
Abstract
Background:
The intercellular adhesion molecule 1 (ICAM1) gene is not expressed in normal thyroid tissue but was shown to be expressed in the thyroid tissue of autoimmune thyroiditis (AIT) patients.
Methods:
This study aimed to explore whether the DNA methylation and hydroxymethylation status of the ICAM1 promoter are aberrantly altered in the thyroid cells of AIT patients and whether this change is associated with dysfunctional expression of ICAM1. A total of 35 AIT patients and 35 sex- and age-matched controls were studied. After the isolation of thyrocytes via density-gradient centrifugation, ICAM1 mRNA expression was measured using real-time PCR. The DNA methylation and hydroxymethylation status were assessed using quantitative PCR following T4 β-glucosyltransferase treatment and MspI/HpaII cleavage at −937 bp, −701 bp, −226 bp, and −65 bp upstream of the transcription start site (TSS). The DNA methylation level was verified via pyrosequencing.
Results:
The AIT group showed increased DNA hydroxymethylation at −937 bp and −226 bp and decreased methylation at −937 bp, −701 bp, and −226 bp upstream of the TSS. Pyrosequencing also revealed DNA hypomethylation at −708 bp, −692 bp, −690 bp, and −688 bp upstream of the TSS. The DNA methylation status at −708 bp, −692 bp, and −226 bp upstream of the TSS was negatively associated with ICAM1 mRNA expression.
Conclusion:
In summary, we identified aberrant DNA methylation and hydroxymethylation of the ICAM1 gene promoter in the thyrocytes of AIT patients. This aberrant epigenetic modification is associated with increased expression of the ICAM1 gene.
Introduction
A
DNA methylation regulates tissue-specific gene expression patterns by silencing the expression of function-irrelevant genes. DNA hydroxymethylation is a recently discovered type of DNA epigenetic modification. In addition to passive demethylation during DNA duplication, hydroxymethylation is a new mechanism of active demethylation that converts 5-methylcytosine (5-mc) to 5-hydroxymethylcytosine (5-hmc), followed by conversion to unmethylated cytosine by the ten-eleven-translocase (TET) family (10). Hydroxymethylation was initially found to be concentrated in embryonic cells and neurons and was subsequently shown to be distributed among all cell types and to be involved in various biological processes (11,12). In contrast to DNA methylation, promoter hydroxymethylation is related to transcriptional activation (13). Notably, DNA methylation and hydroxymethylation cannot be discriminated through traditional bisulfite sequencing, except via oxidation or glycosylation (14,15). Aberrant changes in methylation and hydroxymethylation have been observed in multiple diseases, including various cancers and autoimmune disorders (16,17).
This study aimed to explore the DNA methylation and hydroxymethylation status of the ICAM1 promoter. Polymorphisms of DNA methyltransferase genes and the whole-genome DNA methylation pattern of the CD4+ and CD8+ T cells of autoimmune thyroid disease patients have been studied previously (18,19). The DNA methylation status of the ICAM1 gene in CD4+ T cells was also shown to be associated with GD in genomic studies. However, the expression and DNA methylation patterns are tissue specific, and epigenetic modification of the target organ in thyroid disease has not yet been elucidated. This study analyzed the DNA modification of a specific gene in thyrocytes and focused on AIT patients for the first time.
Methods and Materials
Subjects
The study was approved by the Ethics Committee of the First Affiliated Hospital of China Medical University. The subjects underwent surgery for thyroid adenomas or nodules, or for a partial thyroidectomy to relieve tracheal compression caused by goiter in Hashimoto thyroiditis. Written, informed consent was obtained before surgery. Serum samples were also obtained before surgery to test thyroid function and detect autoantibodies, which were assessed using an electrochemiluminescence method (Architect i2000SR, Abbott Laboratories). Thyroid tissue was obtained through surgical excision and immediately placed on ice. All the patients included in the case group were females who showed positivity for at least one thyroid autoantibody (TPOAb or TgAb) and received a pathologic diagnosis of Hashimoto disease. Age- and sex-matched controls negative for the presence of autoantibodies were included. Thyroid tissue for the control group was obtained from para-adenoma tissue. All of the subjects were euthyroid and not exposed to radioactive iodine, antithyroid drugs, thyroid hormone replacement, and they did not have systemic autoimmune disease or any severe illnesses. Subjects with malignant nodules were excluded. Subjects who had positive thyroid autoantibodies but were not diagnosed with Hashimoto thyroiditis in surgical pathology were also excluded. The baseline clinical characteristics are summarized in Table 1.
