
Abstract
Background:
Macro-thyrotropin (TSH) is a high-molecular-weight form of TSH. Most cases of macro-TSH are TSH complexed with immunoglobulin G. This study was undertaken to characterize macro-TSH.
Methods:
Blood samples taken from patients with subclinical hypothyroidism were screened for the presence of macro-TSH with the polyethylene glycol method and confirmed with gel filtration chromatography. TSH receptor antibody was quantified with an electrochemiluminescence immunoassay. Binding studies were performed using 125I-human TSH, and the specificity of anti-TSH autoantibodies was tested by displacement experiments using excess amounts of the unlabeled related peptides. Macro-TSH and serum TSH levels were evaluated twice over a one- to four-year interval.
Results:
Sixteen patients (11 females and 5 males; aged 8–82 years) were diagnosed as having macro-TSH. None of the patients with macro-TSH tested positive for TSH receptor antibody. Judging from the affinity and the binding capacity of anti-TSH autoantibodies, two classes of binding sites were identified. Regarding specificity, there were anti-human TSH-β autoantibodies that were partially cross-reactive to bovine and/or rat TSH-β. There were also autoantibodies against human glycoprotein α, a common subunit among human TSH, luteinizing hormone, and follicle stimulating hormone. Macro-TSH persisted in 11/13 patients who could be reevaluated over a one- to four-year interval after the first evaluation. Serum TSH levels returned to normal in the remaining two patients whose macro-TSH disappeared.
Conclusions:
It is concluded that anti-human TSH autoantibodies are a major cause of macro-TSH and that macro-TSH may persist for a long time.
Introduction
M
Macro-TSH can be precipitated with 12.5% polyethylene glycol (PEG) (3 –8) and binds to both protein A and protein G in most cases (2 –4,8), leading to the proposal that macro-TSH is a complex of TSH and immunoglobulin G (IgG). Because the binding of TSH to IgG is reversible (2,4,6,8), IgG that binds in the complex may be an anti-TSH autoantibody.
An analogous situation exists with macroprolactinemia, a major cause of hyperprolactinemia (10 –14). Macro-prolactin (macro-PRL) has characteristics that are pathophysiologically similar to macro-TSH. Notably, macro-PRL has a molecular mass of >150 kDa, has low bioactivity, and appears to be a complex of prolactin (PRL) and IgG, especially anti-PRL autoantibodies (13 –15).
This study sought to characterize macro-TSH. It attempted to establish whether macro-TSH comprises a complex of TSH and anti-human TSH autoantibodies, and to determine whether anti-human TSH autoantibodies were identical to anti-idiotype antibodies for TSH receptor antibody (TRAb). Furthermore, the study assessed the persistence of macro-TSH. It is clinically important because patients with elevated serum macro-TSH do not require hormone replacement therapy due to its low activity. If macro-TSH persists for a long time, assessing its presence will become crucial for the differential diagnosis of hyperthyrotropinemia and for determining the most effective therapeutic strategy.
Materials and Methods
Patients
Patients with subclinical hypothyroidism who were being followed at Kobe City General Hospital Outpatient Clinic or who had been referred from other hospitals were examined to see if they had macro-TSH. The serum samples were screened with the PEG precipitation method, and those with a PEG precipitation ratio >75% were subjected to gel filtration chromatography to confirm macro-TSH, as previously described (8). Sixteen patients (11 females and 5 males; aged 8–82 years) were diagnosed as having macro-TSH. The clinical characteristics of the 16 patients are shown in Table 1. Patients 1, 4, 5, and 10 had Hashimoto's thyroiditis. Patient 2 had hypothyroidism due to subtotal thyroidectomy for the treatment of Graves' disease. Patient 6 had systemic lupus erythematosus (SLE). Patient 8 had systemic sclerosis (SSc). Patients 12 and 13 had rheumatoid arthritis (RA), and were all treated with steroids. Patients 7, 11, and 13 had hepatitis C, and were treated with interferon. Patient 14 had also been diagnosed with macroprolactinemia. An endocrine workup was performed in patient 16, who visited a hospital with unspecific complaints and without any symptoms of hypothyroidism. Regarding patients 3, 9, and 15, it was not apparent why the doctors ordered thyroid testing from their medical records. None of these patients had a history of injections of human thyrotropin-α or human chorionic gonadotropin. Patients with subclinical hypothyroidism that had not been attributed to macro-TSH comprised the control group. This group contained five women and five men, aged 21–58 years. This study was approved by the Clinical Research Review Board of Kobe City General Hospital.
Diseases: SLE (systemic lupus erythematosus), SSc (systemic sclerosis), RA (rheumatoid arthritis).
Non-PEG(polyethylene glycol)-precipitable TSH: TSH concentrations in the supernatant after PEG precipitation.
Ratio of PEG-precipitable TSH.
