
Abstract
Background:
Hashimoto's thyroiditis (HT) is considered to be the most common autoimmune disease. It is currently accepted that genetic susceptibility, environmental factors, and immune disorders contribute to its development. With regard to nutritional factors, evidence implicates high iodine intake and deficiencies of selenium and iron with a potential relevance of vitamin D status. To elucidate the role of nutritional factors in the risk, pathogenesis, and treatment of HT, PubMed and the Cochrane Library were searched for publications on iodine, iron, selenium, and vitamin D and risk/treatment of HT.
Summary:
Chronic exposure to excess iodine intake induces autoimmune thyroiditis, partly because highly iodinated thyroglobulin (Tg) is more immunogenic. Recent introduction of universal salt iodization can have a similar, though transient, effect. Selenoproteins are essential to thyroid action. In particular, the glutathione peroxidases protect the thyroid by removing excessive hydrogen peroxide produced for Tg iodination. Genetic data implicate the anti-inflammatory selenoprotein S in HT risk. There is evidence from observational studies and randomized controlled trials that selenium/selenoproteins can reduce thyroid peroxidase (TPO)-antibody titers, hypothyroidism, and postpartum thyroiditis. Iron deficiency impairs thyroid metabolism. TPO, the enzyme responsible for the production of thyroid hormones, is a heme (iron-containing) enzyme which becomes active at the apical surface of thyrocytes only after binding heme. HT patients are frequently iron deficient, since autoimmune gastritis, which impairs iron absorption, is a common co-morbidity. Treatment of anemic women with impaired thyroid function with iron improves thyroid-hormone concentrations, while thyroxine and iron together are more effective in improving iron status. Lower vitamin D status has been found in HT patients than in controls, and inverse relationships of serum vitamin D with TPO/Tg antibodies have been reported. However, other data and the lack of trial evidence suggest that low vitamin D status is more likely the result of autoimmune disease processes that include vitamin D receptor dysfunction.
Conclusions:
Clinicians should check patients' iron (particularly in menstruating women) and vitamin D status to correct any deficiency. Adequate selenium intake is vital in areas of iodine deficiency/excess, and in regions of low selenium intake a supplement of 50–100 μg/day of selenium may be appropriate.
Introduction
H
Although discovered a century ago, the pathogenesis of HT remains unclear. It is currently accepted that the complex interactions of genetic susceptibility, environmental factors, and immune disorders contribute to its development (4). Family and twin studies have confirmed a significant genetic influence on HT susceptibility (4), however, the concordance rate for overt Hashimoto's hypothyroidism found in Danish monozygotic twins was 55%, indicating an almost equally important role of environmental factors in the disease pathogenesis (5). A probable mechanistic model is that in genetically susceptible individuals, several environmental factors may trigger thyroid autoimmunity by increasing the immunogenicity of thyroid autoantigens, enhancing antigen presentation in the thyroid and reducing self-tolerance (4). Consequently, various pro-inflammatory cytokines are produced by immune and thyroid cells, resulting in predominantly Th1 and Th17 responses with an increased Th1/Th2 ratio (4). Meanwhile, increased production of pro-apoptotic cytokines leads to thyrocyte apoptosis and, finally, thyroid destruction (4). In addition, a decreased number or impaired function of regulatory T cells (Tregs), which are pivotal for maintaining peripheral tolerance and suppressing excessive immune response, has been recognized to play an important role in the pathogenesis of HT (6).
Regarding environmental factors, evidence implicates high iodine intake, selenium deficiency, infection, and certain drugs and chemicals in risk, while smoking and moderate alcohol consumption have a protective effect (7,8). Studies have also suggested a role for other common micronutrients, most notably iron and vitamin D, on HT risk (8). Although there is some evidence of an effect of vitamin A and zinc on thyroid metabolism, only sparse data are available on their relationship with HT. Hence, this review concentrates on iodine, selenium, iron, and vitamin D.
Methods
PubMed and the Cochrane Library were searched for publications up to October 2016 using the search terms “Hashimoto's thyroiditis” OR “autoimmune thyroiditis” OR “autoimmune thyroid disease” in combination with “iodine,” “selenium,” “iron,” “vitamin D,” and “nutrition OR diet.” Articles were filtered by relevance of title, abstract, and finally the full text. Relevant conclusions or results were extracted from each article.
Review of Iodine
The micronutrient iodine is an essential constituent of the thyroid hormones that play a pivotal role in growth, development, and metabolism (9). The biologically active form, triiodothyronine (T3), stimulates oxygen consumption, controls basal metabolic rate and thermogenesis, regulates the expression of numerous target genes that affect protein synthesis either positively or negatively, regulates cell activity and growth, and is essential for brain development and function, particularly in fetal life (9).
Role of iodine in the thyroid
Iodide is taken up from the blood by thyroid epithelial cells. At the apical surface of the thyrocyte in the follicular lumen, in the presence of hydrogen peroxide (H2O2), the enzyme thyroid peroxidase (TPO) iodinates tyrosine molecules on the surface of thyroglobulin (Tg), a large glycoprotein synthesized by thyroid epithelial cells (9). The products are mono- and di-iodotyrosines (9). The coupling of these products, still attached to Tg, again catalyzed by TPO in the presence of H2O2, forms the thyroid hormones T3 and thyroxine (T4) (9). The iodinated Tg molecule then enters the thyrocyte and is digested, releasing T4 and T3 into the circulation (9).
Evidence for a relationship between iodine intake/status and HT risk
Iodine intake has a key influence on the spectrum of thyroid disorders in populations (2,10). Severe iodine deficiency causes goiter and hypothyroidism due to reduced production of thyroid hormone, whereas chronic mild-to-moderate iodine deficiency may increase the prevalence of toxic nodular goiter and hyperthyroidism (2,10). Excess iodine intake or a rise in intake after initiating iodine fortification of an iodine-deficient population are associated with an increased risk of subclinical hypothyroidism and thyroid autoimmunity (2,10).