Method: electrochemiluminescence method (Architect i2000SR, Abbott Laboratories, Chicago, IL).
p-Value <0.05 was considered to represent a statistically significant difference.
Isolation of thyrocytes
Thyrocytes were isolated via Percoll density-gradient centrifugation as previously described (20) with minor modifications. Fresh thyroid tissue obtained during surgery was minced into 1 × 1 × 1 mm pieces and incubated with 1.25 mg/mL collagenase IV (Sigma, C1889) in Hank's solution at 37°C. The digested materials were filtered through a nylon mesh with a 75 μm pore size every 30 minutes, and undigested fragments were collected to repeat the digestion procedure. The filtrate was centrifuged at 100 g for 5 minutes at 4°C and resuspended in 0.25% Trypsin-EDTA. A single-cell suspension was obtained via incubation at 37°C for 10 min. Enzyme activity was stopped by the addition of RPMI-1640 medium containing fetal bovine serum. The single-cell suspension was then gently transferred to a three-layer discontinuous Percoll gradient. The densities were 1.030 g/mL, 1.061 g/mL, and 1.077 g/mL from top to bottom, which corresponded to 20%, 45%, and 60% Percoll, respectively. After centrifugation at 400 g for 25 minutes at 4°C, thyrocytes sedimented between the first and second layers. The purity of the thyrocytes was tested via flow cytometry by staining for thyroglobulin. Thyrocyte preparations with a purity greater than 90% were used for experiments. The thyroglobulin antibody was purchased from Abcam (cat. ab76765). Subjects with insufficient tissue for thyrocyte isolation or a thyroid cell purity of less than 90% were excluded.
RNA and DNA preparation
DNA was extracted via the phenol–chloroform method. RNA was extracted using the TRIzol reagent (Thermo, 15596-026). The concentration and 260/280 OD value were determined using a NanoDrop 2000. All of the 260/280 OD values for the DNA and RNA samples were between 1.7 and 1.9 or 1.8 and 2.0, respectively. Reverse transcription of RNA was performed using PrimeScript RT Master Mix (TAKARA, RR036). Genomic DNA and cDNA samples were stored at −20°C until analysis.
DNA treatment
The DNA methylation and hydroxymethylation status of the ICAM1 promoter was analyzed as described in the manufacturer's instructions for the EpiMark 5-hmC and 5-mC Analysis Kit (E3317, NEB). Briefly, the genomic DNA was divided into two equal aliquots, one of which was treated with T4 β-glucosyltransferase in 80 mM uridine diphosphate–glucose at 37°C for 18 hours, while the control was treated with ddH2O. Each of these aliquots was then divided into three equal aliquots and treated with MspI/HpaII or no enzyme at 37°C for 16 hours. The MspI enzyme can recognize and cleave all CCGG sequences except for glycosylated 5-hmc, while HpaII can recognize only unmodified cytosines.
Quantitative PCR
The primers for the detection of ICAM1 mRNA levels and 5-mc or 5-hmc levels in the ICAM1 promoter region were designed using Primer3 (
Pyrosequencing
To validate the DNA methylation status measured using the EpiMark Kit, bisulfite pyrosequencing was performed for 20 subjects randomly selected from each group. The primers were designed using PyroMark Assay Design 2.0 and are listed in Table 2. The genomic DNA was reacted with bisulfite and analyzed on a PyroMark Q96 ID (Qiagen). The results were evaluated with Pyro Q-CpG software.
