
Abstract
Background:
The BRAFV600E mutation is the most frequent genetic abnormality in adult papillary thyroid carcinomas (PTCs). On the other hand, various chromosomal rearrangements are more prevalent in childhood and adolescent PTCs. The aim of the present study was to identify novel rearrangements in PTCs from young patients.
Methods:
Among 63 postoperative specimens of childhood and adolescent PTCs, which had been discovered by the thyroid ultrasound screening program in Fukushima, nine samples without prevalent known oncogenes, BRAFV600E , RAS, RET/PTC1, RET/PTC3, and ETV6/NTRK3, were analyzed in the current study by quantitative real-time reverse transcription polymerase chain reaction to screen for novel fusion genes by comparing transcript expression between extracellular and kinase domains of ALK, NTRK1, NTRK3, and RET.
Results:
Of the above nine samples, five samples were suspected to harbor a fusion, and using subsequent 5′ rapid amplification of cDNA end (RACE), two already reported fusion oncogenes, STRN/ALK and TPR/NTRK1, and three novel fusions, SQSTM1/NTRK3, AFAP1L2/RET, and PPFIBP2/RET, were identified. Functional analyses of these three chimeric genes were performed, and their transforming abilities were confirmed through the activation of mitogen-activated protein kinase (MAPK).
Conclusions:
Three novel fusion oncogenes have been identified in young PTC patients in Fukushima, suggesting that rare fusions may be present among the cases negative for known oncogenes in this age group and that such rearrangements can play a significant role in thyroid carcinogenesis.
Introduction
P
In adult sporadic PTCs, a point mutation in BRAF, especially BRAFV600E , is the most prevalent genetic abnormality (4,6). In children, however, it has been reported that the prevalence of BRAFV600E is much lower (0–37%) (7,8), and various chromosomal rearrangements, including RET/PTC, are more frequent (7,9 –15). The accumulating knowledge of genetic abnormalities in different age groups has increased our understanding of carcinogenic mechanisms of PTCs. However, the reason of different oncogenic profiles between adults and childhood PTCs is still unclear.
After the accident at the Fukushima Daiichi Nuclear Power Plant, the Thyroid Ultrasound Screening Program was started beginning in October 2011 for all children aged 0–18 years old at the time of the accident (16,17). Although many PTCs were found during the first round of screening, those are thought to be spontaneous because (i) the estimated thyroid dose of radiation exposure was quite low, (ii) the latency period was too short, and (iii) most of the patients were teenagers (17). Indeed, the purpose of the first-round screening was to understand background ultrasound findings in thyroids of this age group. Presumably, these PTCs all reflect naturally occurring tumors, including latent cancers in the young population, and many of them could be silent or slowly growing until middle age. However, they were discovered by mass screening using highly sensitive ultrasound instruments. Therefore, it is highly important to perform detailed analysis of these cases because it may help to understand the etiology, mechanisms, and natural course of PTCs in children and other age groups. It has been reported that approximately 85% of the PTCs in the Fukushima area harbor known driver mutations such as the BRAFV600E mutation and the RET/PTC rearrangement (18). However, the oncogenic drivers in the remaining 15% remain to be identified. The present study screened these cases for novel fusion oncogenes to clarify the oncogenic profile of these PTCs further.
Materials and Methods
Samples
In a previous work (18), a total of 63 childhood and adolescent PTC patients operated at Fukushima Medical University Hospital were examined for the presence of the following known genetic abnormalities: BRAFV600E , RAS, RET/PTC1, RET/PTC3, and ETV6/NTRK3. The mean age at the time of surgery was 17.3 ± 2.8 years (range 9–22 years). The sex distribution was 22/63 (34.9%) male and 41/63 (65.1%) female. The size of the tumors varied from 6 to 40.5 mm (M = 14.1 ± 8.2 mm). After surgery, all tissue samples were reviewed by a thyroid pathologist to confirm diagnosis. The study was approved by the ethics committees of Nagasaki University and Fukushima Medical University. Written informed consent was obtained from each patient.
In the current study, nine samples that did not harbor any of the mutations listed above were included. RNA/cDNA samples were used, which were generated in a previous work (18). These samples were screened for possible rearrangements of ALK, NTRK1, NTRK3, and RET by quantitative real-time reverse transcription polymerase chain reaction (qRT-PCR).
