
Abstract
Background:
Myopathic changes are commonly described in hypothyroid and hyperthyroid patients, including muscular atrophy and weakness. Satellite cells (SCs) play a major role in skeletal muscle maintenance and regeneration after injury. A mouse model of resistance to thyroid hormone–TRα1PV demonstrated impaired skeletal muscle regeneration after injury with significant reduction of SCs, suggesting that exhaustion of the SC pool contributes to the impaired regeneration. To test this hypothesis, SC activation and proliferation were analyzed in vivo in response to skeletal muscle injury and during aging.
Methods:
SCs of TRα1PV male mice were analyzed four days after cardiotoxin-induced muscle injury, and they were compared to wild-type (WT) male animals. TRα-knockdown C2C12 myoblasts were injected into injured skeletal muscle, and four days after transplantation, the in vivo behavior was compared to control C2C12 myoblasts. Skeletal muscle regeneration was compared in younger and older TRα1PV and WT animals.
Results:
The total number of SCs in skeletal muscle of TRα1PV mice was significantly lower than control, both before and shortly after muscle injury, with significant impairment of SC activation, consistent with SC pool exhaustion. TRα-knockdown myoblasts showed impaired in vivo proliferation and migration. TRα1PV mice had skeletal muscle loss and significant impairment in skeletal muscle regeneration with aging. This translated to a significant reduction of the SC pool with aging compared to WT mice.
Conclusion:
TRα plays an important role in the maintenance of the SC pool. Impaired skeletal muscle regeneration in TRα1PV mice is associated with insufficient SC activation and proliferation, as well as the progressive loss of the SC pool with aging. Regulation of the SC pool and SC proliferation provides a therapeutic target to enhance skeletal muscle regeneration and possibly slow age-associated sarcopenia.
Introduction
P
Both hyperthyroidism and hypothyroidism are associated with changes in skeletal muscle (6). Thyroid hormone signaling is crucial for skeletal muscle function, metabolism, and skeletal muscle repair after injury (7). The process of SC activation, proliferation, and differentiation is likely regulated by triiodothyronine (T3) at multiple steps. In fact, the expression of the skeletal muscle transcription factors MyoD and myogenin are regulated by T3 (8,9).
Skeletal muscle knockdown of Dio2, the enzyme that converts the prohormone thyroxine to the active form T3 (10), or the T3 inactivating enzyme Dio3 (11), leads to impaired skeletal muscle regeneration, indicating the importance of ligand availability. Skeletal muscle isolated from thyroid hormone receptor alpha (TRα) and -β knockout mice show a switch from type 2 fast muscle fibers to type 1 slow muscle fibers (12,13). It was recently reported that myoblasts with TRα knockdown, or derived from TRα PV (a frame-shift mutation associated with resistance to thyroid hormone) mice, displayed reduced proliferation and myogenic differentiation that could be explained in part by the TRα modulation of the Wnt/β catenin pathway (14). Moreover, skeletal muscle from the TRα1PV mutant mouse had impaired in vivo regeneration after injury (14). These data indicate that TRα plays an essential role in myoblast homeostasis and provides a potential therapeutic target to enhance skeletal muscle regeneration.
Materials and Methods
Animal models and muscle injury
The generation of the TRα1PV mouse has been previously described (15). The study used 8- and 20-month-old male TRα1PV and wild-type (WT) male mice, and injected cardiotoxin (CTX, 25 μL [0.03 mg/mL]; Sigma–Aldrich) into the right tibialis anterior muscle (TAM), or in the quadriceps femoris, to induce muscle injury. The left TAM or left quadriceps femoris was used as an internal control, and a volume of saline equal to the cardiotoxin was injected. The mice were euthanized at day 4 or 14 after skeletal muscle injury. Skeletal muscle was collected, embedded in paraffin, and sectioned for histopathological analysis. The early time point (day 4) was used to investigate early SC activation and proliferation, and the later time point (day 14) was used to study SC differentiation and contribution to skeletal muscle regeneration.
All animal procedures were performed in accordance with approved protocols by the Institutional Animal Care and Use Committee at VA Greater Los Angeles Healthcare System and Children's Hospital Los Angeles.