Ratio of large molecular-sized TSH (>150 kDa) on gel filtration chromatography.
Antithyroid peroxidase antibody.
Antithyroglobulin antibody
Proportion of serum TSH concentrations assayed with HAMA (human anti-mouse antibodies) blockers/those without HAMA blockers. The proportion in controls was 104.3 ± 12.4%.
TSH, thyrotropin; fT4, free thyroxine.
TSH assays
The concentration of TSH was measured with an enzyme immunoassay (EIA) that was established in the laboratory (8). In brief, human TSH-β monoclonal antibody (10-T25C; Fitzgerald Industries International, Acton, MA) was used as a capture antibody, and human TSH-β monoclonal antibody (10-T25B; Fitzgerald) was used as a detection antibody. TSH was sandwiched between a capture antibody-coated polystyrene ball and a detection antibody-horseradish peroxidase (HRP) conjugate. The activity of HRP bound to the ball was assayed in a reaction with 3-(p-hydroxyphenyl) propionic acid (Sigma–Aldrich, St-Louis, MO) as a substrate by measuring fluorescence intensity with a spectrofluorophotometer (FP-6200ST; JASCO Co., Tokyo, Japan), with excitation at 320 nm and emission at 405 nm. Human TSH (AFP3959A) used for the reference preparations (hTSH-RP-2) was a highly purified human pituitary TSH, which was provided by Dr. A. F. Parlow (National Hormone and Peptide Program, Harbour-UCLA Medical Center, Torrance, CA). The limit of detection was 0.0075 mIU/L (conversion factor from ng/mL to mIU/L is 7.54). Based on a serum sample with a total TSH concentration of 4.5 mIU/L, the intra- and inter-assay coefficients of variation were 3.5% and 5.2%, respectively.
PEG precipitation
PEG was used to precipitate γ-globulin associated with TSH, as previously reported (8). In brief, serum samples of 50 μL were treated either with 50 μL of 25% PEG (final concentration of PEG was 12.5%) to precipitate γ-globulin fractions, leaving the non-PEG-precipitable TSH in the supernatant, or with 50 μL of water instead as a control (serum TSH). The percentage of PEG-precipitable TSH, which may be increased in sera containing macro-TSH, was calculated as follows: (serum TSH – non PEG – precipitable TSH)/serum TSH × 100. It was 61.8 ± 7.5% in 10 control patients.
Gel filtration chromatography
Gel filtration chromatography was performed with a 1 cm × 65 cm column of Ultrogel AcA 44 (IBF, La Garenne, France) equilibrated with 0.01 mol/L sodium phosphate buffer (pH 7.0), containing 0.1 mol/L NaCl, 0.1% (w/v) bovine serum albumin, and 0.05% (w/v) NaN3. Serum samples (50–500 μL) were applied to the column, and 1 mL fractions were collected for TSH measurements. The column was calibrated with various molecular weight markers (Sigma–Aldrich).
Thyroid-associated autoantibodies and thyroid hormone
Thyroid-associated autoantibodies and free thyroxine (T4) were assessed with electrochemiluminescence immunoassays and MODULAR ANALYTICS (Roche Diagnostics GmbH, Mannheim, Germany). Antibodies to TRAb were quantified with Elecsys TSHR, with a reference range <2.0 IU/L. Antithyroid peroxidase (TPO) and thyroglobulin (Tg) antibodies were quantified with Elecsys Anti-TPO and Anti-Tg, respectively. The reference ranges of anti-TPO antibody and anti-Tg antibody were <16 IU/mL and <28 IU/mL, respectively. Free T4 concentrations were quantified with Elecsys fT4, with a reference range of 0.81–1.58 ng/dL. The conversion factor of free T4 concentrations from ng/dL to pmol/L is 12.87.
Human anti-mouse antibodies
Human anti-mouse antibodies (HAMA) can produce spuriously high TSH values in sandwich-type immunoassay systems employing two mouse monoclonal antibodies (16) because HAMA bridges the capture antibody and detection antibody in the absence of TSH. Three HAMA blockers were used to counteract the effects of HAMA: mouse whole serum (MP Biomedicals, Santa Ana, CA), mouse IgG (THBR1), and mouse IgM (THBR2; the latter two from Cosmobio Co., Tokyo, Japan). The patients were considered to carry HAMA when TSH values decreased upon addition of the blockers.