Fusion gene screening by qRT-PCR and 5′-RACE
To detect rearrangements, samples were screened for the imbalance in expression of exons 3–4 relative to exons 27–29 of ALK, exons 1–3 relative to exons 15–16 of NTRK1, exons 8–9 relative to exons 15–16 of NTRK3, and exons 2–3 relative to exons 17–18 of RET. qRT-PCR was performed in a Thermal Cycler Dice real-time system (TaKaRa Bio) using SYBR Premix Ex Taq II (Takara Bio). The following PCR primers were used: for ALK ex3–4, 5′-GGG CAG AGC GTT CTA AGG AGA TG-3′ (forward) and 5′-CGG CCA GTG TGC AGT GCT-3′ (reverse); for ALK ex27–29, 5′-CCC TGG GCC TGT ATA CCG GAT A-3′ (forward) and 5′-TAC ATC CGG GTC CTG GGT GC-3′ (reverse); for NTRK1 ex1–3, 5′-CCC TGG ATA GCC TCC ACC AC-3′ (forward) and 5′-ACG GAG ACC ACT CTT CAC GAT G-3′ (reverse); for NTRK1 ex15–16, 5′-CGA GAG CAT CCT GTA CCG TAA GTT C-3′ (forward) and 5′-CCC TGC GTG ATG CAG TCG-3′ (reverse); for NTRK3 ex8–9, 5′-CAA CCT GAC CGT ACG AGA GGG T-3′ (forward) and 5′-GAT GGC ATG AAC ATT GGT CCA G-3′ (reverse); for NTRK3 ex15–16, 5′-CCG ACC AAG GAC AAG ATG CTT G-3′ (forward) and 5′-GCC GCA CAC TCC ATA GAA CTT GAC-3′ (reverse); for RET ex2–3, 5′-AAG CTC AGT GTC CGC AAC C-3′ (forward) and 5′-ATG TGG GTG ACA GGA AGA CC-3′ (reverse); for RET ex17–18, 5′-TGC AGC GAG GAG ATG TAC C-3′ (forward) and 5′-CCA GGT CTT TGC TGA TGT CC-3′ (reverse).
The 5′ rapid amplification of cDNA end (RACE) was then performed for samples suspected to have fusion genes using a 5′-Full RACE Core Set (Takara Bio) according to the manufacturer's protocol. Briefly, first-strand cDNA was synthesized by reverse transcription using 5′ end-phosporylated RT primers. Sequences of each primer were as follows: for ALK, 5′-CAC CTC CTT CAG G-3′; for NTRK1, 5′-GTA CAG GAT GCT CTC-3′; for NTRK3, 5′-CAC ACT CCA TAG AAC-3′; for RET, 5′-TCT CGC GGA GGA AGC-3′. Next, DNA–RNA hybrid was treated with RNase H to degrade RNA. Then, the single-strand DNA was incubated with T4 RNA Ligase to circularize it or to form concatemers. These are amplified by nested PCR using primers located in the region encoding the kinase domain of each target. First PCR primers were as follows: for ALK, 5′-CTG AGC AAG CTC CGC ACC TCG AC-3′ (forward) and 5′-GCT CTG CAG CTC CAT CTG CAT GG-3′ (reverse); for NTRK1, 5′-AGC AGG GAT ATC TAC AGC ACC GA-3′ (forward) and 5′-CCC ACT AGA CAG TTG CGT GTG G-3′ (reverse); for NTRK3, 5′-GGC TGT GAA GGC CCT GAA GG-3′ (forward) and 5′-GCA CTC GGC CAG GAA GAC CT-3′ (reverse); for RET, 5′-GTC CTG AAG CAG GTC AAC CAC CCA-3′ (forward) and 5′-GAC AGC AGG TCT CGC AGC TCA CTC-3′ (reverse). Second PCR primers are: for ALK, 5′-GCT GGC AAG ACC TCC TCC ATC AGT G-3′ (forward) and 5′-GCT CCT GGT GCT TCC GGC GGT AC-3′ (reverse); for NTRK1, 5′-CAC CAT GCT GCC CAT TCG CT-3′ (forward) and 5′-CAG CCA GCA GCT TGG CAT CA-3′ (reverse); for NTRK3, 5′-CTG CAG CAT GAG CAC ATT GTC A-3′ (forward) and 5′-CTC ACC CAG TTC TCG CTT CAG C-3′ (reverse); for RET, 5′-CGC TCC TCC TCA TCG TGG AGT ACG-3′ (forward) and 5′-AAG AAC CAA GTT CTT CCG AGG GAA TTC C-3′ (reverse). The PCR amplicons were checked by agarose gel electrophoresis and subjected to Sanger sequencing using the above second PCR primers.
Cell lines
The murine fibroblast NIH3T3 cell line was maintained in Dulbecco's modified Eagle's medium (DMEM; Wako Pure Chemicals) supplemented with 5% bovine serum (BS; Gibco, Thermo Fisher) and 1% penicillin/streptomycin (Wako Pure Chemicals). The 293FT cell line (Invitrogen, Thermo Fisher) was maintained according to the manufacturer's protocol. For serum starvation, DMEM with 0.5% FBS was used for 293FT cells.
