
Abstract
Background:
Epigenetic factors associated with the development of autoimmune diseases are unclear. Monozygotic twin pairs discordant for positive antithyroglobulin autoantibodies (TgAb) are useful to examine the epigenetic factors because of their identical genetic background. This study aimed to clarify the discordant epigenetic differences affecting the development of TgAb.
Methods:
Subjects were selected from 257 Japanese monozygotic twins, recruited from the registry established by the Center for Twin Research at Osaka University. TgAb positive concordant (PC) pairs were 5.7% (four pairs) and 9.6% (18 pairs) of male and female pairs, respectively. TgAb discordant (DC) pairs were 11.4% (eight pairs) and 8.0% (15 pairs) of male and female pairs, respectively. TgAb negative concordant (NC) pairs were 78.6% (55 pairs) of male pairs and 74.3% (139 pairs) of female pairs. To perform stricter grouping, the cut-off value for positive TgAb was set to 50.0 IU/mL (TgAb negative: <28.0 IU/mL; TgAb positive: ≥50.0 IU/mL; TgAb borderline: ≥28.0 IU/mL and <50.0 IU/mL). Nineteen discordant (6 male and 13 female pairs) and 185 concordant pairs (48 male and 137 female pairs) for TgAb positivity were finally examined. DNA methylation levels of genomic DNA were evaluated using the Infinium HumanMethylation450 BeadChip Kit. Gene polymorphisms were also genotyped using the Omni5-4 BeadChip Kit to clarify genetic background specific for discordant twins.
Results:
No CpG sites were found with significant within-pair differences of methylation levels in TgAb DC pairs after correction for multiple comparisons. However, 155 polymorphisms specific for TgAb DC pairs were significantly different in genotype frequencies from those of concordant pairs, and none of them were located on the HLA region of chromosome 6. In TgAb DC pairs with some specific genotypes of these polymorphisms, four CpG sites were observed exhibiting significant within-pair differences in each DC pair, even after correction for multiple comparisons.
Conclusions:
This study found that the genetic background specific for TgAb DC twins who are susceptible to epigenetic changes are different from that specific for TgAb PC twins, and it clarified the genotype-based epigenetic differences in TgAb DC monozygotic twins.
Introduction
T
Monozygotic and dizygotic twins are very important resources to evaluate the relative contribution of genetic and environmental factors to phenotypes of a certain disease. Previous twin studies have reported that thyroid autoimmunity may be under strong gene influence and environmental factors (7,8). Model-fitting analysis showed that 79% of the liability of Graves' disease development is attributable to genetic factors and 21% to individual-specific environmental factors (9). It is very difficult to identify environmental factors affecting the predisposition to thyroid autoimmunity in the general population. However, it is easy to identify the environmental factors when examined in monozygotic twins discordant for positive thyroid autoantibodies who are susceptible to environmental factors. This study focused on epigenetic changes of DNA in order to estimate the impact of environmental effects on the predisposition to thyroid autoimmunity.
Modifications to genomic DNA, such as DNA methylation and histone modification, are typical epigenetic factors, and they impact cellular behavior by regulating chromatin status and gene expression (10). One of the most important epigenetic modifications is methylation of genomic DNA, which is the covalent addition of a methyl group to the cytosine in C-phosphate-G (CpG) dinucleotides. The CpG sites present in the regions containing high numbers of CpG dinucleotides (CpG islands) are generally unmethylated, whereas those in most other genomic regions are methylated. CpG islands overlap with the promoter regions of 60–70% of genes and are generally protected from methylation, allowing for the expression of downstream genes that are further regulated by histone modification (11). It was supposed that aberrant DNA methylation of CpG sites may be one of the most important epigenetic changes affecting the developmental process of autoimmunity, leading to regulation of gene expression. Discordant twins for type 1 diabetes have been already analyzed, and the results suggest that some abnormalities in DNA methylation patterns may be involved in the pathogenesis of diabetes (12). With regard to thyroid autoimmunity, the genetic–epigenetic interaction involving a noncoding polymorphism in the THSR gene have been reported using cultured thyroid cells (13). However, the epigenetic differences, including DNA methylation, in discordant phenotypes of monozygotic twins with AITD have not been investigated.
This study investigated levels of antithyroglobulin autoantibodies (TgAb), the most sensitive tests to detect thyroid autoimmunity, and the methylation levels of CpG sites in monozygotic (identical) twins to clarify the association between autoantibody production and methylation patterns of genomic DNA. Moreover, twins were also genotyped to find the genomic backgrounds specific for the discordant pair of TgAb positivity to look for individual differences in the sensitivity to epigenetic changes related to TgAb development.