125I-hTSH binding study
Highly purified human TSH (hTSH; National Institute of Diabetes and Digestive and Kidney Diseases [NIDDK] hTSH-SIAFPI-8; AFP-3959A), rat TSH (rTSH; NIDDK rTSH-I-9; AFP-11542B), human luteinizing hormone (hLH; National Hormone and Peptide Program [NHPP] hLH; AFP6780C), human follicle-stimulating hormone (hFSH; NHPP hFSH; AFP7298A), human chorionic gonadotropin-α (hCG-α; NHPP hCG-α; AFP8492A), and hCG-β (NHPP hCG-β; AFP8494A) were all purified synthetic peptides, and were provided by Dr. A. F. Parlow. Bovine TSH (bTSH) was purchased from Sigma–Aldrich. Human TSH was radio-iodinated with Na125I (PerkinElmer, Waltham, MA) in an IODO-GEN pre-coated iodination tube (Pierce, Rockford, IL), according to the manufacturer's instructions. An 125I-hTSH binding study was performed by incubating 20 μL of serum with 125I-hTSH (23,000 cpm/4 μL) at 37°C for 1 h, after which 24 μL of 25% cold PEG solution were added and mixed vigorously. The mixture was centrifuged, and the pellet was washed once with 500 μL of 12.5% PEG. The mixture was centrifuged again, and the radioactivity in the pellet was measured with a gamma counter (Wizard 3; PerkinElmer). This protocol is referred to as the “binding protocol.”
Displacement experiments were undertaken to examine the specificity of anti-TSH autoantibodies. Twenty microliters of serum were mixed with an excess (4 μg or 16 μg) of unlabeled peptides, which included hTSH, bTSH, rTSH, hLH, hFSH, hCG-α, and hCG-β. Then, 125I-hTSH (23,000 cpm) was added, and the mixture was assessed according to the binding protocol. To determine the affinity and binding capacity of anti-TSH autoantibodies, 20 μL of serum was mixed with 125I-hTSH (23,000 cpm) and various amounts of unlabeled hTSH (0.004–16 μg), and the mixture was assessed according to the binding protocol. Binding data were analyzed with nonlinear regression using GraphPad Prism v6 (GraphPad Software Inc., La Jolla, CA). The binding data were also displayed with a Scatchard plot.
Statistical analysis
All measurements are expressed as means ± standard deviation (SD). Values away from the mean ± 2SD were considered to be significantly different.
Results
Clinical characteristics of patients with macro-TSH
All patients diagnosed with macro-TSH had elevated serum TSH but normal free thyroid hormone levels (Table 1). When macro-TSH was removed from the serum by precipitating it with PEG, the non-PEG-precipitable TSH concentrations were all within the reference range, except for patient 4. Gel filtration chromatography revealed that the percentage of TSH that was a high-molecular-weight form ranged from 17.0% to 100%, all of which were significantly higher than that in controls (0.9 ± 1.2%). Anti-TPO antibodies were positive in patients 1 and 5. Anti-Tg antibodies were positive in patients 1, 4, 5, and 10. The proportion of serum TSH concentrations assayed with HAMA blockers/those without HAMA blockers in controls was 104.3 ± 12.4%. HAMA blockers did not significantly reduce the serum TSH levels in any of the 16 patients, indicating that the presence of HAMA was unlikely in these sera.
Characterization of anti-TSH autoantibodies
It is possible that anti-idiotype antibody for the TRAb is an anti-TSH antibody and forms macro-TSH, as illustrated in Figure 1. This possibility led to the assessment of TRAb in the patients in this study. None of them tested positive for TRAb (TRAb <0.3 IU/L).
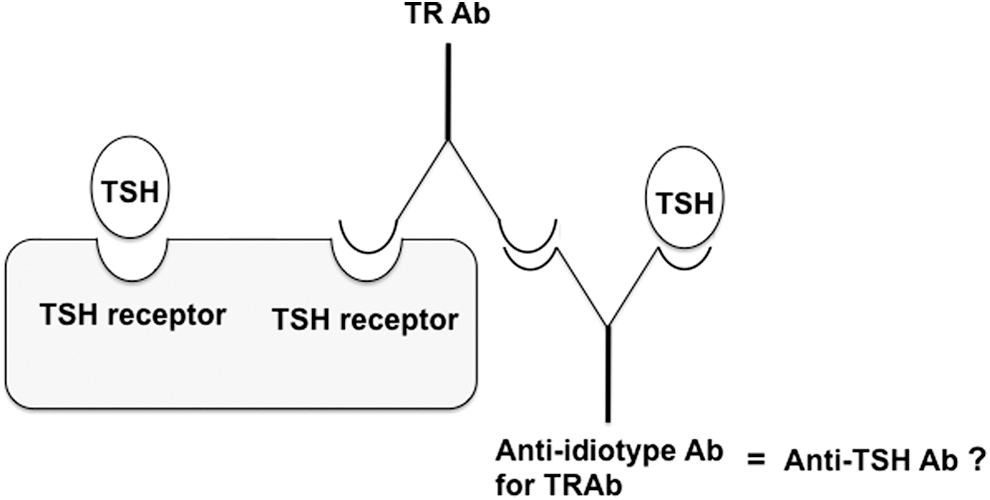
Illustration of the relationship between thyrotropin (TSH) receptor antibody (TRAb) and anti-idiotype antibody for TRAb.