Retrovirus vectors
Full-length coding portions of SQSTM1/NTRK3, AFAP1L2/RET, and PPFIBP2/RET were amplified from each cDNA sample generated in a previous work (18) and subcloned into pDON-AI (Takara Bio) with a C-terminal V5-tag. The accuracy of the sequence of the inserts was confirmed by Sanger sequencing. Recombinant retroviruses were produced by introduction of the above plasmids into 293 10A-1 cells. The titer of the virus vector was checked using NIH3T3 cells.
Focus formation assay
NIH3T3 cells were infected with the appropriate retrovirus at multiplicity of infection (MOI) of 0.1 in the presence of 4 μg/mL of polybrene (Sigma–Aldrich). Infected cells were cultured for two weeks in the presence of 0.75 mg/mL G418 (Wako Pure Chemicals), and the number of transformed foci was counted under an inverted phase-contrast microscope.
Cell growth
NIH3T3 cells stably expressing EGFP-V5, SQSTM1/NTRK3-V5, AFAP1L2/RET-V5, and PPFIBP2/RET-V5 were plated in six-well plates and grown in medium supplemented with 0.5% BS (Gibco, Thermo Fisher), 1% penicillin-streptomycin, and 0.75 mg/mL of G418. At the indicated time points, cells were counted using a TC20 Automated Cell Counter (Bio-Rad Laboratories).
Western blotting
Protein samples were resolved by sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and transferred onto a polyvinylidene fluoride (PVDF) membrane (Millipore). After blocking and incubation with an appropriate primary antibody, the antigen–antibody complexes were visualized using a horseradish peroxidase–conjugated secondary antibody (Cell Signaling Technology) and a chemiluminescence system (Nacalai Tesque). Detection was performed using a LAS-3000 imaging system (Fujifilm). Primary antibodies were obtained from the following sources: anti-phospho-ERK (Thr202/Tyr204), anti-ERK, anti-phospho-MEK (Ser217/221), and anti-MEK from Cell Signaling Technology; anti-β-actin from Santa Cruz Biotechnology; and anti-V5 from Invitrogen.
Statistical analysis
Differences between groups were examined for statistical significance with analysis of variance followed by Tukey's post test. A p-value <0.05 was considered statistically significant.
Results
Cases and genetic analysis
In a previous study (18), mutational analysis for the presence of the following well-established oncogenes was performed in 63 Fukushima PTC samples: BRAFV600E
, RAS, RET/PTC1, RET/PTC3, and ETV6/NTRK3. Forty-three (68.3%) cases with the BRAFV600E