Iodine deficiency has long been an important public health issue worldwide, causing multiple detrimental consequences (11). Since the implementation of iodine fortification, particularly universal salt iodization, substantial progress has been made to prevent and control iodine-deficiency disorders. However, excessive iodine intake, which can also result from the implementation of universal salt iodization programs at too high a level of supplementation, increases the risk of several thyroid disorders, including autoimmune thyroiditis (12). Numerous epidemiological studies have associated high iodine intake with increased prevalence of autoimmune thyroiditis in populations (12 –16). However, the optimization of nutritional iodine intake ultimately results in decreased prevalence of autoimmune thyroiditis (17). The association between high iodine exposure and increased incidence of autoimmune thyroiditis has also been demonstrated in a variety of animal models with genetic susceptibility (18 –20). The ingestion of excess iodide through drinking water by the autoimmune thyroiditis–prone NOD.H2h4 mouse significantly enhances and accelerates the incidence and severity of autoimmune thyroiditis in a dose-dependent manner (21).
The mechanisms by which increased chronic intake of dietary iodine induces autoimmune thyroiditis are unclear. It is thought that in genetically susceptible individuals, chronic high iodine exposure may: (i) increase the immunogenicity of Tg, (ii) induce auto-antigen presentation activity of thyrocytes and dentritic cells, (iii) impair peripheral tolerance by inhibiting Tregs, (iv) cause oxidative stress leading to thyroid tissue injury, and (v) activate autoreactive T cells, which increases cytokine secretion and eventually triggers apoptosis signaling pathways, leading to thyroid destruction (12,21,22).
Recommendations on iodine intake
The level of dietary iodine intake has a very significant effect on the pattern of thyroid disorders in populations. While iodine deficiency is recognized to have multiple adverse effects on the thyroid, with regard to autoimmune thyroiditis/HT, there is more evidence for an association with iodine excess, especially in genetically susceptible individuals (12,16,22). To avoid an increased risk of HT, it is therefore important to ensure, as far as possible, that iodine intake falls within the relatively narrow range of the recommended levels (23), (see Table 1 [24–26]). On a population basis, this would be represented by a median urinary iodine concentration in adults of 100–200 μg/L (26). Authorities introducing iodine fortification of the food supply in a country (e.g., universal salt iodization) need to ensure that such fortification is introduced very cautiously; Denmark provides an excellent example of how this can be done (27).
AI, adequate intake; EFSA, European Food Safety Authority; RDA, recommended dietary allowance; RNI, recommended nutrient intake.
Review of Selenium
Selenium is a trace mineral essential for human health (28). As selenocysteine (an analogue of cysteine), it is incorporated into 25 human selenoproteins that have a wide range of functions from antioxidant and anti-inflammatory agents to the production of active thyroid hormone (28,29). An indication of the importance of selenium to the thyroid is the fact that it contains the highest concentration of selenium in the human body and is able to retain that selenium under conditions of severe deficiency, which causes its loss from many other tissues (30).
Role of selenium in the thyroid: selenoproteins
A number of selenoproteins are expressed in thyrocytes, for example the deiodinase isozymes (Dio1, Dio2, although not Dio3), members of the glutathione peroxidase family (GPx1, GPx3, GPx4), the thioredoxin reductases (TxnRd1 and TxnRd2), selenoprotein 15, selenoprotein P (SELENOP), and selenoproteins M and S (SELENOM, SELENOS) (31). Those below play particularly important roles.
The deiodinases
DIO1 and DIO2 can activate T4 by transforming it into T3 by removal of the 5′-iodine, while DIO1 and DIO3 can prevent T4 from being activated by converting it to the inactive reverse T3 (32). DIO3 can also inactivate T3 by 5-deiodination to 3,5-diiodo-L-thyronine (T2). Outside the thyroid, DIO2 is largely responsible for local conversion of T4 to T3 in target tissues (29). DIO3 is found in fetal tissue, the placenta, and central nervous system where it protects sensitive cells from thyrotoxic concentrations of active T3 (29,33).
The glutathione peroxidases
GPx3, normally found in the plasma, is also secreted at the apical side of the thyrocyte membrane where it degrades excess H2O2 that has not been used by TPO for the iodination of tyrosyl residues of Tg or for iodotyrosine coupling (see Fig. 1) (34). GPx1 protects the intracellular compartment from excessive H2O2 that may diffuse into the thyrocytes, while GPx4 can remove excessive lipid hydroperoxides in the mitochondria (33,34).
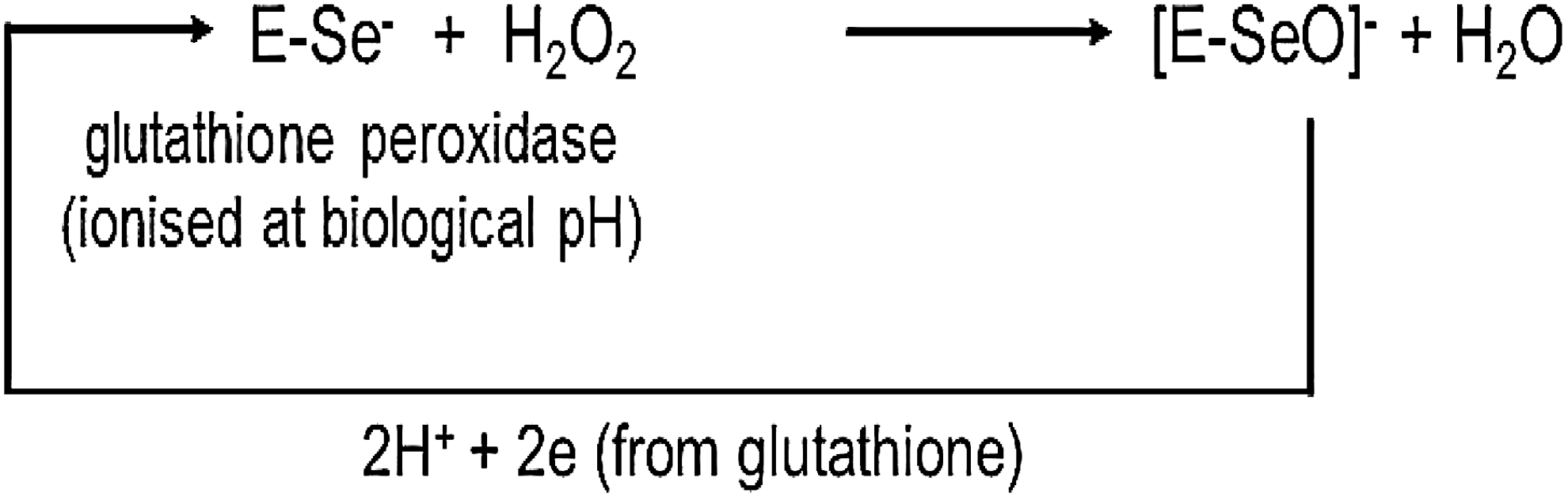
Glutathione peroxidases catalyze the removal of H2O2 (and lipid hydroperoxides) converting it to harmless water, thus protecting the thyroid from excessive exposure to H2O2.
