
Abstract
The global incidence of thyroid cancer is increasing, and metastatic spread to the lymph nodes is common in papillary thyroid carcinoma. The metastatic course of thyroid carcinoma is an intricate process involving invasion, angiogenesis, cell trafficking, extravasation, organ specific homing, and growth. A key aspect in this process involves a multitude of interactions between chemokines and their receptors. Chemokines are a group of small proteins, which act to elicit normal physiologic and immune responses principally through recruitment of specific cell populations to the site of infection or malignancy. Thyroid cancer cells, like other tumors, possess the ability to corrupt the chemokine system to their advantage by altering cell movement into the tumor microenvironment and affecting all aspects of thyroid cancer progression.
Introduction
T
Chemokines (chemo attractant cytokines) are a small conglomerate of specialized proteins, which act together through their cell surface receptors to coordinate a series of actions. The secretion of chemokines by particular cell types promotes homeostasis, organ development, and immune responses by directing cells to specific locations throughout the body (2). Due to the varied and dynamic expression of multiple receptors on the surface of leukocytes and endothelial and tumor cells, chemokines recruit specific cell populations as required.
There are four major groups of chemokines: CXC, CC, CXC3, and C (Fig. 1). Their nomenclature varies depending on the position of the conserved cysteine residues with the corresponding receptors binding to the ligand (2). Chemokines and their receptors are generally known by their systematic names consisting of the family of chemokine followed by the letter “L” for “ligand” or “R” for “receptor,” with a number indicating their order of discovery. For example, the chemokine originally named Stromal derived factor 1 is now termed CXC chemokine ligand 12 (i.e., CXCL12), and its associated receptor is CXCR4 (3). To date, there have been 50 chemokines identified, with 18 chemokine receptors described (4) (Table 1). There are more chemokines than receptors, meaning that many cell surface receptors bind to multiple chemokines and vice versa. This type of system is common within biology and is said to possess redundancy, which means that key pathways are not solely dependent on single factors (5,6).
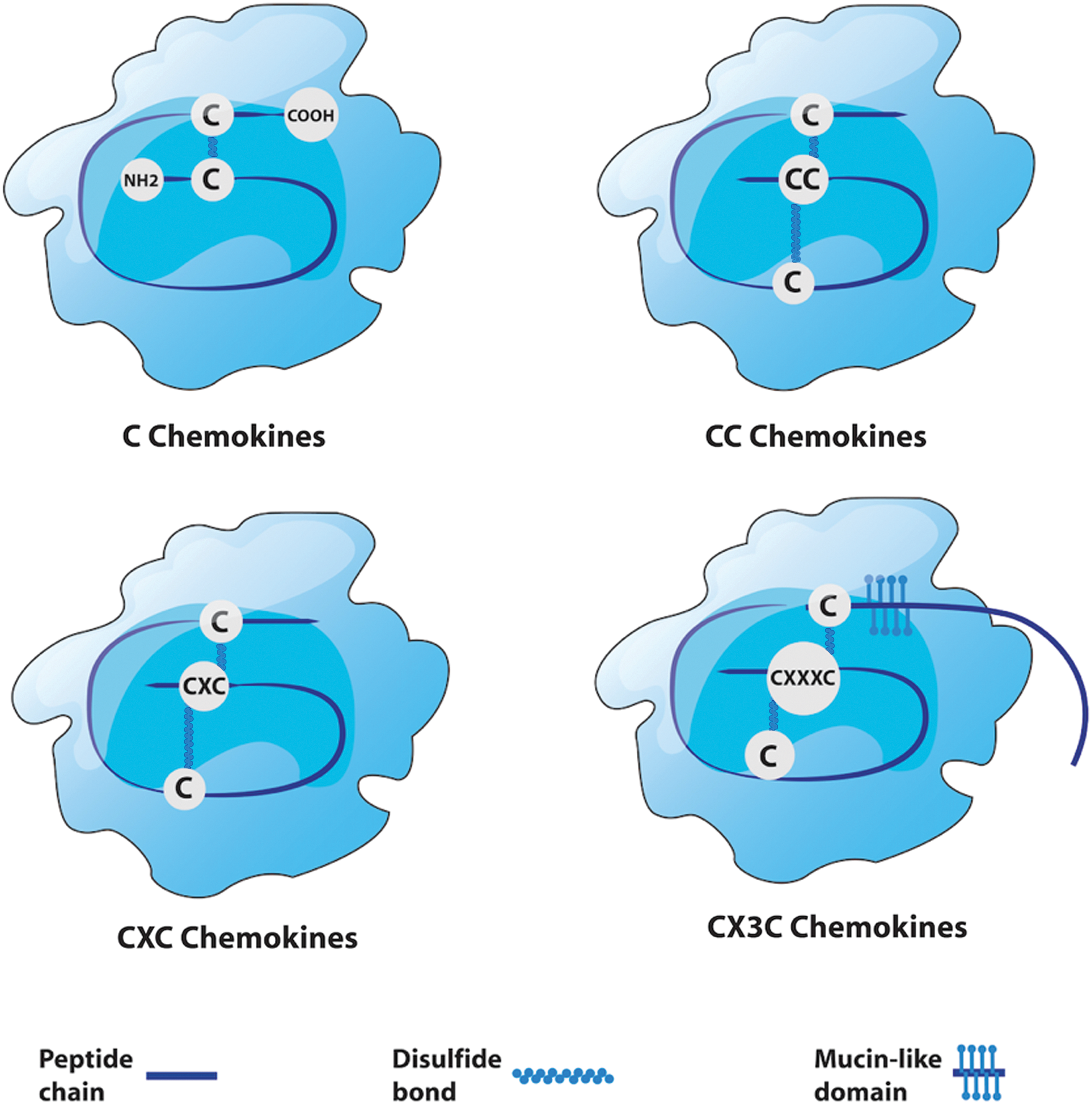
Structure of chemokine subtypes. Chemokines are divided into four subtypes: C, CC, CXC, and CX3C. The CC chemokines are the most abundant group, having two adjacent cysteines near their amino terminus. They principally induce the migration of monocytes, natural killer (NK) cells, and dendritic cells. CXC chemokines have N-terminal cysteines that are separated by one amino acid, represented in their name with an “X.” CXC chemokines are the next most abundant group, and are subdivided into two categories: ELR positive (ELR+) and ELR negative (ELR–). ELR+ CXC chemokines specifically induce the migration of neutrophils and interact with chemokine receptors CXCR1 and CXCR2. C chemokines only have two cysteines, and there are only two chemokines in this subgroup—XCL1 and XCL2—which activate and attract T cells, respectively. CX3C chemokines have three amino acids between the two cysteines. The only one discovered to date is CX3CL1, which is secreted and attached to the surface of the cell that expresses it, serving as a chemoattractant and adhesion molecule. Color images are available online at
Ba, basophil; Eo, eosinophil; iDC, immature dendritic cell; mDC, mature dendritic cell; Mo, monocyte; Mφ, macrophage; NK, natural killer cell; Th1, type I helper cell; Th2, type II helper cell; Treg, regulatory T cell; PTC, papillary thyroid carcinoma; ATC, anaplastic thyroid carcinoma; FTC, follicular thyroid carcinoma; MTC, medullary thyroid carcinoma; LNM, lymph node metastasis.