Cell culture, transfection, and transplantation
C2C12 cells (ATCC) were cultured in Dulbecco's Modified Eagle's Medium and 15% fetal bovine serum and maintained at sub-confluent density. Cells were plated in a six-well dish and transfected with TRα shRNA plasmid (Santa Cruz Biotechnology) and control shRNA plasmid, using the recommendations of the manufacturer. To induce stable transfection, transfected cells were selected with puromycin (2 μg/mL). Polymerase chain reaction and Western blotting was used to verify efficiency of the knockdown, as previously shown (14).
NOD/SCID mice (Jackson Laboratories), six to eight weeks of age, were injected with 25 μL (0.03 mg/mL) of CTX in the right TAM to induce muscle injury. The left TAM was used as an internal control. On day 2, about 1 × 105 C2C12 cells were injected into the pre-injured muscles: TRα knockdown C2C12 cells (shTRα) and control C2C12 cells transfected with scrambled RNA (shRNA). The cells were labeled with 1,1′-dioctadecyl-3,3,3′3′-tetramethylindocarbocyanine percholorate (DiI) to facilitate in vivo cell tracking. This alipophilic fluorescent carbocyanin dye remains stable for several days in living culture, as well as undergoes negligible transfer between intact membranes. The mice were euthanized on day 4 after the cell injection, and the TAMs were collected, embedded in paraffin, and sectioned for histopathological studies.
Immunohistochemistry
Skeletal muscle was harvested at 4 and 14 days after skeletal muscle injury, and after four days from SC transplantation in the pre-injured skeletal muscle, and fixed overnight with 4% paraformaldehyde at 4°C. The tissue was then dehydrated in graded ethanol, cleared in xylene, and embedded in paraffin. For immunohistochemical staining of the paraffin-embedded samples, sections were de-paraffinized in xylene and rehydrated through ethanol baths and phosphate-buffered saline (PBS), followed by rinsing in distilled water for five minutes. The following primary antibodies were used: mouse monoclonal antibodies anti-PAX7 (Santa Cruz) at a dilution of 1:100, rabbit polyclonal antibodies anti Ki67 (Abcam) at dilution 1:100, and rabbit polyclonal anti-laminin (Abcam) at a dilution 1:100. After washing with PBS, detection of bound primary antibodies was carried out with appropriate secondary antibodies conjugated with Alexa Fluor 488 or 568 (Life Technologies). Nuclear DNA was counterstained with 4′,6-diamidino-2-phenylindole (DAPI; Vector Lab).
Confocal microscopy and image quantification
Scanning confocal images for immunofluorescence analysis were obtained by a laser scanning confocal microscope (Leica Microsystems SP5). Skeletal muscle fiber size was measured with an Aperio Scanscope AT Turbo, and the measurements were analyzed using Aperio ImageScope software (Leica Biosystems). Digital images of random 0.15 mm2 fields were taken, and labeled cells were counted by hand. Five 0.15 mm2 fields were randomly chosen for each sample for statistical analysis. Image analysis was performed using ImageJ software. Skeletal muscle fiber size was analyzed by measuring the lesser diameter (minimal Feret diameter) of muscle fibers.
Statistical analysis
Values are presented as mean ± standard deviation (SD). For all quantitative analyses, a minimum of three replicates were performed in independent experiments or individual mice. Significant difference between means was determined using a two-tailed Student's t-test or one-way analysis of variance followed by the Student–Newman–Keuls test.
Results
TRα is essential for in vivo SC activation and proliferation in response to injury
The total number of Pax7-positive SCs in the skeletal muscle of eight-month-old TRα1PV mice was significantly lower than in controls both before and four days after muscle injury (Fig. 1A–D and Supplementary Fig. S1; Supplementary Data are available online at

Satellite cell (SC) activation four days after skeletal muscle injury (4 dpi) in eight-month-old wild-type (WT) mice (
TRα in SCs is important for in vivo engraftment, proliferation, and migration after injury
TRα was inactivated in a murine myoblast cell line, widely used as SC line, as previously reported (14). The ability of C2C12 cells, after TRα knockdown, to engraft, proliferate, and migrate in vivo was evaluated (Fig. 2). Then they were compared to control C2C12 cells transfected with shRNA (Control RNA). The efficiency of TRα knockdown in C2C12 cells using this method has been shown previously (14). Donor cells were labeled with DiI to facilitate in vivo cell tracking (Fig. 2A). Five days after cardiotoxin injection, the skeletal muscle showed an area of inflammation with destruction of the normal fiber architecture (injury area) and a surrounding area with normal skeletal muscle fibers (normal adjacent area; Fig. 2B). Four days after transplantation, the mice injected with Control-C2C12 cells showed an efficient engraftment of donor cells in the injured muscle (Fig. 2C, central panels). An abundance of cells were also observed in the sub-laminar space, adjacent to the muscle fiber (Fig. 2C, arrows), indicating that the control C2C12 cells migrated to the normal anatomical location for SCs. In contrast, TRα knockdown (shTRα) C2C12 cells were reduced in number at the level of the injured muscle, and only a few cells migrated and localized to the sub-laminar space (single arrow, Fig. 2C, right panels, and Fig. 2D). This indicates that TRα is essential for in vivo engraftment, proliferation, and migration/localization in the satellite stem-cell compartment.