Studies of hTSH binding to serum components were conducted with 125I-hTSH. This testing was performed on serum from six patients, the only sera available for this assay. The amount of 125I-hTSH in the bound fraction of the sera from these patients has been expressed in cpm, which represents the bound radioactivity. In sera from patients with macro-TSH, the results ranged from 9926 to 17,648 cpm compared with 5420 ± 576 cpm in sera from control patients. All binding values from patients with macro-TSH were significantly greater than those in the control group.
The specific binding, assessed by displacement of labeled hTSH by excessive amounts of the unlabeled peptides of interest, is shown in Figure 2. In sera from patients 1, 3, 14, and 7, 125I-hTSH was almost completely displaced by 4 μg of unlabeled hTSH. 125I-hTSH was also partially displaced by bovine and/or rat TSH in sera from patients 1, 3, and 14. In serum from patient 7, 125I-hTSH was not displaced by bovine TSH, rat TSH, and hCG-β, but was partially displaced by human LH, human FSH, and hCG-α. Although there was a considerable amount of 125I-hTSH bound to the sera from patients 2 and 15, the binding was barely displaced by 4 μg of unlabeled hTSH and the other related peptides. A greater amount of unlabeled hTSH (16 μg) partially displaced the bound, labeled hTSH.
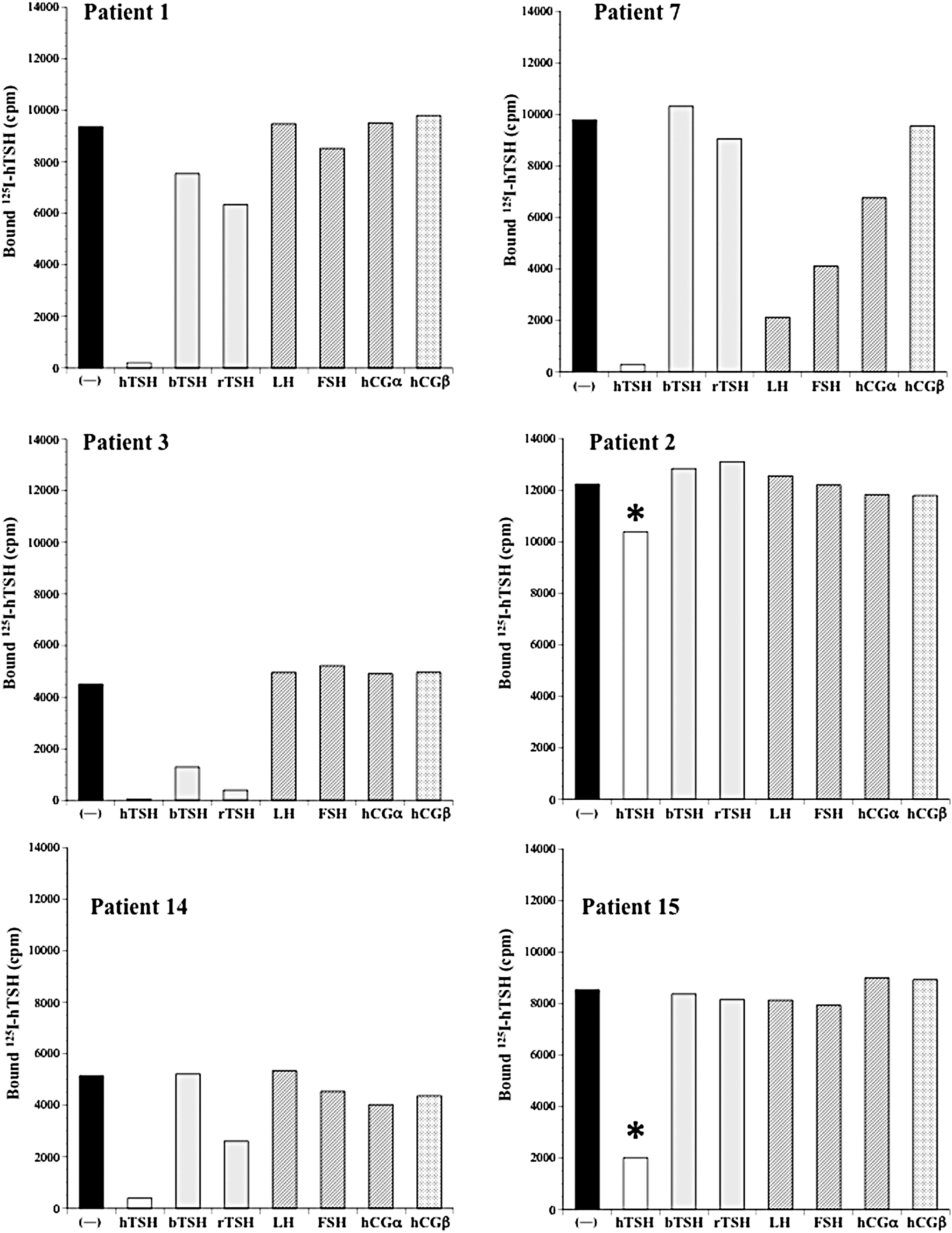
Specificity of anti-TSH autoantibodies in patients with macro-TSH. 125I-human TSH (hTSH) and sera from patients with macro-TSH were incubated either in the absence of (−) or with excess unlabeled hTSH, bovine TSH (bTSH), rat TSH (rTSH), human LH (hLH), human FSH (hFSH), human chorionic gonadotropin-α (hCG-α), and hCG-β. *In patients 2 and 15, 16 μg of unlabeled human TSH was used for the displacement instead of 4 μg in patients 1, 3, 14, and 7.