Chemokine receptors are all structurally related and are members of the seven transmembrane G protein-coupled cell surface receptor family (Fig. 2). Chemokines are highly soluble secretory proteins that either remain free in solution or become immobilized on cell surfaces or more generally in the extracellular matrix (ECM), establishing concentration gradients (7), which facilitates direct movement, or chemotaxis, of cells expressing appropriate receptors to the specific target area (8). The chemokine secretion profile is highly dynamic, and as a tumor develops, the amount and nature of the profile will change. Chemokines are subdivided not only in terms of the receptors they interact with but also their principal roles (e.g., inflammatory) (2,9 –11) (Fig. 3). To date, it is the inflammatory, homeostatic, and atypical chemokine receptors that have been reported to be important with respect to the tumor microenvironment (TME).
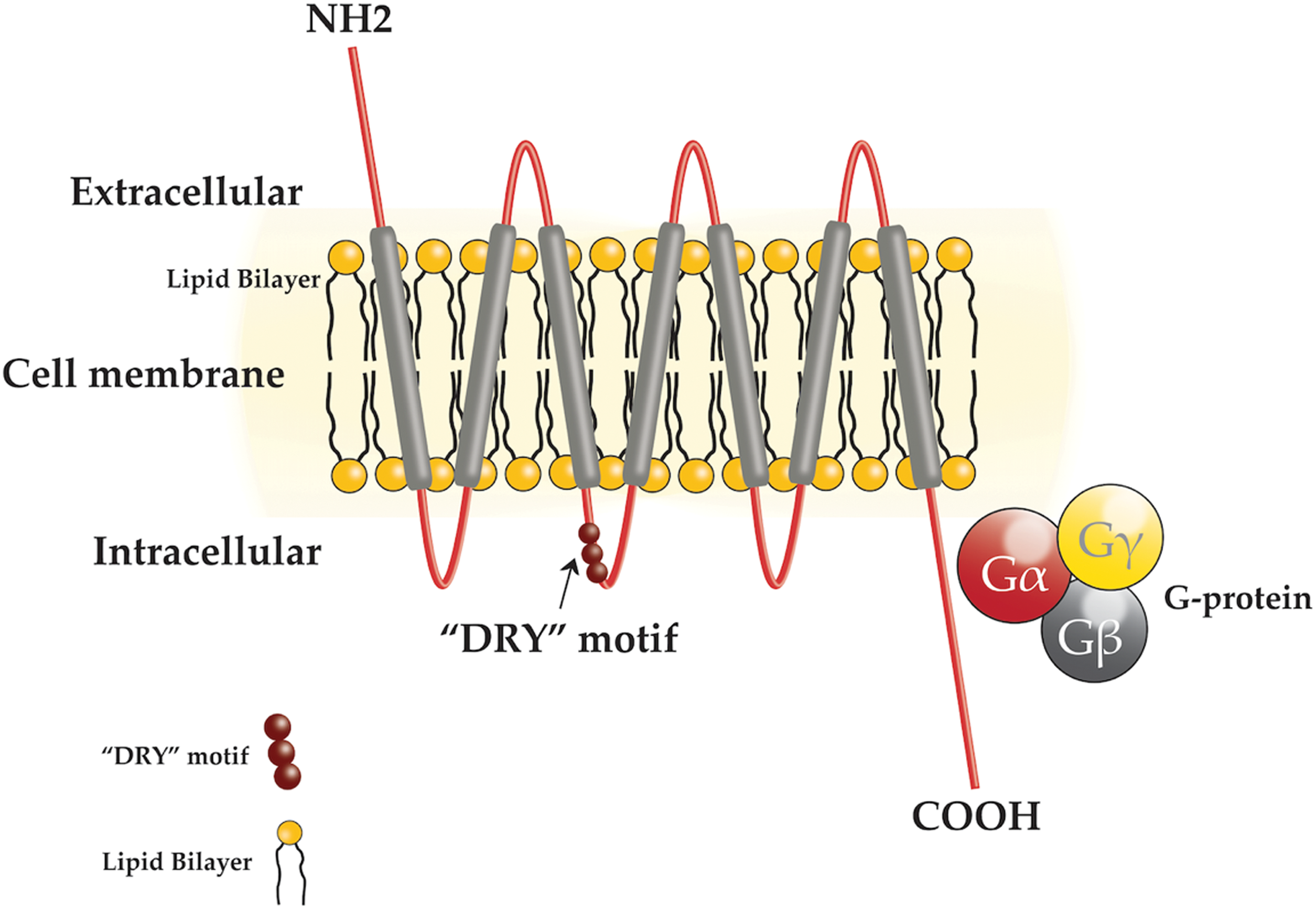
Diagram of a chemokine receptor. Each chemokine receptor comprises an extracellular N terminal, seven transmembrane proteins, three extracellular loops, three intracellular loops, a characteristic “DRY” motif in the second intracellular domain, and a C-terminal segment. These seven transmembrane proteins couple with a “G protein,” making them members of a large protein family known as the G protein (guanine nucleotide binding proteins) coupled receptors. Transmembrane proteins span the entire length of the biological membrane to which they are permanently attached and function as a transporter of various substances across the membrane. G proteins function as molecular switches, which transmit signals from the extracellular environment to the intracellular environment. These proteins are activated by the highly conserved “DRY” motif following binding with the corresponding chemokine ligands, which triggers an influx of intracellular calcium. This induces chemotaxis that acts to drive cells to a desired location within the organism. Color images are available online at
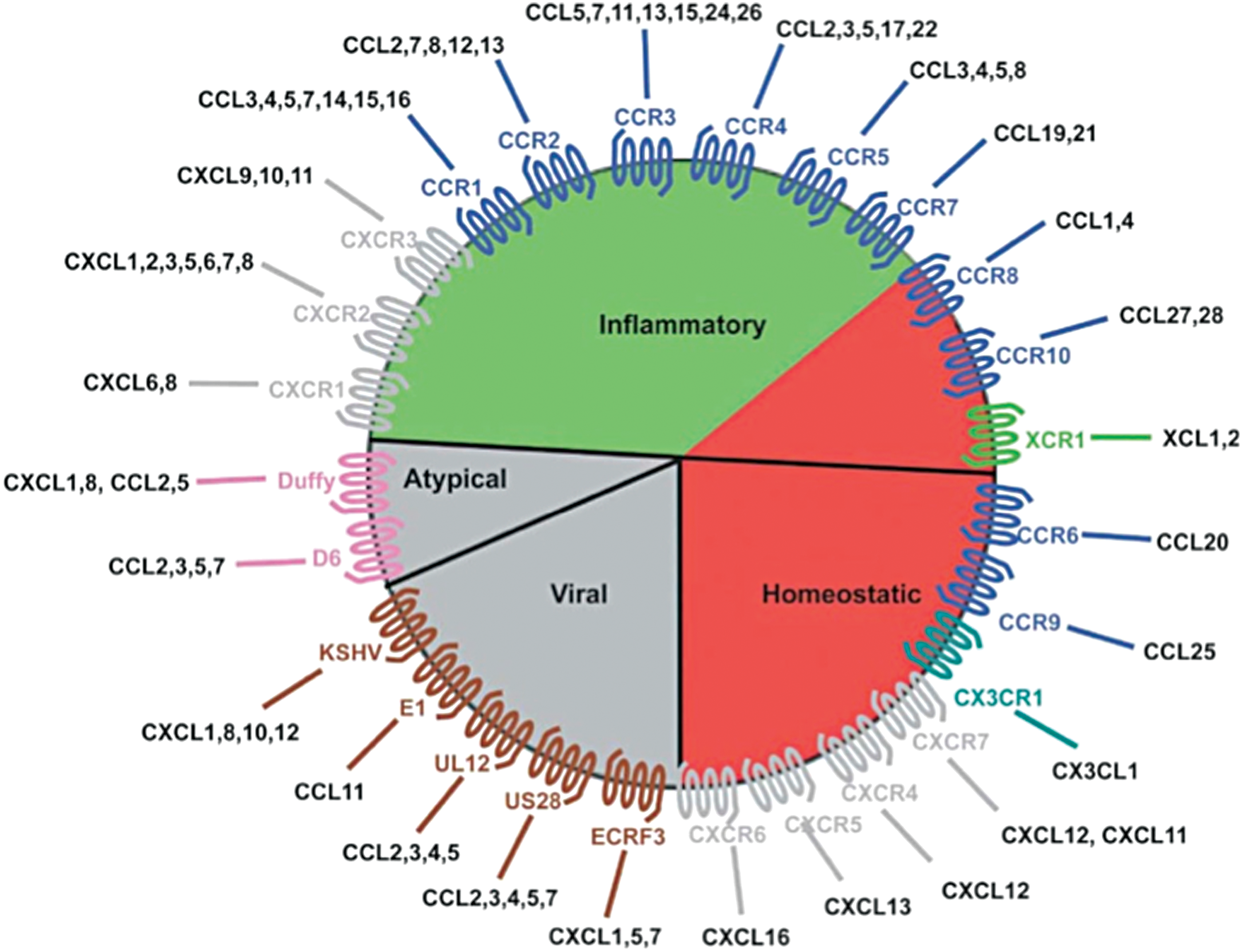
The chemokine wheel. This diagram shows the major features of the chemokine system, chemokine receptors, and their associated chemokine ligands. The inflammatory chemokines are frequently involved with routine immune responses, whereas the homeostatic group contributes to normal development and physiology. The atypical or “silent” receptors act as negative regulators of the system, which decrease the levels of chemokines. Viral chemokines and receptors allow pathogens to change immune responses to infection. Only the inflammatory, homeostatic, and atypical chemokine receptors are found within the thyroid microenvironment. This diagram is reproduced with permission from John Wiley and Sons (2016). Color images are available online at