Statistics
The distribution normality was tested via the Kolmogorv-Smirnov method. Age and serum thyrotropin, free thyroxine, free triiodothyronine, and ICAM1 mRNA expression levels were normally distributed. These data were analyzed with Student's t test and the results are expressed as mean ± standard error. The data on TPOAb and TgAb and the percentages of 5-mc and 5-hmc within the ICAM1 promoter failed the normality test. These variables were analyzed with the Mann–Whitney U test and the results are expressed as medians with interquartile ranges. Correlation analysis was carried out to examine the association between the ICAM1 mRNA level and the 5-mc or 5-hmc level. The analysis was conducted with the AIT group and controls. For the cases with undetectable levels of 5-hmc or 5-mc, the outliers were excluded. Statistical analysis was carried out using SPSS 21 software. p-Values less than 0.05 were regarded as statistically significant.
Results
ICAM1 mRNA expression levels
The ICAM1 mRNA level was calculated relative to that of GAPDH mRNA. As shown in Figure 1, the ICAM1 mRNA expression level in the AIT group was approximately 80% higher than that in the control group [0.053 ± 0.010 vs. 0.089 ± 0.015 (mean ± standard error), p = 0.046].
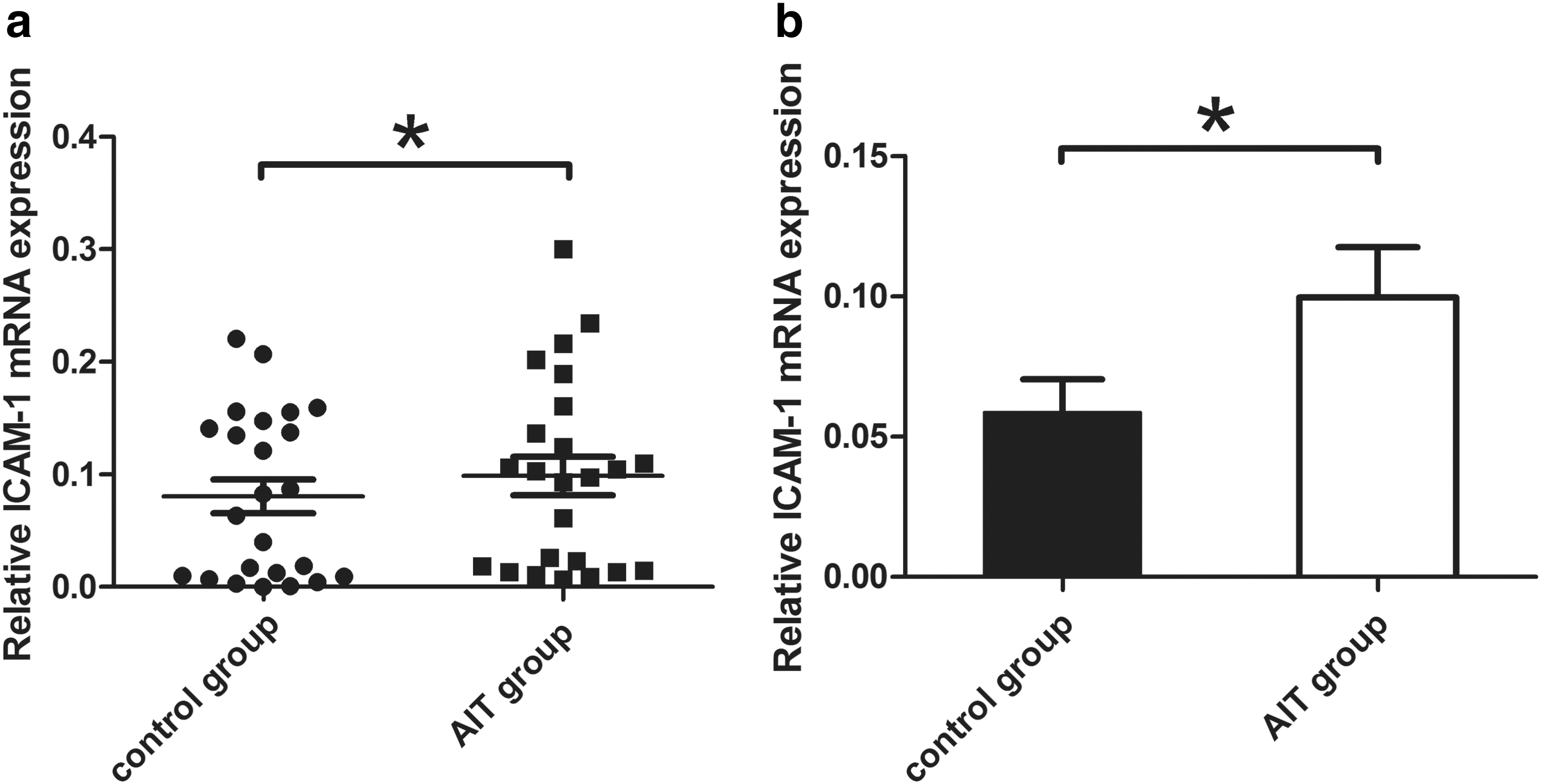
ICAM1 mRNA expression. (
Methylation and hydroxymethylation status of the ICAM1 promoter