mutation, six (9.5%) with RET/PTC1, one (1.6%) with RET/PTC3, and four (6.3%) with ETV6/NTRK3 rearrangements. There were nine samples, however, in which no mutation could be identified. Then, qRT-PCR was performed to screen for novel fusion genes by comparing the expression levels between the extracellular domain (5′) and the kinase domain (3′) of ALK, NTRK1, NTRK3, and RET. A higher expression of the kinase domain suggests the possibility of a gene rearrangement. Representative images are shown in Supplementary Figure S1 (Supplementary Data are available online at
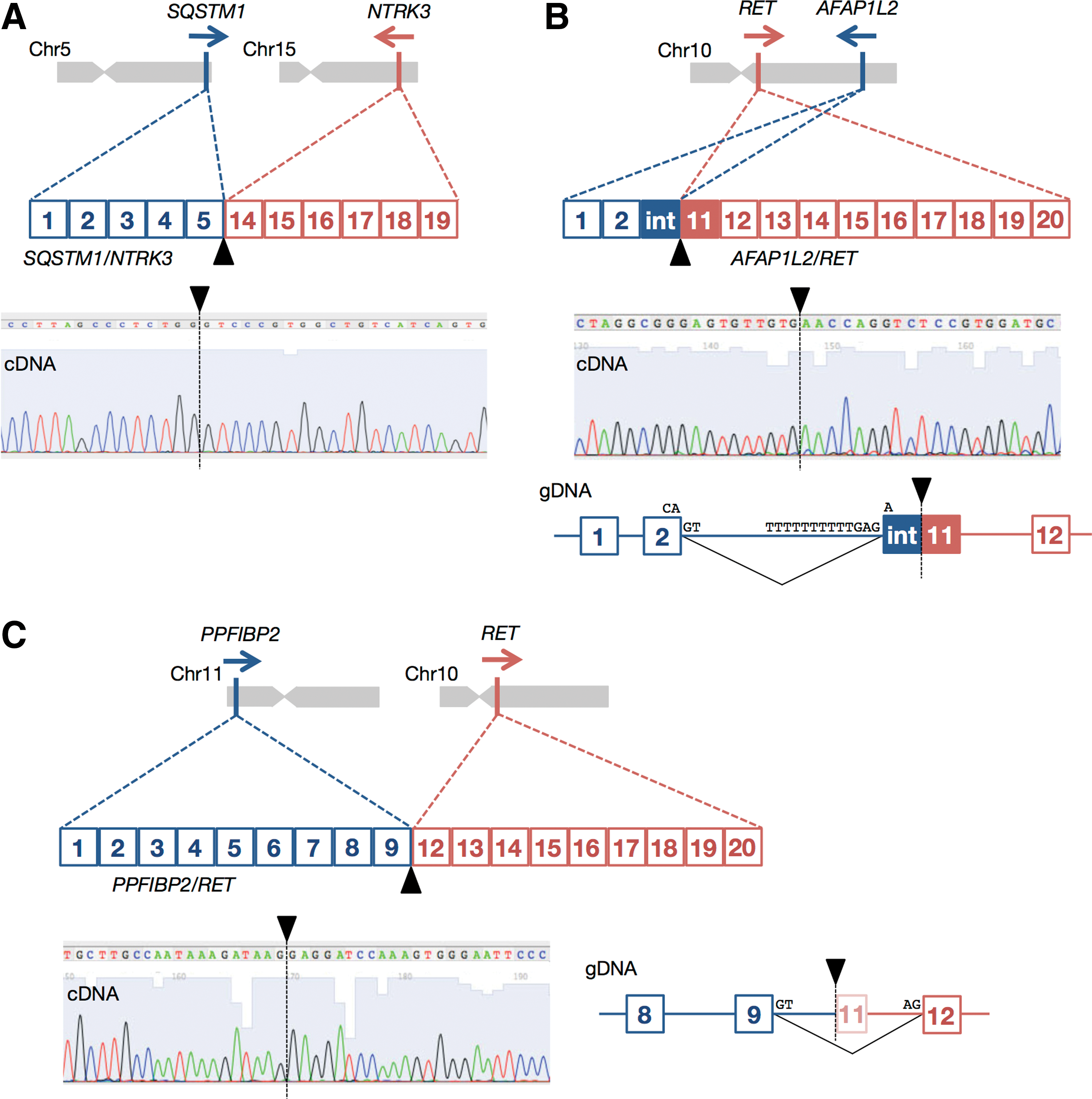
Genomic location, mRNA structure, sequence chromatogram of the breakpoint, and genomic DNA structure around the breakpoint of (
All three cases with novel fusion genes were histologically confirmed to be classical papillary adenocarcinomas without specific pathomorphological features or multifocality. The characteristics of the three patients were as follows: (i) SQSTM1/NTRK3: 18-year-old female, hemithyroidectomy, 7.0 mm in diameter, pT1a pN0 M0, no extrathyroidal extension; (ii) AFAP1L2/RET: 18-year-old male, total thyroidectomy, 40 mm in diameter, pT2 pN1b M1 (lung), no extrathyroidal extension; and (iii) PPFIBP2/RET: 13-year-old female, hemithyroidectomy, 15 mm in diameter, pT1b pN1a Mo, no extrathyroidal extension. None of the three patients had developed a recurrence when this article was initially submitted for publication.
Characteristics of the three novel fusion genes
SQSTM1/NTRK3 is a result of an interchromosomal translocation t(5;15)(q35.3;q25.3), which juxtaposes exons 1–5 of SQSTM1 to exons 14–19 of NTRK3 (Fig. 1A). The fusion point in NTRK3 is localized at the same position as in ETV6/NTRK3 (14). As the lengths of intron 5 of SQSTM1 and intron 13 of NTRK3 are 7805 bp and 93,252 bp, respectively, a breakpoint was not searched for on genomic DNA.
AFAP1L2/RET is a result of an intrachromosomal translocation t(10)(q25.3;q11.2), which fuses exons 1–2 and a part of intron 2 (38 bp) of AFAP1L2 to the 3′ part of exon 11 (33 bp) and exons 12–20 of RET in frame (Fig. 1B). This created a new exon, and genomic breakpoints are located in intron 2 and exon 11 of AFAP1L2 and RET, respectively. As an acceptor site at the 5′ end of exon 11 was missing, the genomic sequence TTT TTT TTT TGA G-A in intron 2 was presumably recognized as an acceptor (Fig. 1B).
PPFIBP2/RET is a result of an interchromosomal translocation t(11;10)(p15.4;q11.2), which juxtaposes exons 1–9 of PPFIBP2 to exons 12–20 of RET (Fig. 2C). Although breakpoints are located in intron 9 and in the middle of exon 11, the part of exon 11 was spliced out. In both latter cases, the kinase domain of RET is fully preserved as in conventional RET/PTC rearrangements.