Malignant cells have found a way to subvert the chemokine system to their advantage via chemotaxis, thereby influencing cell trafficking within the TME (5,12). These “corrupted” chemokines are secreted from both primary tumors and metastatic sites (6). They behave like growth factors (13), contributing to angiogenesis (14), immunosuppression, and proliferation of fibroblast stroma, which in turn increases the survival of tumor cells, enhancing tumor growth and metastatic potential (5,6). This is because malignant cells gain functional chemokine receptors that are often not found on their normal counterparts, and like leukocytes, they can respond to chemokine gradients at sites of metastasis (15,16).
The malignant thyroid microenvironment
It is widely acknowledged that there is a strong relationship between cancer and inflammation. This has been observed since 1863 when Virchow observed leucocytes in neoplastic tissue and hypothesized that chronic inflammation could contribute to the tumorigenic process (17). It is estimated that about 20% of all tumors arise from conditions of chronic inflammation (17), including thyroid carcinoma from chronic thyroiditis (18). The inflammatory microenvironment of thyroid carcinoma is complex, consisting mainly of leucocytes such as macrophages, T lymphocytes, dendritic, mast, and natural killer (NK) cells that circulate among the tumor cells and supporting stroma (19). These leucocytes can account for up to 50% of the tumor mass, with the predominant subset being macrophages (20). Chemokines shown to exist at relatively high levels within the microenvironment are CXCL8 (8,21 –24) and CXCL20 (25 –27); these are produced by both tumor cells and immune cells (Table 1). Production of inflammatory mediators within the thyroid tumor mass also results in de novo chemokine receptor expression on the malignant cells. As expected, a number of receptors have been shown to be upregulated (Table 1). The two most studied are CXCR4 (28 –32) and CCR7 (28,30,32,33).
The majority of thyroid tumor literature has focused on detecting chemokines in the tumor mass and highlighting how these may work through the tumor-associated macrophages, with a relatively small number of studies detailing the role of chemokines in the attraction of other immune cells, most probably because the latter studies are more complicated and expensive. The aim of this review is to comprehensively review the current knowledge of chemokines and their receptors in thyroid carcinoma.