The DNA methylation status and hydroxymethylation status at positions 1 (−937 bp), 2 (−701 bp), 3 (−226 bp), and 4 (−65 bp) upstream of the TSS were tested with an EpiMark Kit and expressed as the percentages of 5-mc and 5-hmc. The percentage of 5-hmc at positions 1 and 3 increased significantly in the AIT group (Fig. 2a, c). The median percentages of 5-hmc (with the interquartile range) at positions 1 and 3 were 6.41% (0, 33.61%) and 1.49% (0, 30.98%) in the AIT group and 0% (0, 0.48%) and 1.46% (0, 5.31%) in the control group, respectively (p < 0.05). The percentages of 5-hmc at the other positions did not show a statistically significant difference. The methylation levels at positions 1, 2, and 3 were decreased significantly in the AIT group (Fig. 2e–g). The median percentages of 5-mc (with the interquartile range) were 0% (0, 2.98%), 0% (0, 20.21%), and 14.89% (0, 34.51%) in the AIT group and 3.21% (0, 18.66%), 29.13% (0, 61.44%), and 56.74% (33.96%, 76.97%) in the control group, respectively (p < 0.05). The methylation levels at the other positions did not show a statistically significant difference. In the subjects selected for pyrosequencing verification, all of the tested positions (−708 bp, −692 bp, −690 bp, −688 bp, and −226 bp upstream of the TSS) showed an increased methylation level in the AIT group (Fig. 2i). This change was statistically significant. The relative positions of the methylated or hydroxymethylated CpG sites and transcription factor binding sites are shown in Figure 3, and the results are summarized in Table 3.