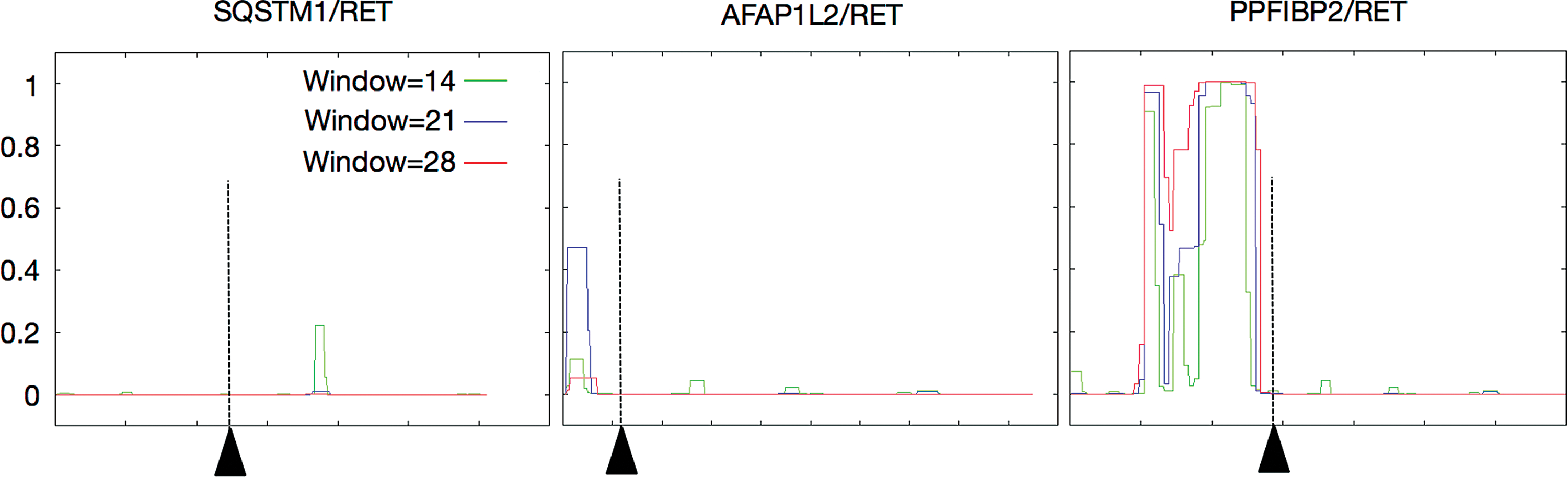
In silico prediction of a coiled-coil domain. Amino acid sequences of SQSTM1/NTRK3, AFAP1L2/RET, and PPFIBP2/RET were entered into the COILS server. Plotted is the probability of a coiled-coil domain existence at three different scanning windows. The breakpoints are indicated as arrowheads and dotted lines. Color images available online at
As these types of fusion proteins are usually activated through dimerization by a domain such as a coiled-coil domain in the N-terminal partner protein, in silico prediction analysis was performed using the COILS Server (
Functional studies
First, the effects of SQSTM1/NTRK3, AFAP1L2/RET, and PPFIBP2/RET on the MAPK signaling pathway were examined. For this purpose, these fusion proteins were overexpressed in 293FT cells (Fig. 3A), and the phosphorylation status of MEK and ERK was assessed by immunoblotting. As shown in Fig. 3B, ERK phosphorylation was induced by all three novel rearrangements, indicative of their ability to activate the MAPK pathway constitutively. AKT phosphorylation was also checked, but a significant change induced by these fusion proteins was not observed.