Selenoprotein S
SELENOS is involved in the control of the inflammatory response in the endoplasmic reticulum (ER) (35). In a Portuguese study, the SELENOS −105G/A promoter polymorphism (rs28665122) was strongly associated with circulating levels of cytokines such as interleukin 1 beta (IL-1β), IL-6, and tumor necrosis factor alpha (36), which are known to be involved in HT pathogenesis. A-allele carriers of this polymorphism were more than twice as likely as GG homozygotes to have HT; in male carriers, the risk was fourfold higher (36).
Evidence for a relationship between selenium intake/status and HT risk/treatment
Deficiency of selenium has been associated with a number of adverse thyroid conditions, including hypothyroidism, subclinical hypothyroidism, and autoimmune thyroiditis/HT (37); an enlarged thyroid (37 –39); thyroid cancer (31,40,41); and Graves' disease (42).
A recent study, very relevant to the effect of selenium status on HT risk, was carried out in two counties of Shaanxi Province, China, which had high genetic, environmental, and life-style similarities and comparable iodine status but very different selenium status—adequate and low (37). Participants (n = 6152) completed demographic and dietary questionnaires, underwent physical and thyroid ultrasound examinations, and had serum samples analyzed for thyroid function parameters and selenium concentration. Selenium concentrations differed almost twofold (median [interquartile range] = 103.6 [79.7–135.9] vs. 57.4 [39.4–82.1] μg/L; p = 0.001) between participants from the two counties. After adjustment for potential confounders, the prevalence of pathological thyroid conditions was significantly lower in the adequate selenium than in the low selenium county (18.0% vs. 30.5%; p < 0.001). Higher serum selenium was associated with lower odds of autoimmune thyroiditis (odds ratio [OR] = 0.47 [confidence interval (CI) 0.35–0.65]), hypothyroidism (OR = 0.75 [CI 0.63–0.90]), subclinical hypothyroidism (OR = 0.68 [CI 0.58–0.93]), and enlarged thyroid (OR = 0.75 [CI 0.59–0.97]) (37). Both these counties had an iodine intake that was more than adequate (26,37,43), which may have accounted to some extent for the high level of thyroid disease prevalence (15,44). This study suggests that in such a situation, having an adequate selenium status may be protective.
Several trials of selenium supplementation have been carried out in both autoimmune thyroiditis (HT) and mild Graves' orbitopathy. In a large, multicenter, randomized, controlled trial (RCT) with selenium, patients with mild Graves' orbitopathy significantly improved (45). There have been a number of systematic reviews/meta-analyses of controlled trials of selenium treatment in patients with autoimmune thyroiditis/HT (33,46 –48). The most recent included 16 trials in a meta-analysis that found that selenium supplementation reduced serum TPOAb levels after 3, 6, and 12 months in a population with chronic autoimmune thyroiditis treated with levothyroxine, and after three months in an untreated population (46). Some of these studies also saw a reduction in TgAb titers at 12 months, an improvement in thyroid echogenicity, and an increase in subjective well-being. However, the methodology of many of the studies was flawed; for example, they were underpowered, not double-blinded, and not placebo-controlled, and disparities in iodine intake were not considered (46,47,49). The beneficial effect in some studies and not in others cannot easily be explained on the basis of baseline selenium status, stage of disease, baseline TPOAb titers, or form or dose of selenium used (8). Hence, well-designed, properly powered RCTs of selenium in the treatment of autoimmune thyroiditis/HT are still needed before we can confidently recommend selenium supplementation in HT patients.
Despite these caveats, there is a rationale for a beneficial effect of selenium—probably through its role within selenoproteins—on autoimmune thyroid disease/HT: (i) selenium, as the glutathione peroxidases and the thioredoxin reductases has an antioxidant, protective function (28,50); (ii) selenium can upregulate regulatory T cells, resulting in increased immune tolerance (in an autoimmune thyroiditis model system) (51); (iii) selenium has anti-inflammatory effects (35,52,53); and (iv) selenium may suppress the expression of HLA-DR molecules on thyrocytes, reducing the development of thyroid autoimmunity (50,54).
Evidence for a relationship between intake/status of selenium and autoimmune thyroid disease in pregnancy and the postpartum period
Several clinicopathologic variants are now thought to be included under the term HT, including postpartum thyroiditis (1). Pregnant women positive for TPOAbs are likely to develop hypothyroxinemia during pregnancy and postpartum thyroiditis in the year after delivery (55). Up to 50% of TPOAb-positive pregnant women develop postpartum thyroiditis of whom 20–40% subsequently become hypothyroid (55).