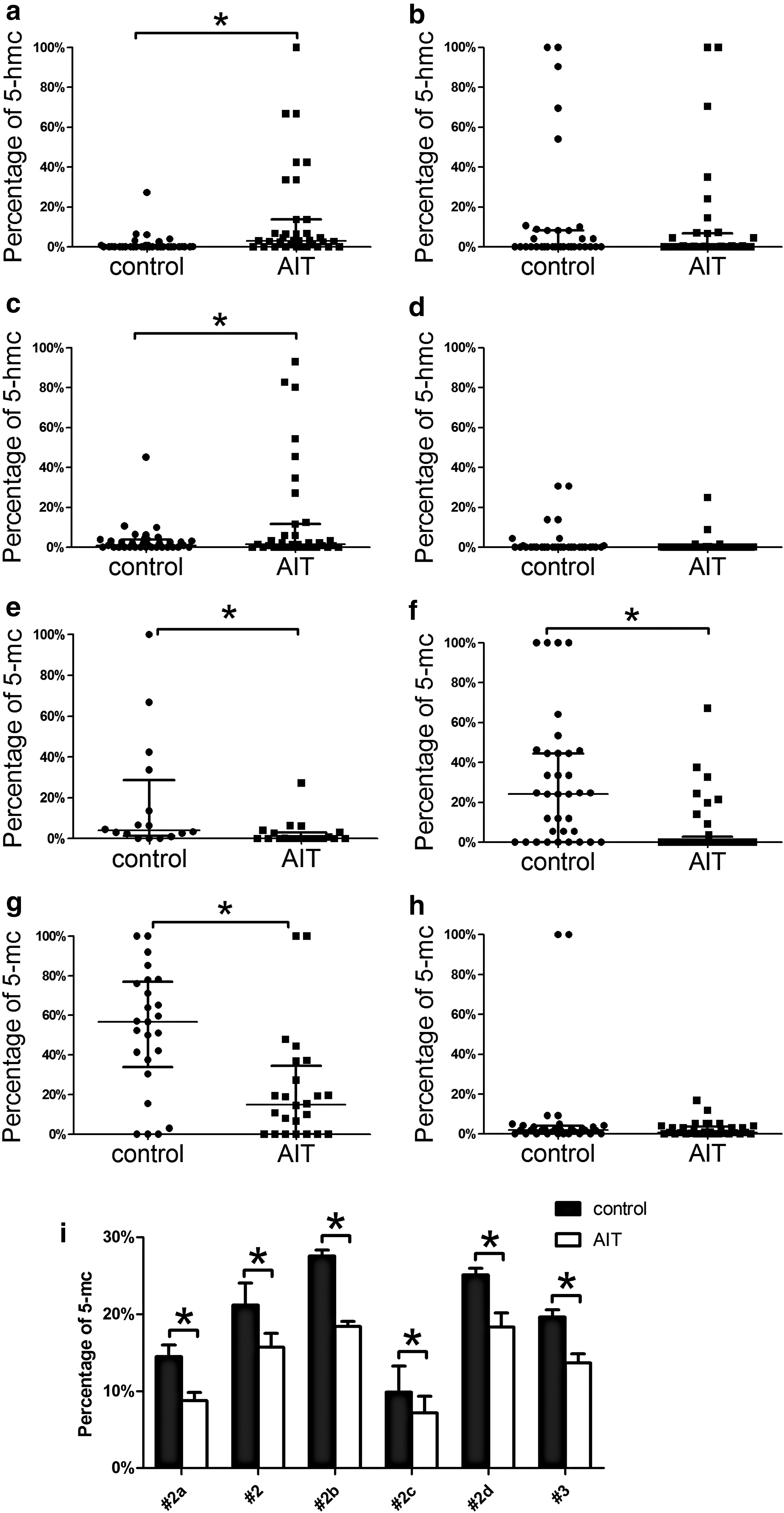
ICAM1 promoter methylation and hydroxymethylation status. (
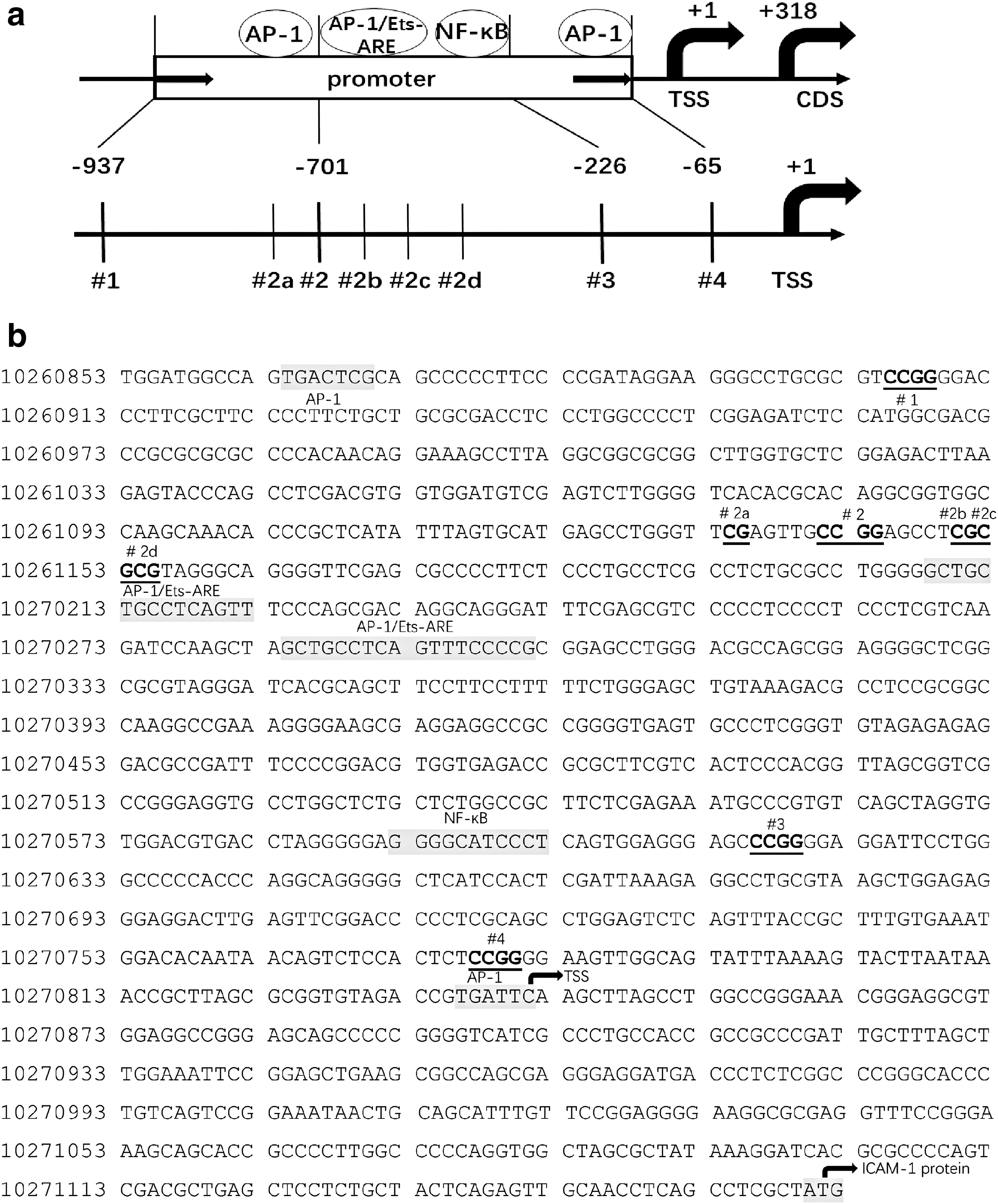
The tested sites in the ICAM1 promoter. (
#1 site is at −937bp upstream of TSS, #2 site is at −701bp upstream of TSS, #3 site is at −226bp upstream of TSS. #2a, #2b, #2c, #2d site are at −708 bp, −692 bp, −690 bp and −688 bp upstream of the TSS respectively.
p < 0.05 was considered to represent a statistically significant difference.
Correlation analysis was conducted between ICAM1 mRNA levels and 5-hmc or 5-mc percentages. The correlation analysis was conducted with the AIT and the control group. Since the 5-hmc and 5-mc percentages failed the normality distribution test, the Spearman correlation analysis for not normally distributed parameters was used. Cases with undetectable 5-hmc and 5-mc, were regarded as outliers and not included in the analysis. The methylation levels at positions 2a, 2b, and 3 showed moderate negative association with ICAM1 mRNA expression. The corresponding Spearman's correlation coefficients were r = −0.445 (p = 0.015, n = 40), r = −0.381 (p = 0.042, n = 40) and r = −0.401 (p = 0.031, n = 40), respectively.
Discussion