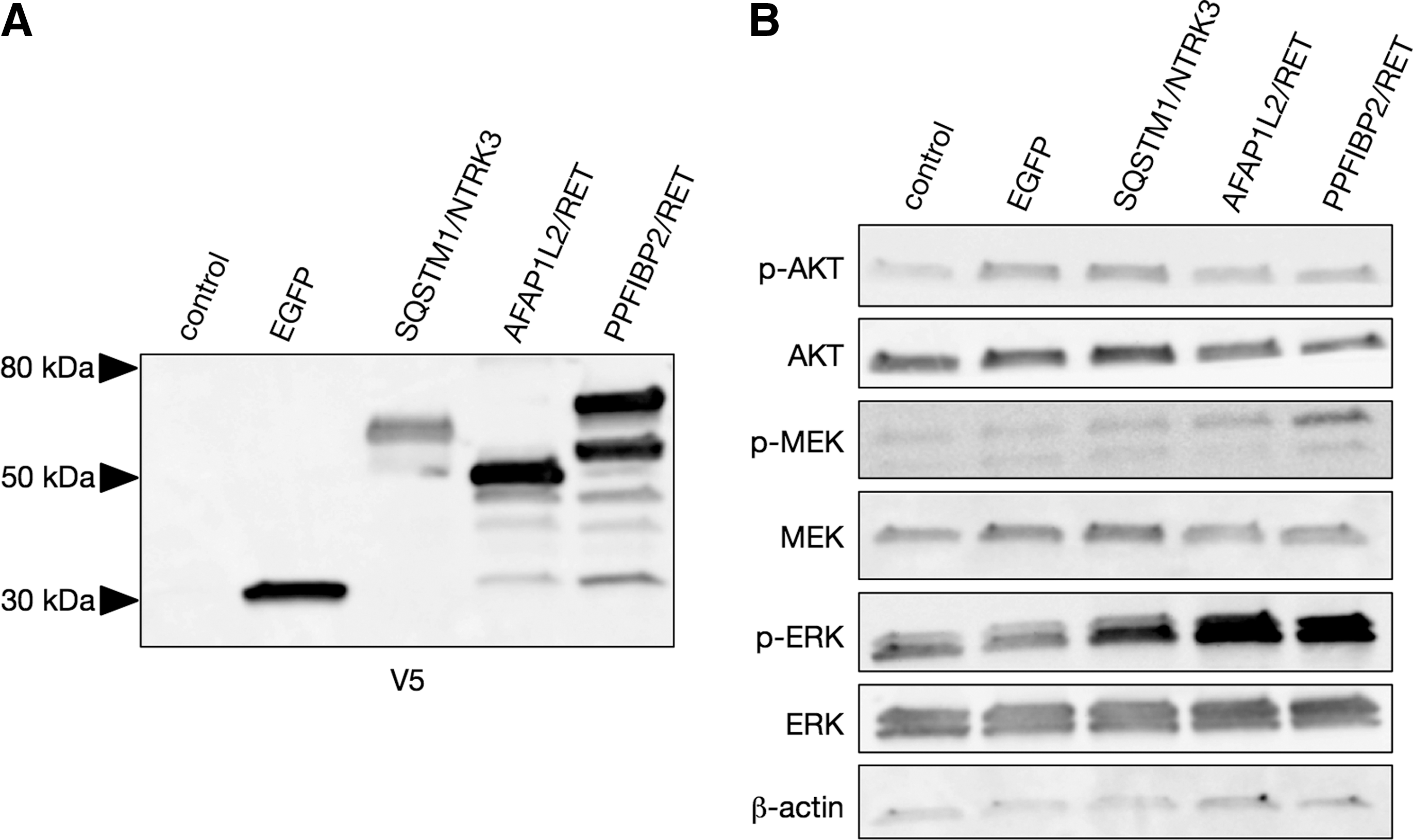
Effect of the novel fusions on signaling pathways. (
Next, we compared cell growth under low serum concentration. In this condition, EGFP-expressing control cells exhibited modest growth. In contrast, transduction with SQSTM1/NTRK3, AFAP1L2/RET, and PPFIBP2/RET significantly promoted cell growth (Fig. 4A). Note that the cells with SQSTM1/NTRK3 grew faster than the other two RET-carrying cells (Fig. 4A).
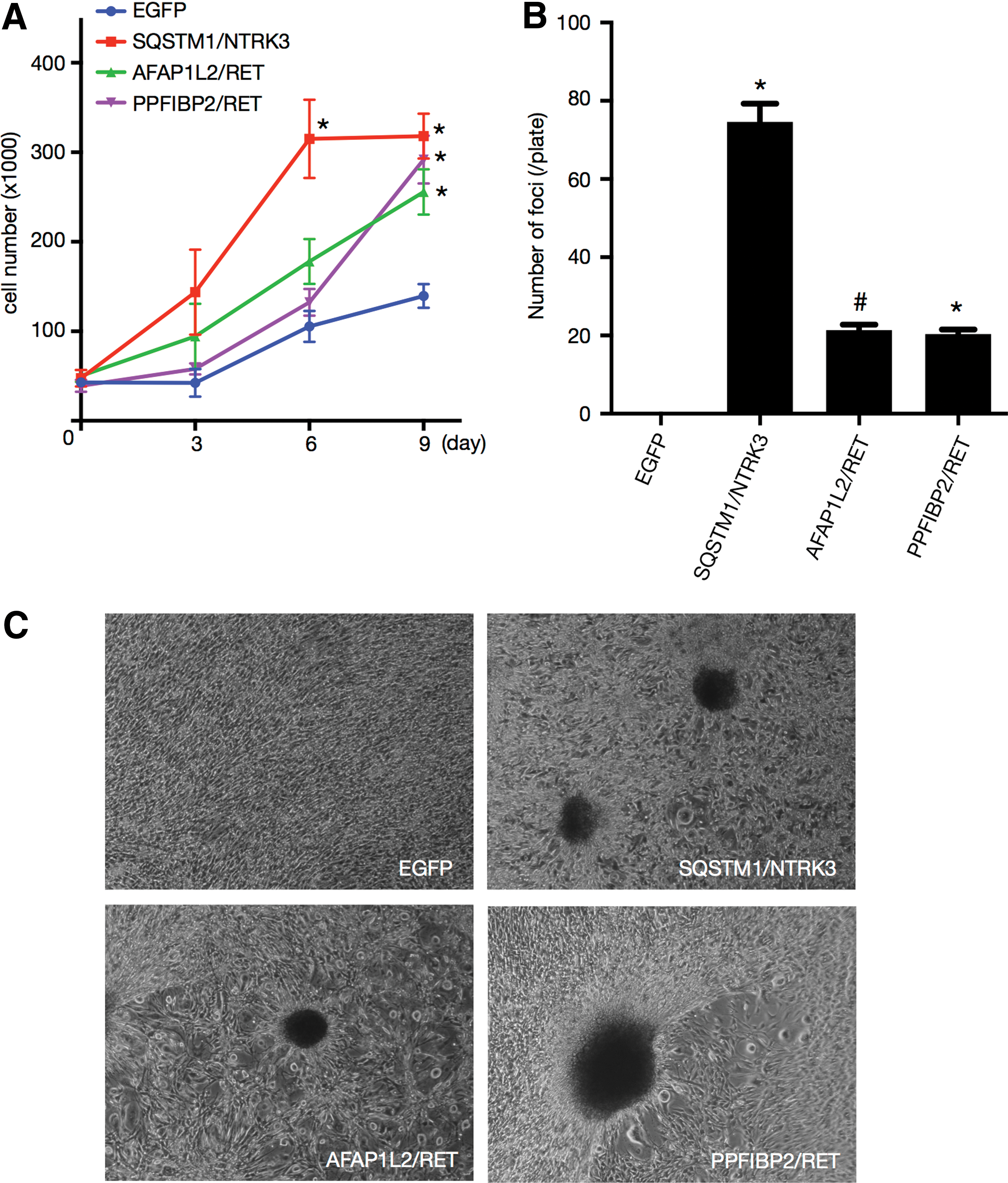
Transforming potential of the novel fusions. NIH3T3 cells were transduced with the indicated retroviruses. (
Then, a focus formation assay was performed to confirm the transforming potential of each of the novel fusion genes. A number of foci were successfully observed in NIH3T3 cells stably transduced by SQSTM1/NTRK3, AFAP1L2/RET, and PPFIBP2/RET (Fig. 4B and C). In contrast, overexpression of EGFP (control) did not result in the formation of any foci (Fig. 4B and C), and the number of foci was statistically different for all fusions compared with the control (Fig. 4B). Again, note that the number of foci after transduction with SQSTM1/NTRK3 was greater than with AFAP1L2/RET and PPFIBP2/RET (Fig. 4B).
These results indicate that the newly discovered three fusions, SQSTM1/NTRK3, AFAP1L2/RET, and PPFIBP2/RET, represent novel driver oncogenes.
Discussion