Nonlinear regression analysis showed that the displacement curves fit well to the two-binding-site model for patients 1, 3, 14, and 7, and to the one-binding-site model for patients 2 and 15 (Fig. 3A). The values for the affinity constant (Ka) of anti-TSH autoantibodies in sera from these patients were determined in this analysis and are shown in Table 2. A Scatchard plot confirmed that patients 1, 3, 14, and 7 had anti-TSH autoantibodies with two binding components: one with high affinity and low binding capacity, and the other with lower affinity and higher binding capacity, compared with the first. Patients 2 and 15 had anti-TSH autoantibodies with one component with low affinity and high binding capacity (Fig. 3B and Table 2).
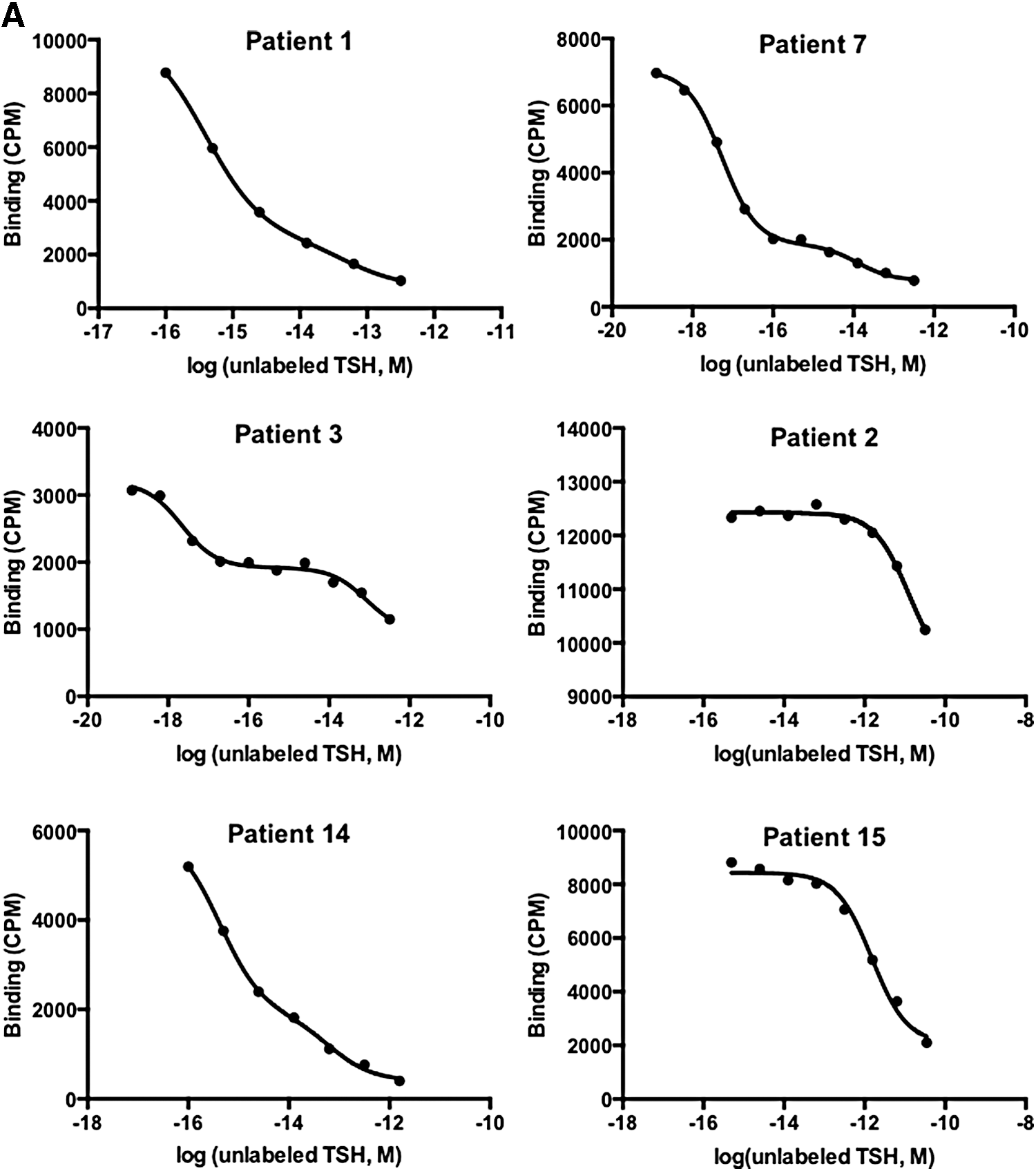
Analysis of binding studies of anti-TSH autoantibodies in patients with macro-TSH. (
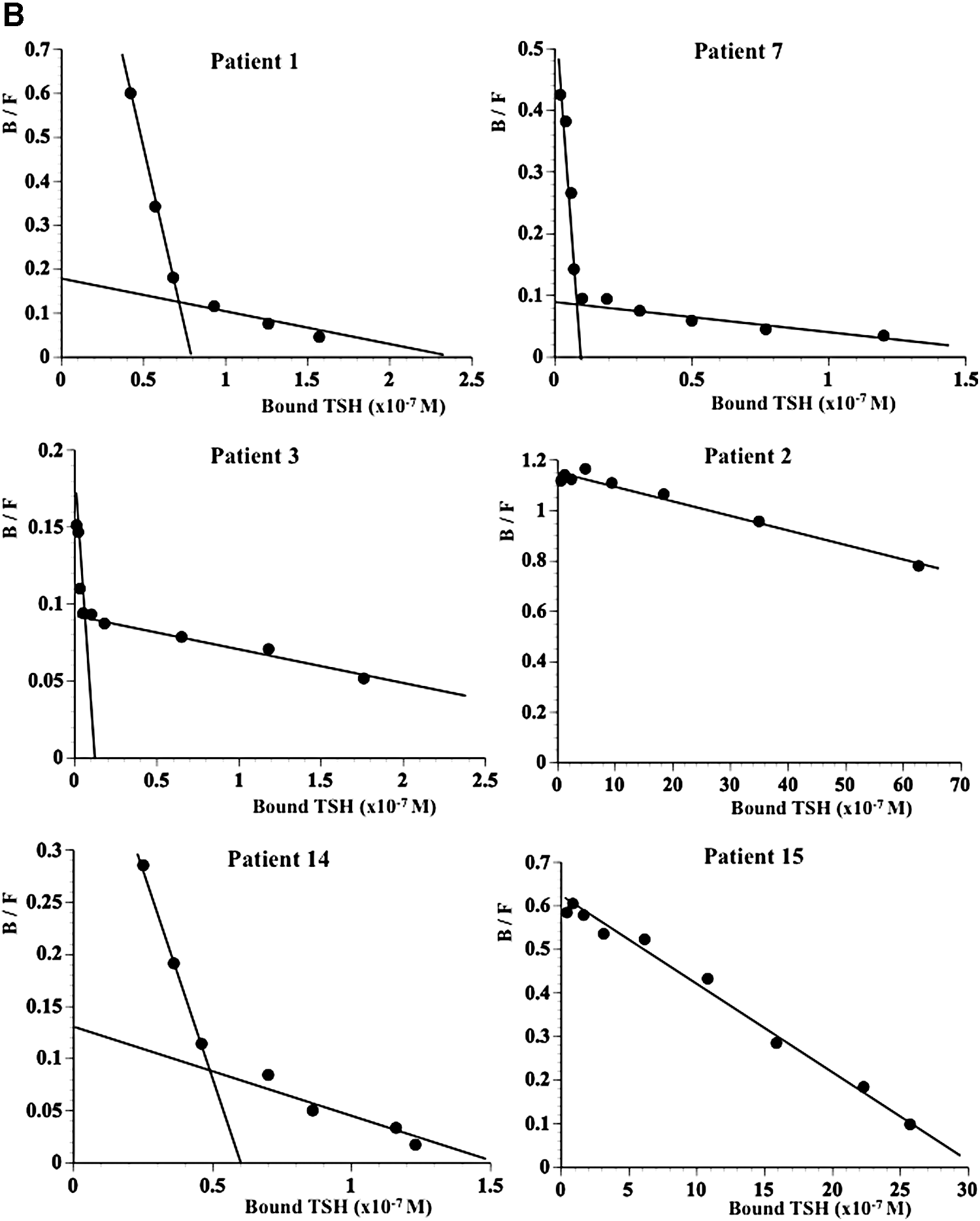
When two binding sites exist, those with lower affinity and higher binding capacity are designated as K2a and B2max, respectively.
K1a, affinity constant; B1max, maximal binding capacity.