In this study, we found an abnormal decrease in DNA methylation and an increase in DNA hydroxymethylation at the ICAM1 promoter; these changes were associated with ICAM1 mRNA expression. The CpG sites that we tested in the ICAM1 promoter, from −931 bp to −226 bp upstream of the TSS, showed hyperhydroxymethylation and hypomethylation. The loss of DNA methylation at these sites in the thyroid of AIT patients was demonstrated via a glycosylation and enzyme digestion–based method and pyrosequencing. Enzymatic digestion after glycosylation is capable of discriminating 5-hmc from 5-mc. Our findings provide the first evidence regarding the DNA hydroxymethylation status of AIT patients, specifically showing that the level of 5-hmc modification at −921 bp and −226 bp upstream of the TSS is elevated. The DNA methylation observed at −708 bp, −692 bp and −226 bp upstream of the TSS was decreased and showed a correlation with ICAM1 mRNA levels. However, the hydroxymethylation status did not show any association with mRNA expression. The reason for this lack of association may lie in the fact that hydroxymethylation is a relatively temporary modification of cytosine. Additionally, the hyperhydroxymethylation did not interfere with the pyrosequencing results showing hypomethylation at −931 bp and −226 bp upstream of the TSS. The lack of abundance of 5-hmc may explain why the percentage of 5-hmc at the ICAM1 promoter is generally less than 10% and is significantly lower than that of 5-mc in a given patient.