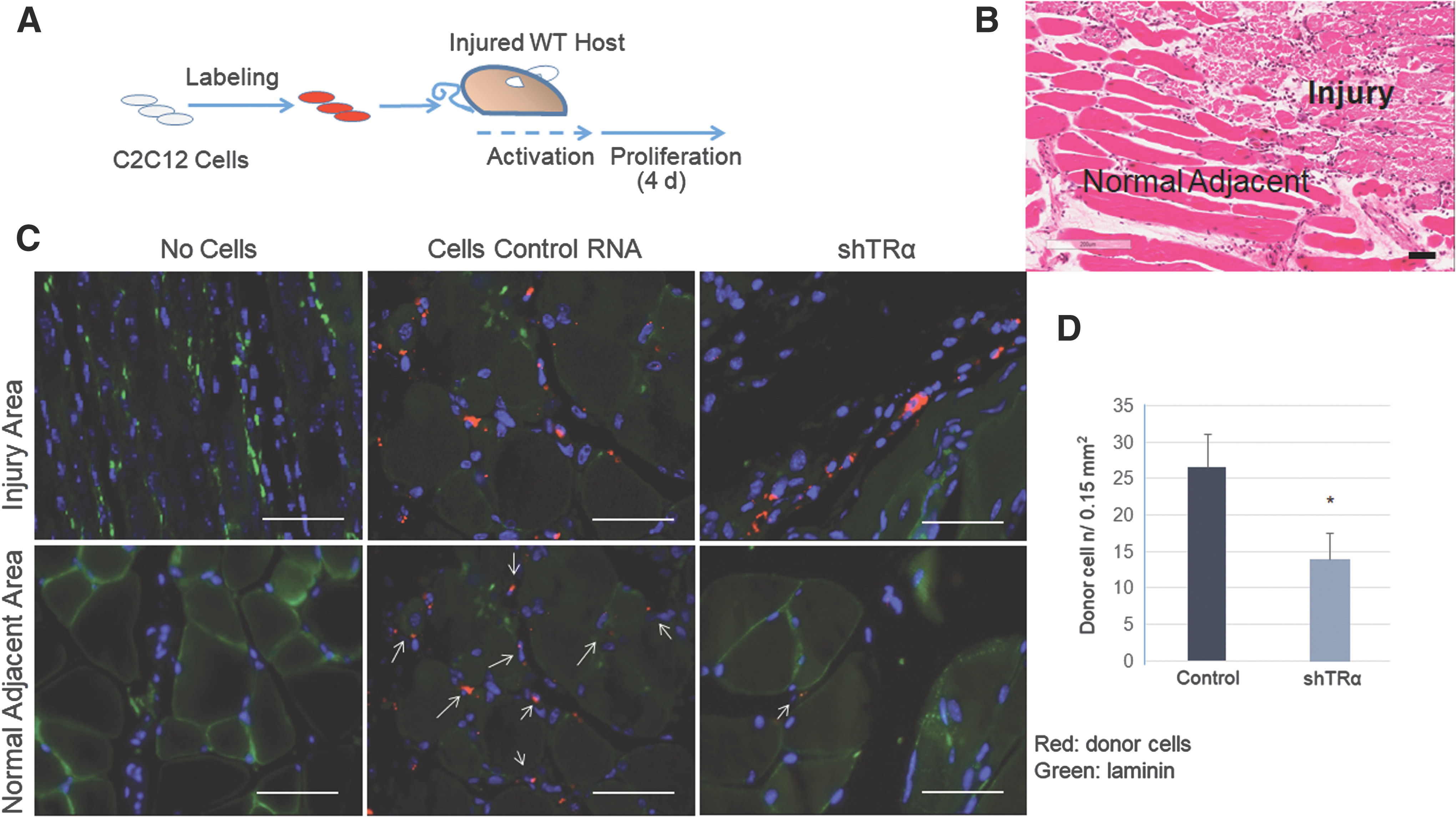
Transplantation of skeletal muscle stem cells (C2C12) into mouse host after tibialis anterior muscle (TAM) injury. (
TRα plays an important role in maintenance of muscle mass and skeletal muscle regeneration after injury with aging
It has been shown that myofibers from eight-month-old TRα1PV mice, compared to WT mice, were smaller before and two weeks after inducing skeletal muscle injury with cardiotoxin, a standard muscle injury technique (14). TRα1PV mice, compared to WT mice, showed reduced muscle fiber cross-sectional area, an indication of skeletal muscle regeneration, four days after CTX injection in the TAM. These findings indicate impaired skeletal muscle regeneration associated with the TRα mutation (14). To investigate further the role of TRα in muscle maintenance with aging, 20-month-old TRα1PV mice were compared to WT age-matched control animals. Skeletal muscle mass in C57bl/6 mice has been reported to increase from 3 to 18 months of age, but then the muscle mass progressively decreases (16). It has also been reported that after 24 months of age, skeletal muscle regeneration after injury is impaired in normal mice (17). The skeletal muscle fiber size of the 20-month-old TRα1PV mice was evaluated by measuring the minimal fiber Feret of the muscle fiber in TAM (Fig. 3). There was a significantly reduced minimal fiber Feret in the TAM of 20-month-old TRα1PV mice compared to age-matched WT mice (Fig. 3A and B), indicating significantly lower muscle mass, similar to what was previously shown in younger TRα1PV