Method
A comprehensive literature search was performed using Pubmed, Google Scholar, and MEDLINE databases between 1996 and 2016 with the following key words: (i) chemokine and thyroid carcinoma, (ii) leukocyte and thyroid carcinoma, (iii) chemokine and metastasis in thyroid carcinoma, and (iv) inflammation and thyroid carcinoma. Four separate searches were performed in each database. A total of 26,200 studies were found, with 140 being relevant to thyroid carcinoma. No restrictions on language were used. Unpublished reports, including dissertations and conference abstracts were not considered.
Inclusion and exclusion criteria
All titles and abstracts of the retrieved articles were reviewed. Studies were included if they met the following criteria: (i) studies investigating chemokines and oncogenic changes in thyroid carcinoma; (ii) studies looking at chemokines, inflammation, and tumor-associated leukocytes; (iii) CK-induced metastatic potential of thyroid carcinoma; and (iv) the therapeutic potential of chemokines in thyroid carcinoma. The major reason for exclusion was studies not being specific to chemokines and thyroid carcinoma.
Protumorigenic Chemokines and Associated Cells in Thyroid Carcinoma
Tumor progression and migration: CXCL8 and CCL20
One of the more abundantly expressed chemokines by thyroid carcinoma cells is CXCL8. This was the first chemokine shown to be secreted by thyrocytes (24), and there is evidence to support a role for CXCL8 and its interaction with receptors CXCR1 and CXCR2 in regulating cell migration, tumor growth, progression, metastatic spread, and tumor-associated angiogenesis in various cancers (8,21,22). Both Yoshida et al. (21) and Weetman et al. (22) observed production of CXCL8 as a chemotactic factor for neutrophils in the supernatants of primary cultures of normal thyroid cells. Since then, Basolo et al. (23) demonstrated that papillary and anaplastic cell lines release large amounts of CXCL8. Kobawala et al. (24) looked at serum levels of CXCL8 in 88 patients with thyroid diseases: 21 with goiter, 16 with autoimmune thyroid disease, and 32 with differentiated thyroid carcinoma (DTC). Serum CXCL8 levels were significantly higher in DTC patients of various histological subtypes compared to autoimmune thyroid disease patients. CXCL8 levels were also similar in healthy controls and early-stage thyroid carcinoma patients, but were significantly increased in patients with advanced thyroid carcinoma.
CCL20 is another prominent chemokine involved in thyroid carcinoma and is the only ligand that activates receptor CCR6 (25). Increased levels of CCL20 are seen in many forms of cancer such as colorectal (34), pancreatic (35), lung (36), and hepatocellular carcinoma (37), and closely correlate to tumor growth (38), metastasis (37), and overall poor prognosis (34 –37). Zeng et al. (26) demonstrated that CCR6 was overexpressed by thyroid cancer cell lines and also that activation of CCR6 by CCL20 promoted invasion and migration while blocking of the receptor reversed the effect. They also observed that CCL20/CCR6 interaction in thyroid carcinoma enhanced tumor growth and invasive capabilities.
Tumor neovascularization and growth: endothelial cells and CXC chemokines
It is well known that neovascularization is crucial for tumor growth, progression, and metastasis, and some chemokines are known to be involved in this process (39). The CXC chemokines can be divided up into two groups: those that have the ELR positive (ELR+) motif, and those that are ELR negative (ELR–). The ELR+ is an important structural domain for angiogenic activity because it determines the specificity of the CK binding to its cognate receptor (39). The ELR+ group includes chemokines CXCL1, CXCL2, CXCL3, CXCL5, CXCL6, CXCL7, and CXCL8 (39) that directly promote migration and proliferation of endothelial cells and eventually neovascularization, mainly by interacting with receptor CXCR2 and not CXCR1 (40). The ELR– groups have antiangiogenic properties and consist of CXCL4, CXCL9, CXCL10, and CXCL11 (40). Furthermore, the Duffy antigen has been shown to suppress the angiogenic effects of the ELR+ CXC chemokines in thyroid cancer, as it can sequester these without eliciting any intracellular signals (27). Thus, the balance between pro- and antiangiogenic CK determines the degree of tumor neovascularization.
Tumor growth, invasion, and survival
Tumor-associated macrophages
The review of the literature highlighted numerous studies looking at the relationship between thyroid carcinoma and tumor-associated macrophages (TAM). TAM are derived from circulating monocytes and are attracted to the TME by a number of CC and CXC chemokines such as CCL2, CCL5, CCL7, CCL8, CCL12 (20), CXCL8 (41), and CXCL16 (42) via their surface expression of receptors CXCR1, CXCR2 (41), and CXCR6 (42).
TAM can differentiate into either M1 type (phagocytic, antitumorigenic) or M2 type (proliferative, protumorigenic) cells. However, within the thyroid TME, TAM generally express M2 polarization (2) and have well-documented tumor-promoting activities (i.e., cancer initiation and progression), and in general, a high TAM count is indicative of poor prognosis (43 –46). This is largely due to TAM promoting tumor progression by producing factors such as VEGF and fibroblast growth factor and suppressing antitumor immunity (47) (Fig. 4).