An RCT in TPOAb-positive women in Italy found that selenium supplementation reduced thyroid inflammatory activity and the risk of postpartum thyroid disease (56). During pregnancy and the postpartum period, 151 TPOAb-positive women were randomized to selenium (200 μg/day as selenomethionine) or placebo. TPOAbs fell significantly during gestation, but the reduction was significantly greater in the selenium-supplemented group (p = 0.01) and remained so in the postpartum period (p = 0.01; see Fig. 2). Importantly, there was a significant reduction in the incidence of postpartum thyroid disease and hypothyroidism in the selenium-supplemented group (28.6% vs. 48.6%, p < 0.01, and 11.7% vs. 20.3%, p < 0.01, respectively) (56). Furthermore, during treatment, women on selenium maintained the same level of ultrasound echogenicity, whereas in those on placebo, echogenicity significantly worsened. At the end of the postpartum period, grade 2–3 thyroiditis had developed in 44.3% of women on placebo but only in 27.3% of women on selenium (p < 0.01).
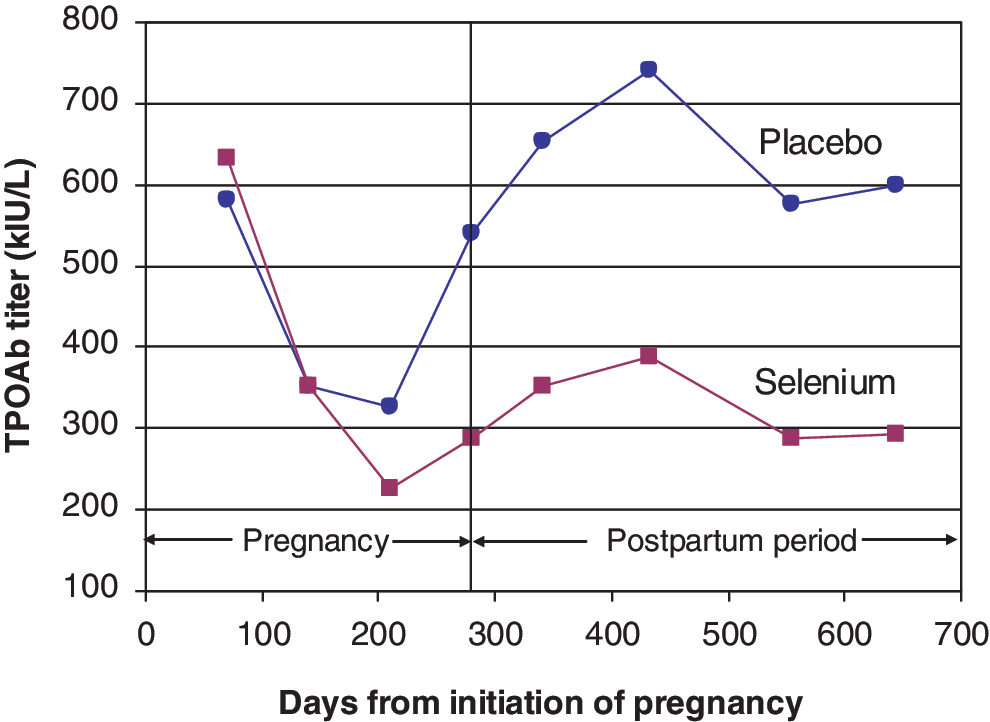
Selenium protects against postpartum autoimmune thyroid disease (adapted from Negro et al. [56]). Color images available online at
The only other RCT that investigated the effect of selenium supplementation on autoimmune thyroid disease in pregnancy found no difference in the magnitude of decrease between selenium and placebo groups (57). However, the trial was underpowered, the median baseline TPOAb concentrations in the women were much lower than those in the study by Negro et al. (56), and the selenium dose given was considerably less (i.e., 60 vs. 200 μg/day). There is clearly a need for a further, high-quality, adequately powered RCT in the TPOAb-positive pregnant population to see if the results of Negro et al. can be replicated (56).
Is selenium intake/status adequate?
The intake of selenium shows tremendous variability from one part of the world to another, with levels ranging from deficient (7 μg/day) to toxic (4990 μg/day) (28). Figure 3 shows the variability and gives an indication of the level of intake believed to be needed to optimize the activity of GPx3 (58), the main selenoenzyme responsible for removing excess H2O2 from the thyroid. This geographical variability in intake (and hence status) relates not only to the selenium content of the soil on which crops and fodder are grown but to many other factors that determine the availability of selenium to the food chain such as selenium speciation, soil pH, and organic-matter content. Mean intake is some 40 μg/day in Europe, and 93 μg/day for females and 134 μg/day for males in the United States (28). Recommended selenium intake varies by authority and averages 60 μg/day for men and 53 μg/day for women (59). Supplements of selenium contribute to intake and are quite commonly consumed, particularly in the United States, where some 50% of the population take dietary supplements (60).