mice (14). Two weeks after inducing skeletal muscle injury with CTX, regenerating muscle fibers from 20-month -old TRα1PV mice were significantly smaller compared to regenerating muscle fibers from WT mice (Fig. 3). Immunostaining for skeletal muscle laminin, 14 days after CTX-induced injury, showed an immature skeletal muscle phenotype in all mice (TRα1PV and WT), consisting of smaller muscle fibers in the regenerating muscle and centrally located nuclei (Fig. 3A). However, the reduction in minimal fiber Feret observed 14 days after CTX in TAM was significantly more pronounced in TRα1PV mice compared to WT mice.
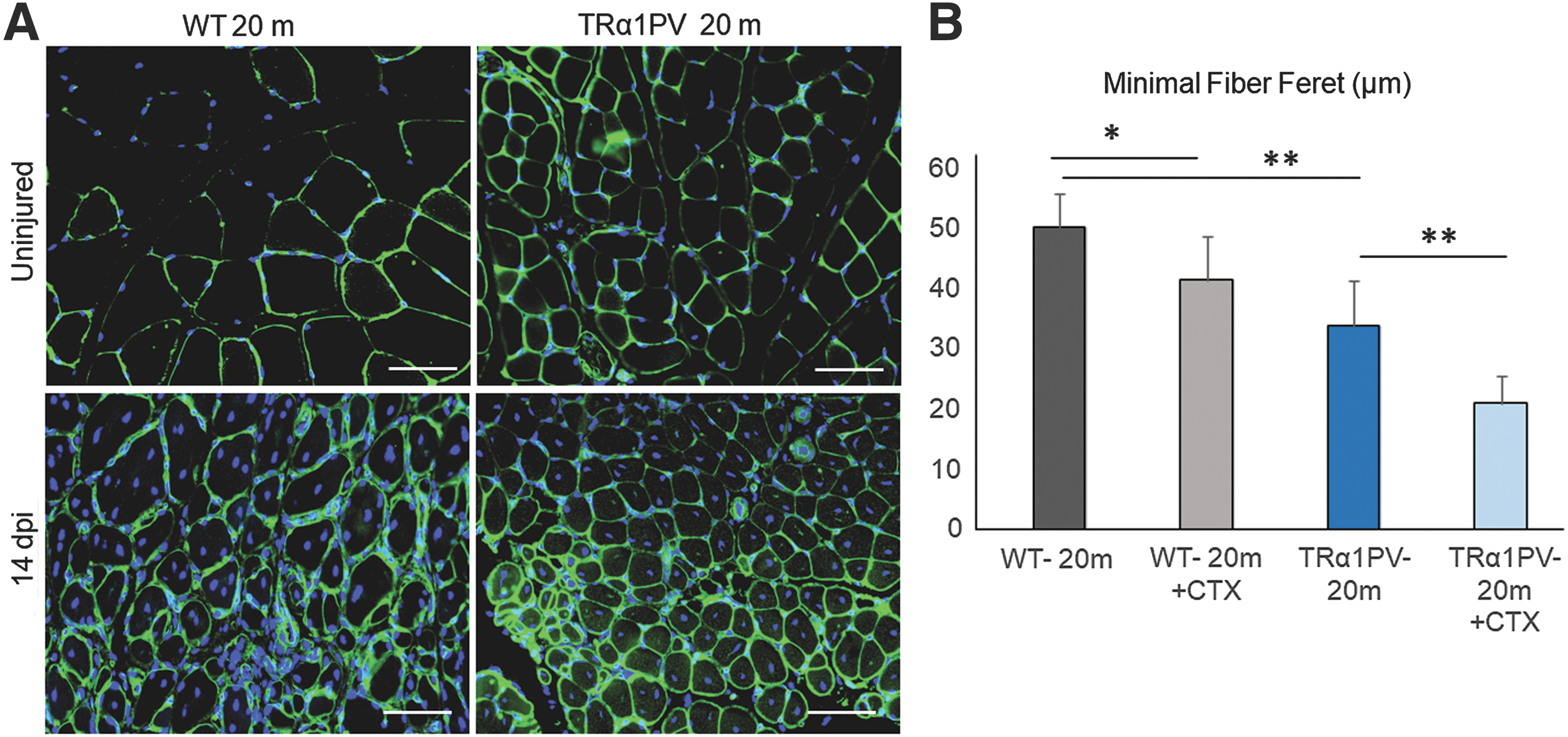
Significant impaired skeletal muscle regeneration in older TRα1PV mice. (
Compared to younger siblings (eight months old), WT animals had a significant increase in minimal fiber Feret from 8 to 20 months of age (Fig. 4A), consistent with previous reports (16). In contrast to these findings in WT mice, the TRα1PV mice showed a blunted and nonsignificant increase in minimal fiber Feret from 8 to 20 months compared to control mice (Fig 3B), consistent with a failure to achieve peak muscle mass (Fig. 4A).

The influence of age on skeletal muscle regeneration after injury in WT and TRα1PV mice. Skeletal muscle regeneration 14 days after CTX-induced muscle injury (14 dpi) in WT and TRα1PV mice. A minimal fiber Feret (in μm) in uninjured TAM from WT and TRα1PV mice at 8 and 20 months of age. (