This study discovered three novel fusion oncogenes, SQSTM1/NTRK3, AFAP1L2/RET, and PPFIBP2/RET, which have a transforming ability through MAPK activation according to the in vitro studies. Taking into consideration previous results of molecular analysis in 63 young PTC patients from Fukushima (18) in which 43 (68.3%) cases with BRAFV600E and 11 (17.5%) with fusion genes were identified, the number of gene rearrangement-positive cases is now increased to 16 (25.4%) cases. In total, 59/63 (93.7%) cases were confirmed to have a driver mutation; only four (6.3%) cases remain negative for any oncogene so far.
The oncogenic potential of the newly discovered fusion genes was analyzed. First, SQSTM1/NTRK3 is a fusion between SQSTM1 on chromosome 5 and NTRK3 on chromosome 15. NTRK3 is a transmembrane receptor tyrosine kinase, whose ligand is neurotrophin-3. Recently, other SQSTM1 fusion genes have been identified in hematological malignancies: SQSTM1/ALK in large B-cell lymphoma, SQSTM1/NUP214 in T-cell acute lymphoblastic leukemia, and SQSTM1/FGFR1 in myelomonocytic leukemia (22 –25). SQSTM1/ALK has also been shown to have a transforming ability by focus formation assay using 3T3 fibroblasts (23). Although SQSTM1 does not have a coiled-coil domain, there is a Phox and Bem1p (PB1) domain at the N-terminus, which enables formation of heteromeric and homomeric complexes (26). Presumably, SQSTM1/NTRK3 is constitutively activated through dimerization mediated by the PB1 domain.