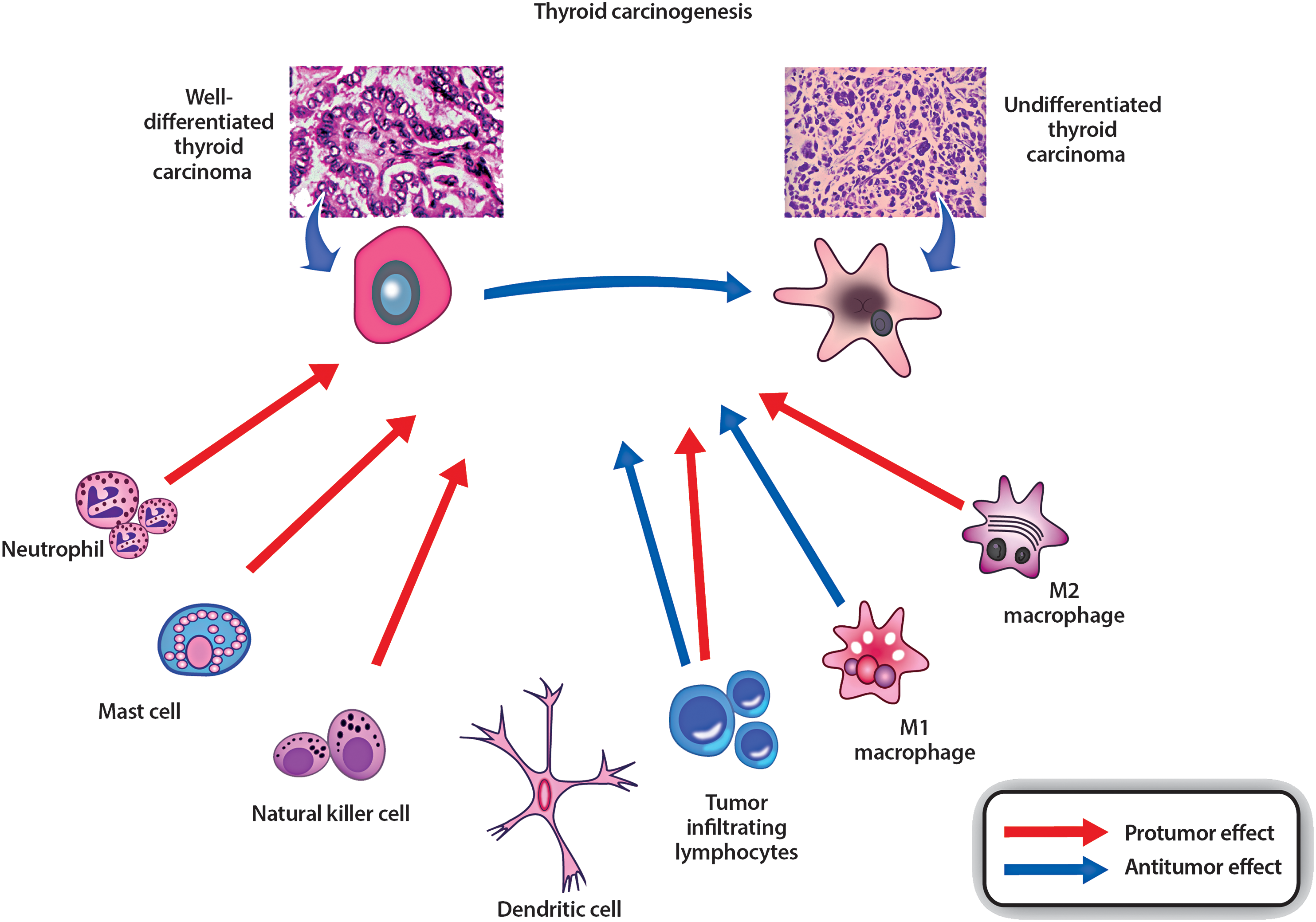
Immune cells involved in anti- or protumor responses. Protumorigenic cells include M2 macrophages, certain lymphocytes, NK cells, mast cells, and neutrophils. Antitumorigenic cells include M1 macrophages, certain lymphocytes, and dendritic cells. The balance of anti/protumorigenic responses may be important for thyroid cancer progression or elimination. Diagram adapted from and reproduced with licensing permission from Bioscientifica Ltd. (2016). Color images are available online at
In 2013 Weiyuan et al. (41) conducted both in vitro and in vivo experiments to evaluate whether the secretion of CXCL8 by TAM promoted the metastatic potential of PTC. In vitro, they analyzed TAM-conditioned medium and its propensity to promote PTC cell invasion. In vivo, they injected mice with both PTC cells and CXCL8, and after 40 days, the metastatic lesions were counted and measured. The results indicated that TAM could initiate and promote PTC by releasing CXCL8, and an increased density of TAM correlated with increased lymph node metastases. TAM density in the tumor core was associated with progressive PTC features, and a medium enriched with TAM enhanced the invasion of PTC. CXCL8 was identified as the most abundant chemokine in PTC-derived TAM. Blocking it with anti-CXCL8 neutralizing antibody significantly decreased PTC metastases, and exogenous CXCL8 enhanced PTC metastases in vivo (41). It was also noted that CXCR1/2 receptors (which CXCL8 binds to) were only present in tumor tissue and were not found in normal healthy thyroid tissue. Therefore, TAM may facilitate PTC cell metastases through CXCL8 and its paracrine interaction with CXCR1/2 (41).
Furthermore, a recent study by Cho et al. in 2016 (42) found that CXCR6 expression positively correlated with lymph node metastasis, and CXCL16 signaling mediated macrophage-enhanced PTC tumor cell invasion and also changed the macrophage phenotypes into M2 macrophages in the PTC environment.
A study of 398 patients with DTC and 132 patients with benign thyroid tissue indicated that TAM were found more frequently in aggressive cases with metastasis at diagnosis, but conversely, macrophage infiltration was also associated with improved disease-free survival (48). Three reasons were identified for this paradoxical result: (i) varying microenvironments in different histological subsets, (ii) TAM modulation of the TME by secretion of a rich repertoire of chemokines and growth factors that exert paracrine effects on tumor cells to facilitate progression, and (iii) TAM phagocytic activity, which could also limit tumor progression (49). This was further confirmed by Fiumara et al. (50) in a study of 121 well-differentiated thyroid carcinoma specimens demonstrating that tumors with TAM in situ showed evidence of active neoplastic cell phagocytosis, which was in turn positively correlated with infiltration of lymphocytes and dendritic cells (DCs) and negatively correlated with vascular invasion. It is possible that these different results are also related to different populations (50).
Mast cells
Mast cells are an important constituent of the thyroid carcinoma microenvironment. Recent studies indicate that PTC induces mast-cell activation and chemoattraction via secretion of CXCL1, CXCL10, and CXCL8 (51). Mast cell numbers are elevated in PTC compared to normal thyroid tissue, and their increased density correlates with a poor prognosis (51). Further experiments using mast cell lines showed that PTC culture medium induced mast-cell activation and chemoattraction, thereby increasing vascularity, proliferation, survival, invasiveness, and DNA synthesis of cancer cells and inhibiting apoptosis. These actions were mediated by histamine, CXCL1, and CXCL10. Therefore, targeting these pathways may be of therapeutic benefit in the future.
Tumor invasion, metastasis, and aggression
CCR7
CCR7 is present in benign thyroid tissue and in PTC and medullary thyroid carcinoma (MTC) (30). It binds to CCL21, which causes proliferation and migration of tumor cells (30,33) and also plays a critical role in lymphocyte and dendritic cell trafficking into lymph nodes, which are the preferential sites of metastasis of PTC and MTC (33).