Moreover, if the loss in minimal fiber Feret 14 days after CTX-muscle injury is compared at different time points (8 and 20 months of age), 20-month-old TRα1PV mice have a significant deficit in skeletal muscle regeneration after injury compared to the younger eight-month-old siblings. This was shown by a loss of 45% of the minimal fiber Feret after injury at 20 months compared to a loss of 25% at eight months (Fig. 4B). Interestingly, WT mice did not show worsening of skeletal muscle regeneration after injury at 20 months compared to 8-month-old siblings (Fig. 4B). This indicates a worsening of skeletal muscle regeneration capacity over time in TRα1PV but not WT mice.
Loss of skeletal muscle mass with aging in TRα1PV mice is associated with loss of SC pool
The study further investigated the role of SC in the loss of muscle mass in TRα1PV mice. The number of Pax7-positive SCs was significantly lower in the 20-month-old TRα1PV mice compared to age-matched WT animals (Fig. 5). This confirmed the previous observation of reduced SCs in younger TRα1PV mice (14). Interestingly, when Pax7-positive SCs were compared in young (eight months old) and older mice (20 months old), a significant decrease in Pax7 expression was consistently found with aging in TRα1PV mice but not in the WT animals (Fig. 5B). Skeletal muscle from 20-month-old WT mice showed the same degree of Pax7 expression as the skeletal muscle from 8-month-old WT mice. In contrast, skeletal muscle from 20-month-old TRα1PV mice showed a significant reduction in Pax7 expression compared to the skeletal muscle of eight-month-old TRα1PV mice (Fig. 5B). This suggests a progressive loss of the SC pool in TRα1PV skeletal muscle with aging.