The SQSTM1 gene product is involved in several intracellular signal transduction cascades comprising a signaling node for multiple pathways maintaining cellular homeostasis. SQSTM1 may functionally contribute to aging, autophagy, and the development of degenerative diseases (27). How the disruption of one copy of SQSTM1 may contribute to tumor growth remains unclear. Note that mutations in the C-terminus of SQSTM1 have been reported to cause Paget's bone disease (28). Although the SQSTM1/NTRK3-expressing NIH3T3 cells showed a higher growth rate and transformation than the other two RET fusions, it cannot definitely be concluded that this fusion has a distinct malignant potential compared with the other two rearrangements because of the intrinsic limitations of the assay systems.
The two other fusion oncogenes, AFAP1L2/RET and PPFIBP2/RET, are new variants belonging to the RET/PTC family. The RET gene was originally identified as a proto-oncogene >30 years ago (29), and then a rearrangement named RET/PTC was found in PTC (30,31). At present, >15 types of RET/PTC rearrangements with different partner genes are known (32), to which the present work adds two more. RET encodes a transmembrane receptor tyrosine kinase; binding of a ligand, glial cell line–derived neurotrophic factor (GDNF) stimulates receptor dimerization, which is a critical step for gaining tyrosine kinase activity. Although the expression of RET in thyroid follicular cells is very limited, the fusion proteins are commonly expressed in thyroid follicular cells and possess coiled-coiled, leucine zipper, lis homology (LisH), or other domains that enable homo-dimerization of RET/PTC fusion proteins. As a result, RET/PTC gene products are constitutively activated without ligand binding in thyroid cells.
The novel partner gene AFAP1L2 is located on chromosome 10 and belongs to the actin filament-associated protein (AFAP) family. Although AFAP1L2 is expressed in the human thyroid as well as in other organs, the role of AFAP1L2 in thyroid carcinogenesis is not yet formally clarified (33). In AFAP1L2/RET, it was found that only exons 1–2 were fused to RET, but according to in silico analysis, a coiled-coil domain likely exists in this portion and enables dimerization, leading to constitutive activation of the RET kinase. All of the results of this functional study suggest that AFAP1L2/RET is a novel oncoprotein. However, as the breakpoint of this fusion is very unique (combining intronic sequences and an exon), this is probably a rare event.
Finally, another newly discovered partner gene is PPFIBP2. It is located on chromosome 11, and encodes the protein-tyrosine phosphatase receptor-type f-polypeptide (PTPRF)-binding protein. The PPFIBP2 product plays a role in axon guidance and neuronal synapse development; its function in the thyroid is unknown. Of note, rs12791447 SNP in the PPFIBP2 intron was found to be a susceptibility marker for prostate cancer (34). Also, elevated PPFIBP2 mRNA expression was detected in endometrial cancer (35), suggesting that it may be associated with cancer pathogenesis. Again, the in silico analysis revealed that exons 1–9 of PPFIBP2 highly likely harbor a coiled-coil domain and promote dimerization, leading to constitutive activation of RET.
The limitation of the present work consists in the lack of in vivo tumor formation studies. However, all driver oncogenes that have a focus-forming ability in NIH3T3 cells also formed tumors in immunodeficient mice in our hands.
In summary, three novel fusion oncogenes, SQSTM1/NTRK3, AFAP1L2/RET, and PPFIBP2/RET, were identified in the PTCs from one child and two adolescents from a cohort consisting of 63 members from Fukushima. The findings suggest that rare fusions may be present among the cases negative for known oncogenes in this age group and that they play a significant role in thyroid carcinogenesis. The results also indicate that almost all PTCs (93.7%) in young patients have oncogenic driver mutations activating the MAPK intracellular signaling pathway.
Footnotes
Acknowledgments
This work was supported in part by JSPS KAKENHI, grant number: 25257508 (S.Y.), 26293222 (S.Y.), 26293142 (N.M.), and 16K09804 (M.M.). This study was also supported in part by Cooperative Research Grant of Atomic Bomb Disease Institute, Nagasaki University (Sh.S.). We thank the members of the Departments of Thyroid and Endocrinology, and Organ Regulatory Surgery, Fukushima Medical University, the Thyroid Examination Unit of the Radiation Medical Centre for the Fukushima Health Management Survey, and the Consensus Meeting of Pathological Diagnosis of Fukushima Thyroid Cancers, who participate in the thyroid ultrasound screening program in Fukushima Prefecture.
Author Disclosure Statement
We declare no competing interests.