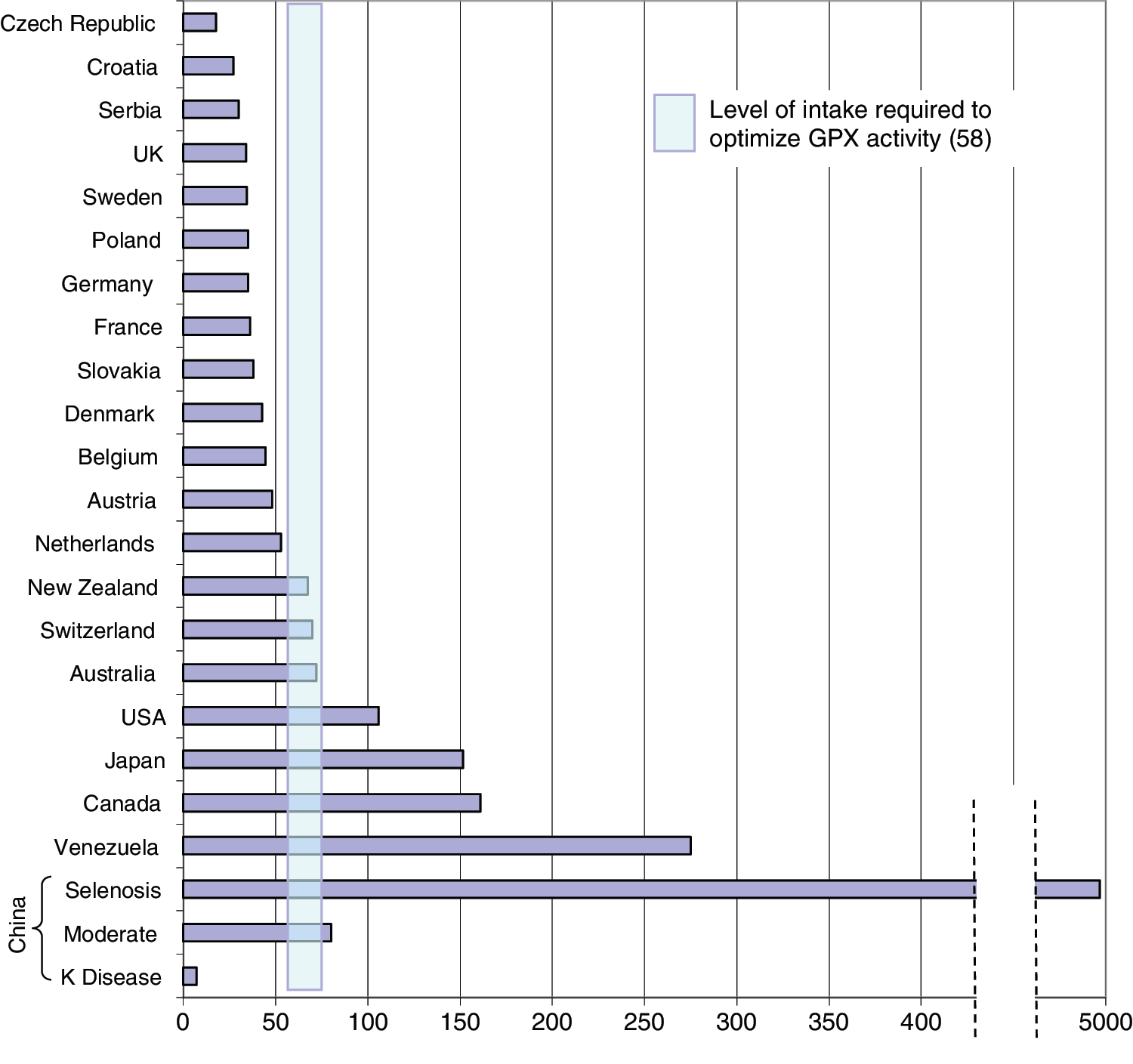
Mean selenium intake levels (μg/day) in different countries and the range of selenium intake (55–75 μg/day) believed to be required for optimal activity of plasma glutathione peroxidase (GPX3; adapted from Rayman [58]). Color images available online at
Food sources of selenium
Brazil nuts are the richest food source of selenium, though they are generally not a commonly eaten food. In any case, they cannot be recommended as a main selenium source, as the content is very variable, ranging from 0.03 to 512 mg/kg fresh weight, and they are high in barium (59). Organ meat and seafood are good sources, followed by muscle meats, cereals, and grains, though the selenium content of the latter varies widely (see Fig. 4) (28, 59). Thus, in the United States, grains such as wheat are excellent selenium sources and provide some 37% of dietary selenium (61), whereas in the United Kingdom they only provide 26% of selenium intake (62).
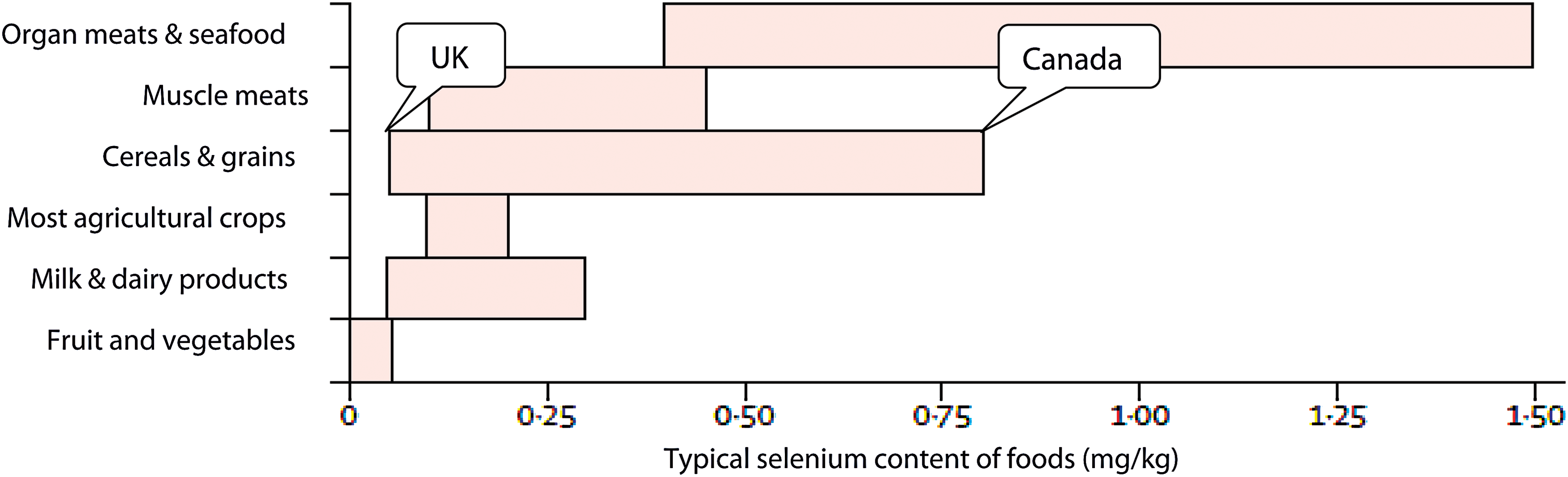
Typical selenium content of food sources, adapted from the World Health Organization. Selenium. A report of the International Programme on Chemical Safety. Environmental Health Criteria number 58. World Health Organization, Geneva, 1987 (reproduced from Rayman [28]). Color images available online at
Recommendations for selenium intake
Though evidence is lacking that selenium supplementation results in clinical improvement in autoimmune thyroiditis (other than in mild Graves' orbitopathy), it still makes sense to ensure that selenium intake is adequate, given the roles played by selenium/selenoproteins in human health (28) and particularly in the thyroid. Regions of deficient, more-than-adequate, or high iodine intake may have more need for selenium because of the capacity of selenoproteins to protect the thyroid from damage from H2O2, reactive oxygen species, and inflammation and to increase immune tolerance (see above). Hence, under these circumstances, clinicians need to be especially vigilant to ensure that selenium intake/status is adequate. Women are at greater risk of thyroid disorders and may thus have a higher requirement for additional selenium, particularly in pregnancy. In addition, geographical location will give a good indication of selenium adequacy or otherwise (see Fig. 3). It is also important to enquire into patients' dietary habits to see they eat foods that supply selenium (see above) (28). In China, for instance, selenium-enriched tea is an excellent selenium source (37) and is available in many areas.