Changes in macro-TSH and serum TSH levels over a long period
The changes in the ratios of macro-TSH and in serum TSH levels were examined over one to four years in 13 patients with macro-TSH (Fig. 4). Macro-TSH persisted in patients 1–11, while it disappeared in patients 12 and 13 whose serum TSH levels decreased within the reference range. In 11 patients with persisting macro-TSH, serum TSH levels were still elevated in patients 1–6, while they were normalized in patients 7–11. Eight patients (patients 1, 2, 4, 5, 6, 7, 10, and 13) were under thyroid hormone replacement therapy on the second evaluation, and patient 2 had also received thyroxine on the first evaluation.
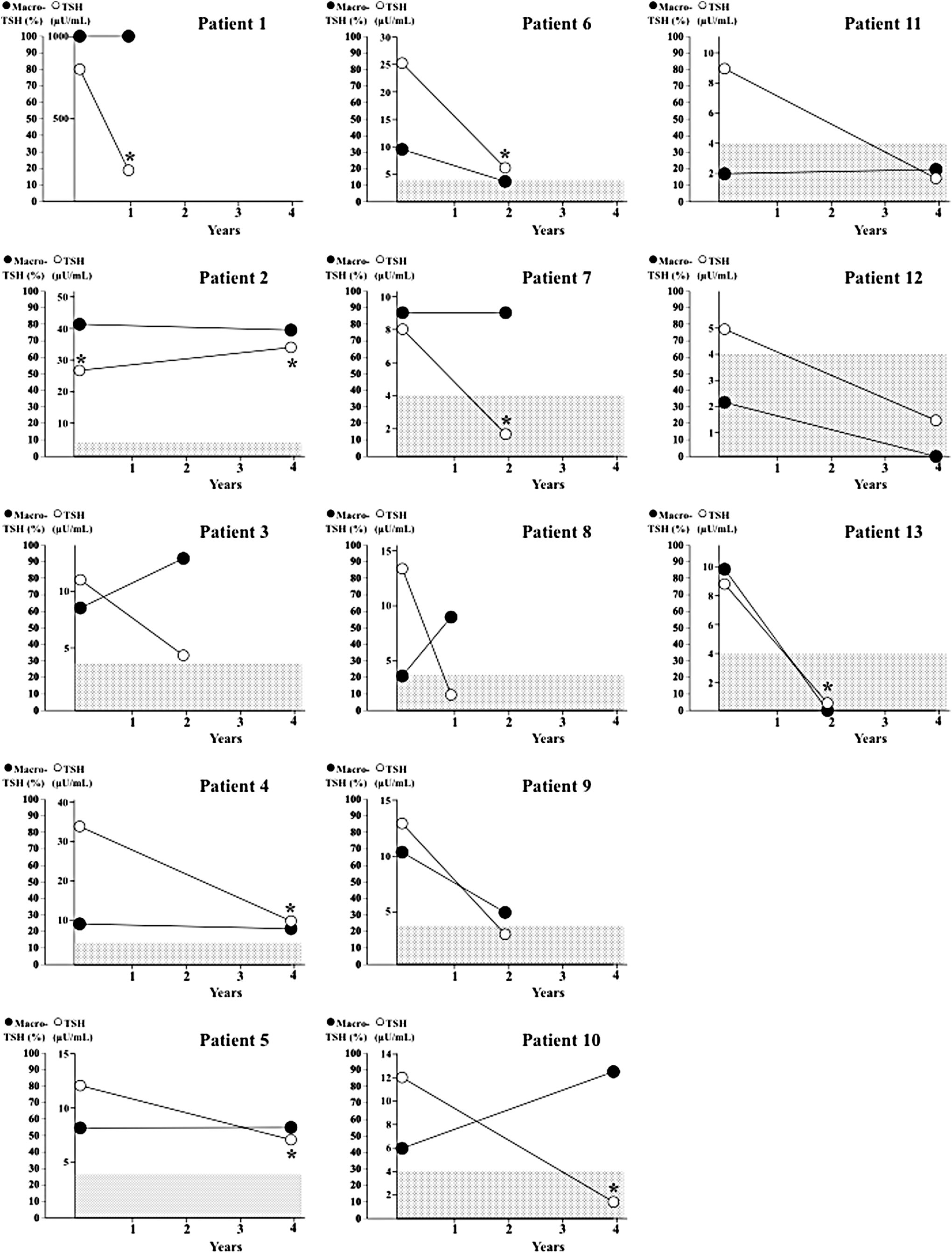
The changes in the ratios of macro-TSH and serum TSH levels over an interval of one to four years between the first and the second evaluation. Closed circles, ratios of macro-TSH; open circles, serum TSH levels; dotted areas, serum TSH reference range. *Patients were under thyroid hormone replacement therapy when blood samples were taken.