It is well established that poorly differentiated thyroid carcinoma preferentially metastasize to the bone, lung, liver, and brain, whereas DTC metastasize to lymph nodes. PTC has two variants: follicular variant of PTC, and classic PTC. The latter has a higher propensity for lymph node metastasis. Sancho et al. (33) analyzed CCR7 expression in 46 normal thyroid tissue and thyroid carcinoma specimens. The expression of CCR7 was higher in classic PTC and MTC, which are both prone to lymph node metastasis compared to follicular variants, and was absent in healthy thyroid tissue. As only a subset of PTC develop lymph node metastasis, this would support the concept that CCR7 signaling is not only involved in proliferation and migration but may also play a key role during the tumor initiation phase. CCR7 expression is regulated in vitro by extracellular conditions, namely nutrient deficiency, which may mimic areas inside tumors with high proliferation rates (33), and its activation is significantly increased after 48 and 72 hours of stimulation with CCL21, suggesting that the lymph nodes, where CCL21 is abundant, provide favorable conditions for the proliferation of metastatic thyroid cells once they migrate and arrest at these secondary sites.
CXCR4
CXCR4 is the most widely expressed chemokine receptor in malignant cells (12,19,52) and plays a crucial role in migration, growth, and metastasis (30,53,54) upon interaction with the ligand CXCL12. The binding of CXCL12 to CXCR4 has been shown to promote metastasis and survival in numerous human tumors, including the lung (55,56), breast (57), prostate (58), and colon (59). CXCR4 is not expressed at appreciative levels by normal thyrocytes. However, it is upregulated in normal thyrocytes transformed by the RET oncogene, which subsequently increases the proliferation, survival, and migration of these cells in vitro (28,31). Wagner et al. (30) conducted in vitro experiments looking at the expression of CXCR4 and CXCL12 on 88 samples of PTC using immunohistochemistry. Staining was matched with clinicopathologic features, and revealed that high-intensity staining for CXCR4 expression by the malignant cells was associated with larger tumor size and tumor aggression. These findings suggest that tumor cells may stimulate their own growth and invasion via an autocrine signaling mechanism (30).
Gonzalez et al. (32) found increased expression of CXCR4 and CCR3 in PTC compared to the control thyroid tissue, indicating that these two receptors could be potential markers for PTC (32). In addition, increased CXCR4 correlated with lymph node metastases and extranodal extension in the classical variant of PTC, suggesting that great expression of CXCR4 could be a factor contributing to aggressive tumors and negative prognosis (60,61).
Hwang et al. (29) demonstrated increased expression of CXCR4 on anaplastic thyroid carcinoma (ATC) cell lines compared to that of normal thyroid cells, and CXCL12 increased migration but not proliferation of ATC cells. The CXCL12/CXCR4 activation caused stimulation of ERK1/ERK2 (intracellular pathways designated to influence cell proliferation), and inhibiting these pathways by neutralizing them with anti-CXCR4 significantly decreased ATC cell migration.
CXCR7/CXCL12
CXCR7 is a receptor for CXCL12, which is expressed in many cancer cell lines. It is produced by human thyrocytes (29) and is involved in cell migration and metastasis (62). Using immunohistochemistry, Zhen et al. (62) showed higher expressions of both CXCL12 and CXCR7 in PTC compared to benign tissue (nonmalignant tissue 2 cm from the edge of the cancer specimen; n = 79), and these expression patterns correlated significantly with lymph node metastasis. Additional work by others showed that downregulation of CXCR7 in PTC suppressed cell growth and invasion and induced S-phase arrest, as well as promoting apoptosis, indicating that it could be a promising target for therapeutic intervention (2).
CXCR1
CXCR1 is one of two high-affinity CXC chemokine receptors for CXCL8, and it is a major mediator of immune and inflammatory responses in tumor growth. It routinely induces chemotaxis of leukocytes and tumor cells. Tang et al. (63) looked at the expression of CXCR1 using immunohistochemistry in 129 PTC, 61 nodular hyperplasia, and 118 normal thyroid specimens. They found significantly higher expression in PTC specimens. Furthermore, CXCR1 expression was correlated with increased incidence of lymph node spread and has been suggested as a potential tool for predicting this risk in the future, but additional confirmatory studies are required.
A schematic diagram showing the protumorigenic chemokines and their receptors in the thyroid TME is shown in Figure 5.
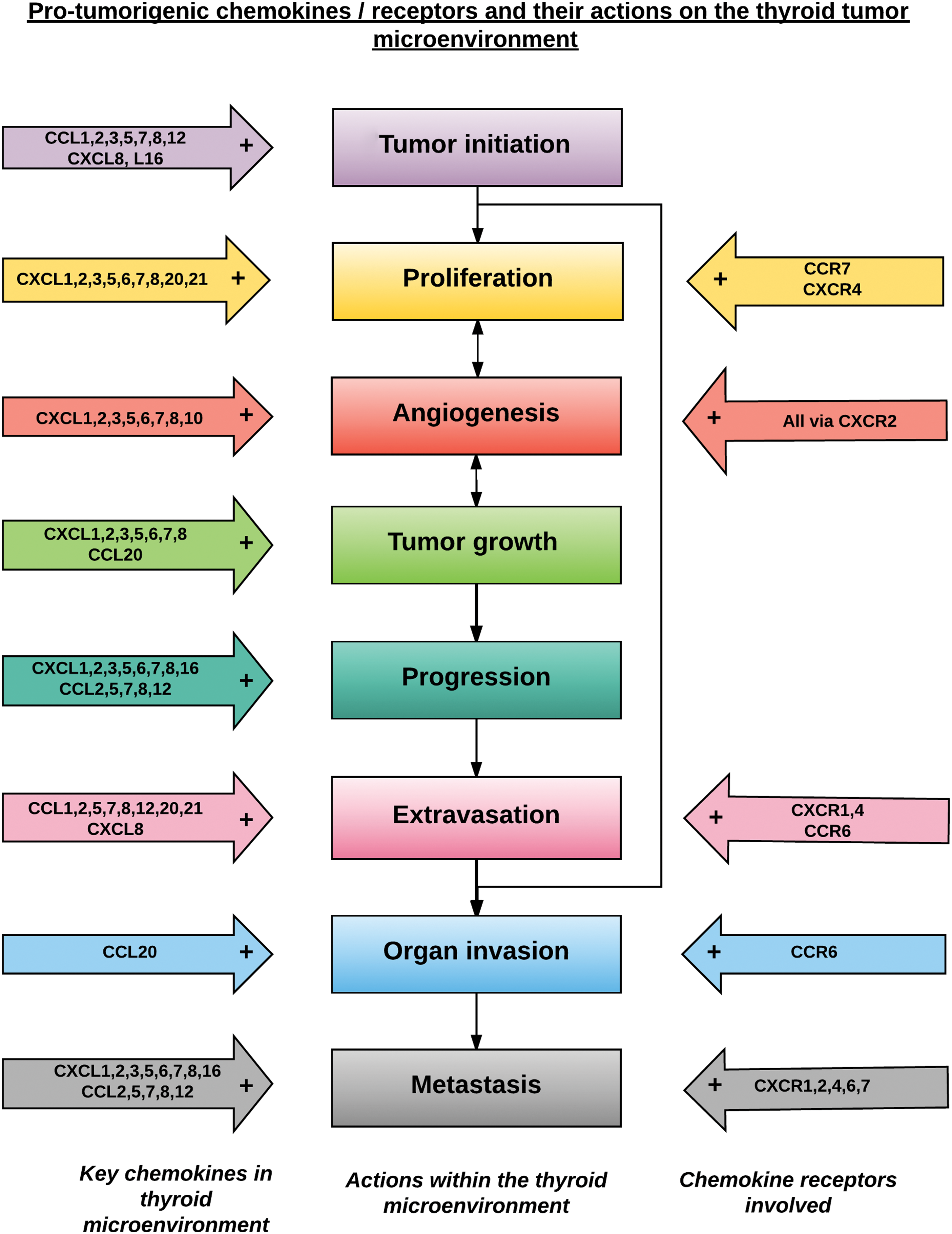
Schematic diagram showing a possible order of sequential stages in tumor development, with the associated protumorigenic chemokines and receptors involved at each step within the tumor microenvironment identified. Color images available online at
Antitumorigenic Chemokines and Associated Cells in Thyroid Carcinoma
Decreased cell growth and increased apoptosis: CXCL10 and NK cells