If there appear to be few or no selenium-rich sources in a patient's diet, low-dose supplementation (50–100 μg/day) is suggested. Multivitamin/mineral tablets may contain 50 μg/day of selenium, an amount that will generally be adequate for women. A dose of 100 μg/day of selenium (as selenium yeast) given to someone in the United Kingdom will raise plasma selenium to around 140 μg/L, which is more than enough to optimize the synthesis of all the selenoproteins (63). Either selenium yeast (which behaves in the body like wheat selenium) or sodium selenite (the latter is not non-specifically incorporated into body proteins in place of methionine) is appropriate (59).
Even if a patient with HT is being treated with levothyroxine, one needs to be aware that some studies found that giving Se as well as levothyroxine resulted in a greater reduction in TPOAbs, inflammatory cytokines, and C-reactive protein (53,64).
It is also important to bear in mind that though selenium is essential, excessive intake of selenium is toxic, and supplements of selenium of 200 μg/day, generally considered to be quite safe, have been associated with toxic effects (alopecia, dermatitis, squamous cell carcinoma, type-2 diabetes mellitus) in North American men (65 –67), though these men had a higher selenium status than European men. As for many nutrients, there is a U-shaped relationship between selenium status and disease risk. Therefore, it is recommended to aim for an adequate intake that does not stray into levels associated with potential toxicity (28).
Review of Iron
Iron is an essential major mineral. Through its presence in hemoglobin, myoglobin, and many iron-containing enzymes, it is involved in a great number of metabolic processes in the body. These include oxygen transport and storage, DNA synthesis, ATP generation, oxidation-reduction reactions, electron transfer, and regulation of the cell cycle (68 –70). In healthy adults, iron metabolism is strictly regulated to maintain body iron content within a restricted range. This is because iron deficiency leads to decreased oxygen transport and impaired activity of iron-containing enzymes, and because iron excess may predispose to iron-overload diseases and cancer (68,69).
Role of iron in the thyroid
TPO, the enzyme required for the organification and coupling reactions in thyroid hormone synthesis (71,72), becomes active at the apical surface of thyrocytes only after it binds a prosthetic heme group (73). Hence, an adequate iron status is essential for the production of the thyroid hormones T3 and T4.
Evidence for a relationship between iron intake/status and HT risk/treatment
HT and iron deficiency
Studies have revealed that HT patients with subclinical hypothyroidism have lower serum iron concentrations and a higher prevalence of iron deficiency than healthy controls (74,75). As an organ-specific autoimmune disease, HT is frequently associated with other autoimmune disorders. Indeed, a considerable proportion of HT patients have celiac disease (76 –78) or autoimmune gastritis comorbidity (79 –82), and these comorbid conditions are regarded as the major cause of iron deficiency in HT patients. Iron-deficiency anemia is the most common extra-intestinal manifestation of celiac disease, which impairs iron absorption and leads to iron deficiency (83). Autoimmune gastritis is characterized by serum anti-parietal cell antibodies and anti-intrinsic factor antibodies. It can finally evolve to severe atrophic gastritis with subsequent hypochlorhydria and chronic iron deficiency (84). Because of the abnormal gastric secretion and low acidity, dietary non-heme iron cannot effectively be solubilized, leading to iron malabsorption (85).
It appears likely, however, that hypothyroidism per se, which is common in HT patients, impairs gastrointestinal iron absorption. Early experiments in hypothyroid rats showed diminished gastrointestinal iron absorption that was restored to normal on supplementation with T3 (86). In two studies in patients with coexisting iron-deficiency anemia and subclinical hypothyroidism, treatment with iron and T4 together was considerably more effective in improving iron status than treatment with iron alone (87,88).
Iron deficiency affects thyroid metabolism
Not only do HT patients have a higher prevalence of iron deficiency, but iron deficiency also impairs thyroid metabolism. It reduces thyroid hormone production by decreasing the activity of the iron-dependent enzyme TPO (71 –73). In rodent studies, iron deficiency, with or without anemia, decreased serum T4 and T3 concentrations, lowered 5′-deiodinase activity, and reduced the ability to thermoregulate in response to a cold environment (72,89 –91). Apart from the observed effect of iron deficiency on thyroid hormone production (72,73), it has been suggested that it may lead to functional hypothyroidism by altering the central regulation of the thyroid axis (90) and hampering the binding of T3 to hepatic nuclear receptors (92).
