounts in drinking water, can cause NOD.H-2h4 mice to develop AIT, which is a prototype mouse model of AIT in humans (21 –24). Lymphocytic infiltration in the thyroid, an increased relative thyroid weight, and a high serum TgAb titer are characteristics of iodine-induced thyroiditis in NOD.H-2h4 mice. The results of the present study confirm that excessive iodine can induce AIT and worsen the severity of disease in NOD.H-2h4 mice.
HMGB1 is a multifunctional protein that has a dual role. HMGB1 can bind DNA and facilities transcription, and it can also act as a cytokine by being passively released from necrotic cells inducing the early inflammatory response or it can be actively secreted from mononuclear macrophages. HMGB1 tends to activate pro-inflammatory responses by secreting cytokines and by stimulating antigen-presenting cells (6). HMGB1 is also involved in the development of inflammatory diseases by binding to RAGE or TLRs, which leads to further activation of the pro-inflammatory response and inflammatory injury (25). Many studies have shown that HMGB1 may act as promoter in the pathogenesis of many autoimmune diseases, such as RA and SLE (26,27). In this study, HMGB1 mRNA expression was significantly increased in the NaI group compared with the control group after iodine supplementation for eight weeks, and remained elevated after 16 weeks of iodine administration. HMGB1 is considered to be a late mediator of inflammation, and the fact that the occurrence of HMGB1 during the late phase of inflammation may provide an efficient therapeutic window for the treatment of chronic inflammatory diseases, and also act as an inhibitor of early pro-inflammatory cytokines, including TNF-α, IL-6, and IL-1β. A previous study showed that HMGB1 is upregulated in the thyroids of AIT patients and that HMGB1 expression is very low in healthy thyroids (28). Recently, HMGB1 has been considered a “danger signal” or alarmin, which is activated by the innate immune response system. Moreover, extracellular HMGB1 has been discovered to activate the innate immune response through the activation of antigen-presenting cells (APC). Recent studies have also shown that passive release of HMGB1 is a feature of apoptosis (29), and studies of immunostained pancreatic cells from NOD mice have found passive secretion of HMGB1 by infiltrated islets (30). HMGB1 expression in islet tissue also suggests that HMGB1 expression may be associated with the pathogenesis of type 1 diabetes. Recently, it was reported that NOD.H-2h4 thyrocytes exhibit a high sensitivity to NaI-induced apoptosis, which may play a role in the mechanism of AIT development (31). Apoptotic cascades in thyrocytes could be triggered by iodine and have been demonstrated both in vivo and in vitro.
GL is a traditional medicine derived from the root of licorice and is an HMGB1 inhibitor. A recent study demonstrated that GL could have an anti-inflammatory effect and protect spinal cord ischemic injury in rats by reducing the expression of inflammatory cytokines and inhibiting the production of HMGB1 (32). It has been reported that GL significantly reduces the expression of pro-inflammatory cytokines, including TNF-α, IL-1β, and IL-6, by inhibiting HMGB1 chemo-attraction and mitogenic activity (33). GL is also often used in liver disease due to its antiviral effects (34).
In this study, HMGB1 protein and mRNA expression, as well as HMGB1 serum levels, were high in the thyroids of NOD.H-2h4 mice treated with NaI, indicating that HMGB1 may participate in the pathogenesis of autoimmune thyroiditis. In addition, early pro-inflammatory cytokines, such as TNF-α, IL-1β, and IL-6, were elevated in conjunction with the production of HMGB1. In the NaI-treated group, these three cytokines reached much higher levels compared with those in the control group. In this study, GL not only decreased the serum levels of the pro-inflammatory cytokines including TNF-α, IL-6 and IL-1β, but also reduced the serum level of HMGB1 in the NaI-treated group. In addition, GL reduced the expression of HMGB1 in the thyroids of iodine-induced thyroiditis in NOD.H-2h4 mice. This suggests that the inhibition of HMGB1 by GL is likely associated with a decrease of tissue injury by attenuating the inflammatory response. The NaI-treated group had lymphocytic infiltration of the thyroid with severity scores of 1+ to 4+, along with higher thyroid weights and serum TgAb titers compared with mice in the control group. There was a reduction in the severity of lymphocytic infiltration in the thyroids of NaI + GL mice in the current study. After GL treatment, mice had lymphocytic infiltration severity scores of 0+ to 2+, along with lower thyroid weights and reduced TgAb titers. This suggests that GL therapy significantly alleviated thyroid tissue damage and reduced the incidence of thyroiditis.
The current study also examined whether the anti-inflammatory effect of GL occurred via activation of the TLR2 signaling pathway. The expression of TLR2 downstream adapter proteins was first analyzed by Western blot. The expression of TLR2 signal molecules in the NaI-treated group was significantly higher than in the control group, suggesting that the activation of the TLR2 signal may play an important role in the development of autoimmune thyroiditis. The results show that GL inhibits the expression of TLR2, suggesting that the molecular target of GL is TLR2. Moreover, NF-κB and MyD88 protein expression was also inhibited by GL. However, the mechanisms for the effects of GL remain unclear. HMGB1 interacts with several receptors, including TLR2, TLR4, and TLR9, thus activating the expression of NF-κB, and induces the production of proinflammatory cytokines, which promote tissue damage and inflammation (35). The interaction of HMGB1 with TLR4 was previously examined in isolated islets (36). Results of that study suggested that the TLR4 signaling pathway may participate in the development of autoimmune diabetes (36). TLR4 expressed on islet β cells senses the changes of HMGB1 and contributes to the initiation of type 1 diabetes. It has been demonstrated that HMGB1 antibody treatment in NOD mice not only prevents the progression of insulitis, but also significantly reduced the incidence of diabetes. GL is a natural anti-inflammatory compound that is usually used to treat patients with chronic inflammatory diseases. Thus, GL may be a compound that is useful for the prevention and treatment of autoimmune thyroiditis.
There are several limitations in the present study. GL attenuated the severity of AIT and downregulated the expression of HMGB1. However, ROS and apoptotic markers in thyroid tissue were not measured. Further studies are needed to evaluate the antioxidant and apoptotic activities of GL treatment in the context of AIT.
In conclusion, the results of the current study suggest that there is evidence that HMGB1 is involved in the pathogenesis of autoimmune thyroiditis. GL seems to inhibit the progression and the severity of AIT significantly in this murine model using NOD.H-2h4 mice, and the mechanism for this inhibition may be related to an attenuation of the inflammatory response via the TLR2-HMGB1 signaling pathway.
Footnotes
Acknowledgments
The study was supported by the Chinese National Natural Science Foundation (Grant 81400777, U1508219, 81570708) and program for Liaoning Excellent Talents in University (Grant LJQ2015115).
Author Disclosure Statement
The authors declare no conflict of interests.