NK cells are lymphocytes that can kill a broad range of tumor cells and are also found in the tumor mass (64). NK cells migrate to inflamed tissue, including tumor sites and lymph nodes, in response to signaling through chemokine receptors CCR1, CCR2, CXCR3, and CX3R1 (65). Gogaldi et al. (66) investigated NK infiltration in 65 patients with PTC and 25 patients with thyroid nodular goiter. They found a significantly increased number of NK cells in the PTC samples, and demonstrated an inverse correlation between NK infiltration in advanced stages of the disease, suggesting that the changes in the malignant TME could render NK cells redundant (66). Furthermore, a recent study in 2014 by Wenneberg et al. (67) demonstrated that ATC cell lines are sensitive to lysis by CXCR3 positive NK cells.
CXCL10, which binds to CXCR3, is upregulated in PTC (68 –70), and its secretion is increased tenfold by induction with interferon gamma (IFN-γ) and tumor necrosis factor alpha (TNF-α) in PTC compared to normal thyroid follicular cells (TFC) (71). CXCL10 is involved in chemotaxis, induction of apoptosis, regulation of cell growth, and mediation of angiostatic effects (72), mediating its effects in T cells, macrophages, or NK cells (73,74). In 2012, Antonelli et al. (75) compared the expression of CXCL10 in ATC and normal TFC. They demonstrated that only ATC produced CXCL10 under basal conditions. However, IFN-γ stimulated secretion of CXCL10 in both cell lines in a dose-dependent fashion. IFN-γ and TNF-α induced a synergistic but variable release of CXCL10 in different ATC samples. However, the effect was more reproducible in the TFC biopsies.
Angiostasis, decreased metastasis, and recurrence, improved prognosis: DARC/Duffy antigen and lymphocytes
Duffy or Duffy antigen/receptor for chemokines (DARC) is a glycosylated membrane protein that acts as a decoy by scavenging and binding to angiogenic chemokines such as CCL2 and CXCL8 (CK bearing the ELR+ motif) (76,77). Previous studies of DARC function in the breast (78,79), lung (80), and laryngeal cancer (81) have associated DARC overexpression with better prognosis, decreased metastatic potential, and inhibition of tumor-associated neovascularization.
Using immunohistochemistry, Latini et al. (76) demonstrated decreased DARC expression in malignant thyroid tissue compared to that of benign origin. Diminished DARC expression in malignant samples also correlated with increased tumor size and lymphocyte infiltrate. There was no significant link between DARC and metastasis probably due to the propensity of PTC to metastasize via lymphatics rather than blood. It was concluded that larger tumors with decreased DARC expression may present more chemokines to the TME. This in turn could foster increased vascularization and tumor growth. This study clearly indicates that expression of these atypical CK receptors may be a key step in arresting tumor development. However, further research needs to be done to confirm this.
Lymphocytes are frequently found within and surrounding thyroid tumors (82,83). Their recruitment and trafficking is controlled by CC and CXC chemokines, particularly CXCR3 and associated ligands CXCL9 and CXCL10 (84). As cytotoxic T lymphocytes (CTL) can recognize tumor-associated antigens, they may confer some protection against the malignant process by attacking tumor cells (85). However, as mentioned previously, it is the balance between the type of cells present that will ultimately determine the nature of the TME. In 2011, Villagein et al. (86) investigated 157 patients with thyroid carcinomas and classified them by the degree of lymphocytic infiltration (i.e., diffuse, peri-tumoral, and absent). After a mean follow-up period of eight years, significantly higher recurrence occurred in the group with absent lymphocytic infiltration, suggesting that the presence of lymphocytes in the TME may be associated with a more favorable prognosis in patients with DTC, thus confirming previous similar studies done by Matsubayashi et al. (87), Gupta et al. (88), and Modi et al. (82).
Oncogenic changes inducing a pro-inflammatory response in thyroid carcinoma
In PTC, genomic rearrangement and activating point mutations create dominantly transforming oncogenes, the most common and best studied being RET/PTC1 (89,90) and V600E in the BRAF gene (91), which both contribute to the induction of a protumorigenic inflammatory microenvironment. RET is a proto-oncogene encoding for a tyrosine kinase receptor.
The RET/PTC1 oncogene has been shown to induce the expression of a pro-inflammatory program, which includes the upregulation of various chemokines, including CCL2, CCL20, CXCL8, CXCL1, CXCL10, and CXCL12, (28,68,92) and the chemokine receptor CXCR4 (28,68), along with other proinflammatory cytokines and enzymes. Mantonavi et al. (6,19) postulated that rearrangement of the RET tyrosine kinase (RET/PTC) in thyrocytes represents a frequent, early, causative, and sufficient genetic event in the pathogenesis of PTC (6). CXCL8 was also shown to be associated with angiogenesis and induction of the chemokine receptor CXCR4 on transformed thyrocytes. Furthermore, there was an abundance of key elements of the RET/PTC activated program in biopsy specimens, and it was noted that patients with lymph node metastasis had higher levels of inflammatory molecules in their primary tumors.