ICAM1 is expressed on the surface of immunological cells and mediates homing and trafficking of these cells to target organs. It also plays an important role in cell interactions during antigen presentation or cell signaling transduction. In physiological conditions, ICAM1 is expressed at a low concentration on thyrocyte membranes and regulates the localization of immunological cells in thyroid tissue. ICAM1 expression is upregulated in thyrocytes of AIT patients (2). It acts as the binding site for inflammatory cells and enhances their recruitment to the thyroid at an early stage of the disease. ICAM1 gene expression can be increased by IFN-γ, excessive iodine and other environmental factors (8,21,22). Recently, several studies have found that increased soluble ICAM1 (sICAM1) in circulation is a hallmark of Hashimoto thyroiditis and GD (23 –25). However, the mechanism of ICAM1 overexpression in thyrocytes has not been elucidated.
Epigenetic regulation links environmental factors and gene expression, which could be one of the mechanisms involved in enhancing thyroiditis. The loss of promoter hypermethylation could activate previously silenced immune-related genes in thyrocytes. The pathways for DNA demethylation include passive demethylation during replication and active demethylation via oxidation by the TET family. A recent study demonstrated that TET2 can induce hypomethylation in the ICAM1 promoter and enhance ICAM1 gene expression in hepatocytes (26,27). We identified DNA hyperhydroxymethylation and hypomethylation in the ICAM1 promoter in AIT thyrocytes. Since DNA demethylation always leads to transcriptional activation (28), we hypothesized that the hypermethylated cytosine site is then hydroxymethylated by TET and converted to demethylated cytosine, leading to ICAM1 gene expression.
To the best of our knowledge, this is the first report on the DNA modification of a specific gene in thyrocytes from AIT patients. The involvement of epigenetic mechanisms in autoimmune thyroid disease has been reported by previous studies. In addition to the studies concentrating on DNA methylation, Stefan et al. (29,30) discovered that the transcription factor binding motifs in the promoter regions of the thyroglobulin and thyroid stimulating hormone receptor genes were enriched with mono- and trimethylated Lys-4 residues of histone H3 (H3K4me1), which is a type of active chromatin modification that enables transcription factor binding and gene expression. In addition, genomic H3K4me1 enrichment can be enhanced after IFN-γ treatment. Similar studies have identified an association of miRNA expression with autoimmune thyroid disease (31 –34). The data reported here demonstrate DNA modification through hydroxymethylation of the ICAM1 gene for the first time in AIT research. However, there are some limitations in our study. First, this is a cross-sectional observation. Correlation analysis indicate an association between the DNA methylation and mRNA expression of ICAM1, but this is not a strong evidence for a causal relationship. Second, our study focused on the DNA hydroxymethylation and methylation status of one specific gene. However, the biological behavior of a specific type of cell is regulated by the epigenetic modification and gene expression at the whole-genome level. Further studies are needed to elucidate the DNA modification pattern of the thyroid in AIT patients at the genomic scale.
In summary, our findings show that hydroxymethylation and methylation of the ICAM1 promoter was aberrantly changed and the methylation status is associated with expression of ICAM1 mRNA. Our results highlight the importance of studying epigenetics to elucidate the mechanism of environmental influence in disease onset. Further studies are needed to determine how risk factors affect the disease process of AIT at the genomic scale.
Footnotes
Acknowledgments
This work was supported by the National Natural and Science Foundation of China (grant number 81430018). We thank all the volunteers for their participation in this study and the donation of samples. We also thank Yushu Li, Haixia Guan, and Jing Li, physicians from the Department of Endocrinology and Metabolism of the First Affiliated Hospital of China Medical University, for their comments and advice.
Author Disclosure Statement
No competing financial interests exist.