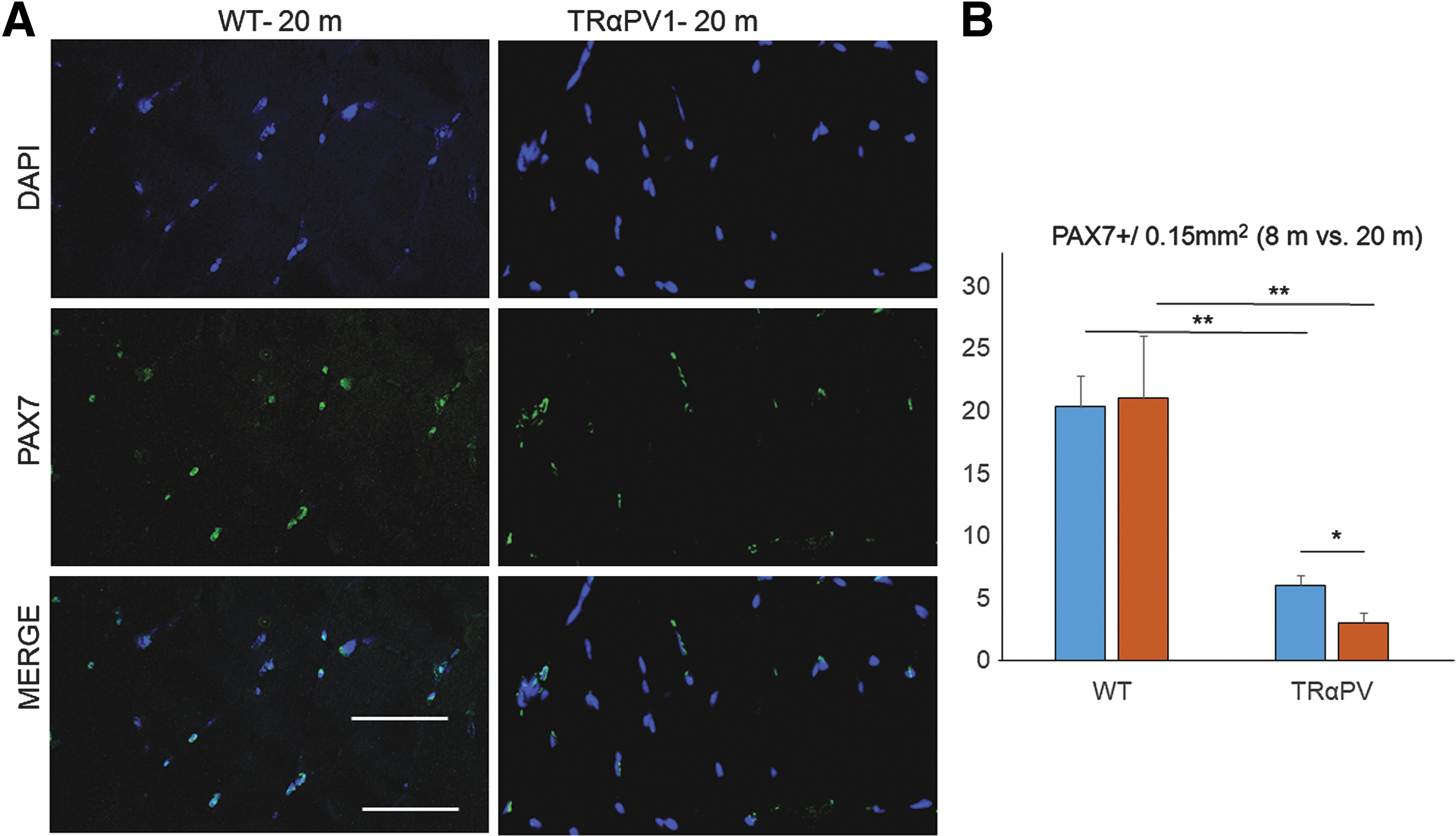
Influence of age on PAX7-positive SCs. (
Discussion
The self-renewing and proliferation of SCs leads to new myofiber formation and is crucial for the maintenance of the SC pool. Pax7 coordinates self-renewal and maintenance of the SC niche and is required to instruct the myogenic fate of SCs. Impaired skeletal muscle regeneration was recently reported in a mouse model of RTH-TRα1PV (14). One salient finding was a significant reduction of Pax7-positive SCs in the skeletal muscle of TRα1PV mice, suggesting that the exhaustion of the SC pool was the primary reason for the impaired skeletal muscle regeneration. Moreover, after knocking down TRα in C2C12, a myoblast mouse cell line that has been widely used as a SC model, Pax7 expression was significantly reduced compared to control C2C12 cells (14). Both TRα knockdown cells and TRα1PV myoblasts showed impaired in vitro proliferation and differentiation (14). Here, the role of TRα in SC maintenance, activation, and proliferation was investigated in models of aging and injury.