Melillo et al. (68) demonstrated that activation of the RAS and BRAF/ERK pathways were also able to induce upregulation of chemokines that contribute to neoplastic proliferation, survival, and migration. Inflammation-related genes were further investigated by Muzza et al. (93) in a large series of PTC divided into two groups according to the presence or absence of tumor-associated thyroiditis. RET/PTC changes were more abundant in patients with both PTC and thyroiditis, whereas BRAFV600E was significantly more common in patients with PTC without thyroiditis. Fugazzola et al. (17) demonstrated that expression of chemokines CCL20 and CXCL8 was enhanced either in BRAFV600E or RET/PCT tumors compared to controls. Interestingly, thyroiditis specimens displayed the same levels of expression of CCL20 and CXCL8 compared to normal healthy thyroid tissue, suggesting that these chemokines could be associated with tumor-related inflammation rather than a primary autoimmune process. Torregrossa et al. (94) retrospectively evaluated CXCR4 expression in 200 PTC and compared the following clinicopathologic features: type of PTC, degree of infiltration, cancer stage, presence of lymphoid thyroiditis, and mutation status of the BRAF gene. They reported that raised CXCR4 expression was strongly correlated with the BRAFV600E mutation, and in combination, these factors were associated with local aggressiveness of neoplastic cells. This could induce and promote a more aggressive phenotype in PTC, thereby increasing tumor spread outside the thyroid gland.
Chemokines and Their Receptors as Potential Screening Tools and Therapeutic Targets in Thyroid Carcinoma
Enhancing antitumorigenic chemokine pathways and tumor-mediated immunity
As TAM are the most predominant leucocytes in tumors (20) and can promote tumor progression by inducing angiogenesis (43 –46), blockade of common TAM-associated chemokines such as CCL2 and CXCL8 and their associated receptors could be a potential target for therapy. Migration of DCs to lymph nodes is mediated by CCL19 and CCL21 via CCR7 (8). Although there are no studies demonstrating the role of DCs and their associated chemokines in thyroid cancer, injection of CCL21 into mice with bilateral multifocal lung adenocarcinoma showed increased lymphocytes and DCs at the tumor site and reduction in tumor burden, which enhanced survival (95). Therefore, further research into the possibility of therapies based on DCs for thyroid carcinoma may be warranted. Neovascularization is also key for tumor growth, progression, and metastasis (96), and targeting the ELR+ CXC chemokines may arrest tumorigenesis (8). Alternatively, the actions of ELR+ chemokines can be countered by addition of ELR– chemokines, which inhibit angiogenesis (8).
Promoting oncogenic cellular senescence
Oncogenic cellular senescence or oncogene induced senescence (OIS) is defined as a stable and long-term loss of proliferative capacity, despite continued viability and metabolic activity (97). In 2012, Weyemi et al. (98) first described RAS-induced senescence in thyroid cells by transducing human thyrocytes with an expression vector carrying cDNA for H-RasV12. The expression of the mutated RAS induced DNA damage responses by triggering generation of reactive oxygen species, which are critical for promoting OIS. OIS acts as a protective mechanism against oncogenic transformation by suppressing unscheduled proliferation of any neoplastic cells (99).
BRAF mutations, most commonly V600E as mentioned above, are found in about 40% of PTC and play an important role in tumor initiation and progression. BRAF also downregulates the expression of genes involved in thyroid hormone synthesis and promotes dedifferentiation in PTC (100,101). In PTC, the frequency of malignancy increases as levels of thyrotropin (TSH) increase (102,103). In 2016, Zou et al. (104) conducted experiments looking at the effects of BRAFV600E -induced PTC, which were transplanted into mice with both normal TSH levels (nude mice group) and mice with elevated TSH levels (TPO transgenic BRAF mice). The results showed heavy lymphocytic and macrophagic infiltration with regression of transplanted tumors in both groups secondary to OIS. It was noted that the levels of transformation related protein 53 (Trp53), a tumor suppressor gene, increased in tumors undergoing OIS regression, and Trp53 deactivation reversed OIS and enabled tumor growth with the characteristics of ATC (i.e., transformation of PTC to ATC tumor cells). BRAF-induced PTC could be regulated via a p53-dependent mechanism. However, both OIS and TSH seemed to cause major disruption of this mechanism. Zou et al. therefore stressed the importance of controlling serum TSH in PTC patients.
Furthermore, cells undergoing OIS interact mainly with CXCR2 along with CCL20, interleukin (IL)-8 and IL-6, and knockdown of either CXCR2 or IL-8 can lead to a reduction in OIS and re-proliferation of tumor cells (99). Therefore, increasing the CXCR2/IL-8 complex could be used as a potential therapeutic target to promote the arrest of cellular growth and eventually delay the early onset of tumorigenesis (99).
Use as clinical biomarkers
CXCL8 (IL8) and its receptor CXCR2 are increased in MTC, with higher levels in patients with lymph node metastases (105 –107). Sunitinib is a multikinase inhibitor with both antiangiogenic and antitumoral activities (107). Broutin et al. (107) demonstrated that the CXCL8 gene displayed major changes during sunitinib treatment both in vivo and in vitro, indicating that secreted CXCL8 can be used as a biomarker of favorable clinical response to sunitinib. Administration of the drug to tumor-inoculated mice reduced tumor growth and angiogenesis but also significantly decreased the serum concentration of CXCL8. Resistance to the drug was countered by administering CXCL8 neutralizing antibody, which re-sensitized the tumor to it, thus indicating that CXCR8 is a useful positive biomarker and a therapeutic target of sunitinib. As mast cells are increased in PTC and their abundance is positively correlated with tumor growth, survival, and invasiveness, monitoring the density of mast cells could be used as a clinical biomarker of tumor progression, and targeted blocking of chemokine CXCL1 and CXCL10 pathways may be of therapeutic benefit in the future.
Conclusion
It is evident that research during the last three decades has found multiple links between the chemokine system and cancer as our understanding of the complex signaling pathways has developed. Chemokines and their receptors are involved in every stage of thyroid cancer development, from control of the tumor cellular microenvironment to malignant cell survival and metastatic spread. Cancer cells are known to alter the chemokine system that under normal circumstances enable healthy embryonic development, homeostasis, and successful immune responses to a pathogen. With further research, it may be possible to identify common chemokine markers and pathways by blood or tissue testing, which may have the potential to characterize further and treat some forms of thyroid carcinomas and prevent metastatic progression.
Footnotes
Author Disclosure Statement
The authors have nothing to disclose.