Human studies have provided equivocal outcomes. While a few studies found no significant association between serum thyroid hormone concentrations and iron status (93,94), others had different results. Lower serum T4 and/or T3 and higher thyrotropin (TSH) levels were reported in women with iron-deficiency anemia than in non-anemic controls, and iron supplementation partially normalized plasma thyroid hormone concentrations (95,96). A small Finnish study illustrates that low iron stores may contribute to symptom persistence in patients treated for hypothyroidism (97). Twenty-five women with persistent symptoms of hypothyroidism despite appropriate levothyroxine therapy became symptom-free when treated with oral iron supplements for 6–12 months. None of the women had anemia or red-cell indexes outside the reference range, though all had serum ferritin <60 mg/L (97). A study conducted in 4392 women of childbearing age indicated that iron deficiency was independently correlated with isolated hypothyroxinemia in both pregnant and nonpregnant women (98). Two cross-sectional studies showed significantly higher risks of goiter in children with low serum iron (99,100).
Recommendations for iron intake
As explained above, HT patients have a high prevalence of iron deficiency or low iron stores that may impair thyroid metabolism. Hence, routine screening of HT patients for iron deficiency is recommended. If either deficiency or a serum ferritin of <70 μg/L is found (97), comorbid celiac disease or autoimmune gastritis should be suspected as a potential cause and treated if necessary. If celiac disease is diagnosed, it should be formally documented. Hematological tests can be used to distinguish between iron-deficiency anemia that will respond to iron supplementation and the anemia of chronic disease that will not. Assuming the latter is not involved, supplementation to restore iron sufficiency should be instituted and will help to prevent the deleterious effects of iron deficiency on thyroid function (95, 96). There are alternative supplements to ferrous sulfate (e.g. ferrous bisglycinate) that may be better tolerated by the gastrointestinal tract (101,102).
Review of Vitamin D
Vitamin D is a steroid hormone precursor, pivotal for bone and mineral homeostasis that balances serum levels of calcium and phosphorus (103). It has two major forms of which vitamin D2 comes exclusively from diet and vitamin D3 is largely synthesized in the human skin through sunlight exposure. Despite the difference in side-chain structure, both are hydroxylated in the liver by 25-hydroxylase to 25-hydroxyvitamin D (25[OH]D, calcidiol), which is carried by vitamin D binding protein (VDBP), and is used as the circulating indicator of vitamin D status (104). In the classic pathway, 25(OH)D is then converted to 1α,25-dihydroxyvitamin D (1α,25[OH]2D, calcitriol) by a cytochrome P450 enzyme, 1-α-hydroxylase (CYP27B1) in the kidney. This is the hormonally active form and exerts its endocrine effects by binding to the vitamin D receptor (VDR) and regulating VDR-responsive genes (105). However, numerous recent studies have shown that many tissues have local 1-α-hydroxylase that can produce 1α-25(OH)2D that has both autocrine and paracrine effects (103,106). Moreover, the discovery of VDRs in >35 tissues unrelated to bone metabolism demonstrates the pleiotropic effects of vitamin D (107). In fact, many observational studies have demonstrated inverse correlations between circulating 25(OH)D concentrations and the risk of extra-skeletal diseases (108 –110).
Role of vitamin D in the thyroid
If vitamin D has a role in the thyroid, it is likely to be via its effect on the immune system and its role in dealing with infection. The potential for chronic infectious agents to be a causal factor for autoimmune disease has long been recognized (111). For instance, Epstein–Barr virus (EBV) is a ubiquitous herpes virus that is suspected to be involved in the pathogenesis of many autoimmune diseases (112). There is also some serological evidence of bacterial infection in patients with HT (113 –115).
Multiple in vitro studies have provided compelling evidence that 1α,25(OH)2D, acting through the VDR, induces innate antimicrobial activity by regulating the expression of antimicrobial peptides (cathelicidin hCAP18 and defensin beta 4) that are responsible for extensive antimicrobial action (116 –119) and the activation of antibacterial autophagy (120,121). While 1α,25(OH)2D is a promoter of innate immunity, it suppresses the adaptive immune response (116) by inhibiting the pro-inflammatory effects of Th1 and Th17 cells and enhancing the anti-inflammatory activities of Th2 and Treg cells (122 –124). Furthermore, 1α,25(OH)2D is believed to play a protective role against autoimmunity. On the one hand, it exerts special immunoregulatory and tolerogenic effects by hampering the maturation and autoantigen presentation of many dendritic-cell subsets (125). On the other hand, it increases the count of CD8+ T cells that are capable of controlling EBV infection and clearing EBV-infected autoreactive B cells (112).
Evidence for a relationship between intake/status of vitamin D and HT risk/treatment
Since excessive activation of Th1 and Th17 cells, as well as impaired function of Treg cells (6) and deficiency of CD8+ T cells (112), are implicated in the pathogenesis of HT, it is conceivable that vitamin D status may affect the development of this disorder.
Evidence from animal studies
In vivo studies showed that a low-dose combination of 1α,25(OH)2D3 and cyclosporine could effectively prevent the induction of experimental autoimmune thyroiditis (EAT) in an mouse model of thyroiditis similar to human HT (126). Injection of high-dose 1α,25(OH)2D3 showed a therapeutic effect in established EAT rat models by improving thyroid-gland structure and restoring Th1/Th2 cytokine equilibrium (127).
Evidence from human studies
Evidence from human studies is not strong. Almost all studies were cross-sectional in nature, and there are no RCTs of vitamin D and thyroid autoimmunity.
A number of case-control studies (128 –137) reported lower mean levels of 25(OH)D, as well as higher rates of vitamin D deficiency or insufficiency in HT patients than in healthy controls (see Table 2). Furthermore, subjects with vitamin D deficiency had a higher risk of developing HT than those with normal levels; for example, for every 5 nmol/L increase in plasma 25(OH)D concentration, a 1.62-times decrease in HT risk was found (129). Moreover, inverse relationships of serum 25(OH)D concentrations with TPOAb and TgAb titers in HT patients have been seen in a number of studies (130,132,133,135,136). With regard to thyroid function, data from a case-control study demonstrated that serum 25(OH)D status correlated inversely with TSH levels and positively with T3 levels in hypothyroid patients (134), while hypothyroidism at diagnosis was more prevalent in HT patients with serum 25(OH)D concentrations <10 ng/mL (25 nmol/L) than in those within the normal range (138). In a comparison of chronic and new-onset HT patients and healthy controls, a clear association was found between the severity of vitamin D deficiency and disease duration, as well as a positive correlation between serum 25(OH)D levels and thyroid-gland volume in the patients (133).