To investigate CS activation and proliferation, an in vivo model of skeletal muscle injury was used. SCs are located close to the muscle fiber and are poised to be rapidly activated in response to injury (2). After activation, SCs enter the cell cycle, migrate from their niche, proliferate, and differentiate (2). This study shows that SC activation immediately after muscle injury (four days after) is blunted in TRα1PV mice. Moreover, knockdown of TRα in C2C12 cells is associated with reduced cell migration and proliferation in the injured muscle four days after transplantation in the injured muscle. These could, in part, explain the impairment in skeletal muscle regeneration in TRα1PV mice (14).
To investigate SC maintenance, skeletal muscle atrophy with aging was examined. Degenerative skeletal muscle states, sarcopenia secondary to motor nerve denervation, advanced age, deconditioning, or immobilization are associated with a reduction in SC number and proliferative potential (18). This contributes to a reduction in skeletal muscle regenerative capacity and contractility.
The number and function of skeletal muscle SCs declines with age and in aged mice, and muscle repair is blunted because of SC dysfunction (17,19 –21). After a finite number of divisions, SCs enter into an irreversible arrest in the G1 phase of the cell cycle, called replicative senescence (22). It was found that TRα1PV mice had early muscle atrophy and a deterioration of skeletal muscle repair with aging. Muscle atrophy is the result of a protein degradation rate that exceeds protein synthesis (23), and results in compromised skeletal muscle regeneration and muscle stem-cell function (17).
The TRα1PV mice failed to increase skeletal muscle mass from 8 to 20 months of age compared to age-matched WT animals. Moreover, after skeletal muscle injury, 20-month-old TRα1PV mice showed a more profound and significant impairment in skeletal muscle regeneration compared to the younger (8-month-old) siblings, indicating progressive loss of skeletal muscle mass and regeneration. Although several possible mechanisms could explain the impaired muscle regeneration, the study focused on the role of SCs. In the TRα1PV mice, the SC activation and proliferation immediately after skeletal muscle injury was significantly impaired compared to control sibling mice. Moreover, Pax7 expression, which represents a marker for SC number, was reduced in the younger animal compared to the WT mice, and this reduction in number was more pronounced and significant in the older 20-month-old mice. Taken together, these data indicate that skeletal muscle loss and impaired regeneration in TRα1PV mice can, in part, be explained by the loss of SC number and function.
The question that still remains unanswered is how TRα influences SC behavior. The in vivo behavior of SC in the TRα1PV mice cannot distinguish direct from indirect effects of TRα on the SC niche, and it must be kept in mind that other systemic factors can affect SC self-renewal and proliferation. However, previous data utilizing in vitro culture of myoblasts from TRα1PV mice and C2C12 cells with knockdown of TRα showed a reduction in both proliferation and differentiation, suggesting a cell-intrinsic mechanism (14). In addition, the in vivo behavior of TRα knockdown C2C12 cells after transplantation in a normal injured muscle indicates that a direct role for TRα in SC regulation is likely.
In summary, the present data indicate that TRα plays an important role in the maintenance of the SC pool. Impaired skeletal muscle regeneration in TRα1PV mice may be explained by insufficient SC activation and proliferation, and progressive loss of the SC pool. Regulation of SC proliferation provides a therapeutic target to enhance skeletal muscle regeneration.
Footnotes
Acknowledgments
This study was supported by NIH K08DK097295 (A.M.), VA Merit Review (A.M.), NIH RO1DK98576 (G.A.B.), and VA Merit Review (G.A.B.). The research described was supported by NIH/National Center for Advancing Translational Science (NCATS) UCLA CTSI Grant Number UL1TR000124.
Author Disclosure Statement
No competing financial interests exist.