Discussion
This study demonstrates that anti-human TSH autoantibodies are a major cause of macro-TSH. There are several case reports addressing TSH binding components in some patients' blood. Anti-bovine TSH autoantibody or bovine TSH-binding immunoglobulins have been detected in some patients with Graves' disease or Hashimoto's thyroiditis (17 –19). Antibodies to human TSH were first detected in sera from patients with hypopituitarism who had undergone surgery for craniopharyngioma that was accompanied by injections of bovine TSH or human chorionic gonadotropin (20). Subsequently, anti-human TSH autoantibodies were found in sera from some healthy infants with transient neonatal hyperthyrotropinemia as well as their mothers (21,22).
Several research teams found anti-human TSH autoantibodies in sera containing thyroid-stimulating immunoglobulins in some patients with Graves' disease, claiming that the anti-idiotype antibody for TSH receptor antibody, TRAb, might be an anti-TSH antibody (23,24). Antibodies against the variable region of TRAb—anti-idiotype antibody for TRAb—may not only bind to TRAb but also to TSH, as shown in Figure 1. Nonetheless, in this study, TRAb was not detected in any of the patients with macro-TSH. Thus, TRAb is unlikely to be a predominant mechanism in the formation of macro-TSH.
All six patients with macro-TSH who were tested in the binding study had anti-human TSH autoantibodies with various characteristics in regards of the specificity. One patient with macro-TSH (patient 7) had anti-human TSH autoantibodies for which binding was not displaced by excess bovine TSH, rat TSH, or hCG-β, but was displaced by human TSH, human LH, human FSH, and hCG-α. TSH, LH, FSH, and hCG consist of α- and β-subunits, and the α-subunit is a glycoprotein that is common to all of these peptides (25). These results could therefore be explained if this patient has autoantibodies against the human α-subunit. Three patients (1, 3, and 14) had anti-human TSH autoantibodies that partially recognized bovine and/or rat TSH in addition to human TSH. Human TSH-β consists of 118 amino acids and exhibits 89.0% and 90.6% homology with the amino acid sequences of bovine TSH-β and rat TSH-β, respectively (26). It may be that the epitopes on TSH that are shared in part among different species may have been antigenic and led to the production of anti-human TSH-β autoantibodies that partially cross-react to bovine and/or rat TSH-β. Judging from the affinity and the binding capacity of anti-human TSH autoantibodies, two classes of binding sites appear to exist, although the possibility of non-specific binding and/or non equilibrium effects cannot be ruled out. In patients 2 and 15, despite the high binding of 125I-hTSH to the serum components, large amounts of unlabeled human TSH were required to partially displace it, raising the possibility that serum components other than autoantibodies are involved to form macro-TSH.
The causes developing anti-human TSH autoantibodies are currently unknown. None of our patients had received injections of human thyrotropin-α or hCG, which had been reported in a previous study (20). Thus, these antibodies must be anti-human TSH autoantibodies that naturally developed in these patients, as in previous reports (21,22). Because serum samples examined in this study were those that clinicians ordered for thyroid tests in the first place, it is natural that there were several patients with thyroid autoimmune disorders among the 16 patients with macro-TSH. It is interesting, however, that there were patients with other autoimmune diseases such as SLE, systemic sclerosis, and rheumatoid arthritis, and some were being treated with steroids or interferon. It is therefore suggested that in some cases, systemic immune disorders might be involved in the development of anti-human TSH autoantibodies. It is also of note that patients with macro-TSH except patients 14 and 16 were middle-aged or elderly. Ageing may alter the antigenicity of TSH or break down immune tolerance, facilitating the production of autoantibodies to TSH.
Another interesting question is the length of time that macro-TSH persists in the blood. In this study, in 11/13 patients with macro-TSH, the complex was still present one to four years after the first evaluation. The relationship between serum macro-TSH ratios and TSH concentrations is difficult to interpret because eight patients were receiving thyroid hormone at the time of the second evaluation. In addition, the proportion of macro-TSH in sera is dependent on the binding characteristics of the anti-TSH autoantibodies and serum TSH levels. TSH that has bound weakly to anti-human TSH autoantibodies may dissociate during gel filtration chromatography. Thus, the proportion of high-molecular-weight TSH on gel filtration chromatography may not quantitatively reflect the level of anti-TSH autoantibodies. Nevertheless, the finding that serum TSH levels normalized in two patients whose macro-TSH disappeared supports the assumption that macro-TSH is a cause of hyperthyrotropinemia.
It is concluded that anti-human TSH autoantibodies are a major cause of macro-TSH and that macro-TSH tends to persist for a long time.
Footnotes
Acknowledgments
We thank Keiko Yamagami, Hidetoshi Ikeda, Fumio Otsuka, Kishio Toma, and Selin Elmaogulları for supplying clinical samples. We also thank Geoff Gillespie for his assistance in preparing this manuscript.
This work was supported by grants from the Ministry of Culture, Sports, Science, and Technology of Japan; from the Ministry of Health, Labor, and Welfare of Japan; and from Ritsumeikan University.
Author Disclosure Statement
No competing financial interests exist.