AITD, autoimmune thyroid disease; GD, Graves' disease; HT, Hashimoto's thyroiditis; TPOAb, thyroid peroxidase antibodies.
However, two studies had different findings. In a case-control study, serum 25(OH)D concentration was found to be no lower in HT patients than it was in controls (139). More significantly, a longitudinal study failed to show lower vitamin D levels in women who developed TPOAbs during follow-up than in those who remained thyroid-antibody negative, indicating a lack of relationship between early-stage thyroid autoimmunity and vitamin D insufficiency (140). Some of the divergence between study results might be attributable to differences in latitude, season, sunlight exposure, ethnicity, body mass index, assay methods, as well as inadequate matching between cases and controls with respect to confounding factors that can affect vitamin D levels (139,141,142).
Could the association between low vitamin D status and HT be a result of ill health?
However, a more likely scenario may be that low concentrations of 25(OH)D in HT patients are simply a result of disease. For instance, in HT, increased body-fat mass due to hypothyroidism as well as other comorbid autoimmune diseases may predispose to vitamin D deficiency (143). Low serum concentrations of 25(OH)D have been observed in many extra-skeletal diseases and may simply be a sign of ill-health (144). The validity of this hypothesis is strengthened by the fact that none of the numerous randomized trials carried out with vitamin D (though none was in HT) confirmed the health benefits of increased 25(OH)D, even when high doses of supplementation were given to participants with low vitamin D status prior to randomization (144,145).
Could the association between low vitamin D status and HT be a result of VDR dysfunction?
Another explanation for the low serum 25(OH)D observed in autoimmune diseases, including HT, is VDR dysfunction in phagocytes resulting from chronic infection with intracellular bacteria that dysregulate vitamin D metabolism (111). Because the VDR controls expression of the cathelicidin and beta-defensin antimicrobial peptides, dysregulation of the receptor greatly compromises the innate immune response (113). Bacterial-induced VDR dysfunction can explain the low concentrations of 25(OH)D and high concentrations of 1,25(OH)2D (111). Thus, in inflammatory conditions, unregulated extra-renal production of 1,25(OH)2D occurs and escapes breakdown by binding to the pregnane X nuclear receptor (PXR), thus inhibiting the activity of the deactivating enzyme, 24-hyroxylase (CYP24A1), which would normally degrade it to a mono-hydroxy vitamin D (e.g., 25[OH]D) (111,114). The end effect of this VDR dysfunction is lowered 25(OH)D, implying low vitamin D status as usually measured, and elevated 1,25(OH)2D (though this is seldom measured). In other words, the low level of 25(OH)D observed in autoimmune disease is the result of the autoimmune disease process rather than its cause (113). In support of this explanation, there is evidence that some autoimmune diseases can be reversed by gradually restoring VDR function with the administration of a VDR agonist, olmesartan, in conjunction with bacteriostatic antibiotics (111). Eight of nine genetic studies have implicated SNPs of both the VDR or 1-alpha-hydroxylase genes in HT risk, though results are not wholly consistent (partly reviewed in Djurovic et al. (146) and see Supplementary Table S1; Supplementary Data are available online at
Is vitamin D status adequate?
A review of six different geographical regions has demonstrated that vitamin D deficiency is widespread across the world (147). Although both the Institute of Medicine (148) and the European Food Safety Authority (EFSA) (149) consider that a dietary intake that achieves a serum 25(OH)D concentration of 50 nmol/L is sufficient, a number of organizations prefer to define sufficiency as the higher value of 75 nmol/L, values between 75 and 50 nmol/L as insufficient, and those <50 nmol/L as deficient (141). In most of the studies that have investigated the relationship between vitamin D and autoimmune thyroid disease, vitamin D deficiency has been defined as a serum 25(OH)D concentration <50 nmol/L (20 ng/mL). There are no available data on the optimal vitamin D concentration that can support diverse tissue responses, though it appears likely that local tissue levels need to be higher than typical serum levels (139). Locally synthesized 1α,25(OH)2D3 has been shown to be degraded immediately after its autocrine action without entering the circulation. Hence, even measurements of serum 1α,25(OH)2D3 may not be meaningful (139).
Recommendations for vitamin D
It appears highly questionable that the low serum/plasma 25(OH)D concentration in HT patients is a true reflection of a deficient vitamin D status, let alone that vitamin D deficiency is a cause of HT. Studies need to measure serum/plasma concentrations not only of 25(OH)D but of 1,25(OH)2D and indeed of 24,25(OH)2D to get a clearer picture. Even then, the concentrations in thyroid and immune cells will not necessarily be revealed. Trials are needed to elucidate the association between vitamin D and HT so that clear evidence-based suggestions can be made. In the meantime, however, it would be wise to ensure that patients avoid overt vitamin D deficiency.
However, it has been suggested that at high levels (e.g., achieved from supplementation), 1,25(OH)2D may have the potential to displace the natural ligands from nuclear receptors such as the α- and β-thyroid hormone receptors (111,113). If T3 is indeed displaced from the thyroid hormone receptors, there may be adverse effects on the endocrine system. New data also show that the greater the increase in 25(OH)D on supplementation, the greater the conversion to the inactive 24,25(OH)2D, resulting in a null effect (150).
Conclusions
HT affects more people than any other autoimmune condition. Hence, awareness of the nutritional factors discussed above that can interact to alter the risk, progression, or development of HT or associated conditions can provide an additional strategy in the hands of concerned clinicians to the benefit of a large number of patients.
Footnotes
Acknowledgments
This project was partly supported by a grant from the Office for the Education of Talented Students of Xi'an Jiaotong University.
Author Disclosure Statement
No competing financial interests exist.