Subjects and Methods
Subjects
A total of 257 Japanese monozygotic twins (70 male and 187 female pairs) were recruited from the registry established by the Center for Twin Research at Osaka University (14). Written informed consent was obtained from all the twins, and the Ethics Committee of Osaka University approved the study protocol (no. 506). Blood samples were taken at 9:00am after a 12 hour fast. A clinical examination was performed, and the twins filled in health-related questionnaires, including questions regarding thyroid disease. The twins were examined on the same day. Genomic DNA was isolated from peripheral blood mononuclear cells using a commercial kit (QIAamp DNA Mini Kit; QIAGEN, Hilden, Germany). Zygosity of subjects was confirmed by perfect matching of 15 short tandem repeat (STR) loci using the PowerPlex® 16 System (Promega, Madison, WI).
TgAbs
TgAbs were examined with commercial electrochemiluminescence immunoassay (ECLIA) kits (ECLusys anti-Tg; Roche Diagnostics, Tokyo, Japan). The reference value for TgAb is <28.0 IU/mL
Methylation levels of CpG islands
Analysis of methylation levels was performed using the Infinium HumanMethylation450 BeadChip Kit (Illumina, San Diego, CA) according to the manufacturer's standard protocol, which interrogated 485,577 highly informative CpG sites at a single-nucleotide resolution for each sample. The experiment was performed with 0.5 μg of high-quality genomic DNA. On the chip, there were two bead types for each CpG site per locus. The raw data were analyzed by Genome Studio software (Illumina), and the fluorescence intensity ratios between the two bead types were calculated. A ratio value of 0 was equal to nonmethylation of the locus; a ratio of 1 was equal to total methylation. These raw data were corrected to normalize the differences in detection ranges between the two probes of the Infinium Assay using a peak-based correction method (15). Normalized data were filtered to exclude invalid probes such as null probes and probes with low reliability. R Statistical Software v3.1.2 (The R Foundation, Vienna, Austria) was used to perform the information processing.
Genotyping
Genomic SNP genotyping was performed using the Illumina® Infinium HumanOmni5-4 v1.1 BeadChips (Illumina). SNP editing was on call rate (>99%) and minor allele frequency (>1%). Subjects with >2.5% of missing genotypes were discarded. A total of 47,360 markers were retained after edits.
Statistical analysis
The SNPs categorical data association analysis was performed by using Pearson's chi-square test to detect differences in genotypic frequencies. Bonferroni correction was used to reduce the chances of obtaining false-positive results (type I errors). A p-value of <5.0E-8 was the threshold for significance (before Bonferroni correction). Statistical analysis was performed using Ruby v2.2.2p85 with statsample (
A paired t-test was used for within-pair analysis of methylation rate. Bonferroni correction was used to reduce the chances of obtaining false-positive results (type I errors). A p-value of <5.0E-7 was the threshold for significance (before Bonferroni correction). Statistical analysis was performed using R v3.1.2.
Results
Serum levels of TgAb
After setting a cutoff value for TgAb to >28 IU/mL (recommended cutoff value), 19 males (13.6% of all male) and 72 females (19.3% of all female) subjects were positive for TgAb. To exclude the subjects with marginally elevated levels and in order to perform stricter grouping, the cutoff value for positive TgAb was set to 50.0 IU/mL (TgAb negative: <28.0 IU/mL; TgAb positive: ≥50.0 IU/mL; TgAb borderline: ≥28.0 IU/mL and <50.0 IU/mL; Fig. 1). In order to avoid incorrect categorization of twin pairs due to small intra-individual variations of TgAb, subjects showing TGAb levels in the borderline zone were excluded. As shown in Figure 1, using these criteria, TgAb positive concordant (PC) pairs accounted for 5.7% (four pairs) and 9.6% (18 pairs) of male and female pairs, respectively. TgAb discordant (DC) pairs (one twin was positive and the co-twin was negative) accounted for 11.4% (eight pairs) and 8.0% (15 pairs) of male and female pairs, respectively. TgAb negative concordant (NC) pairs accounted for 78.6% (55 pairs) of male pairs and 74.3% (139 pairs) of female pairs. No significant differences were found in the frequencies of PC, NC, and DC pairs between male and female pairs. The male pairs were significantly older than female pairs in each of the PC, NC, and DC groups (Table 1). Female pairs in the PC group were older than those in the DC group (p < 0.05; Table 1). TgAb, thyrotropin (TSH), and free thyroxine (fT4) levels did not show sex-related differences in each of the PC, NC, and DC groups (Table 1). TSH and fT4 levels were not different between TgAb positive and negative twins in the DC group (Table 1). However, a few females in the NC group showed elevated TSH levels, probably due to reasons other than AITD.
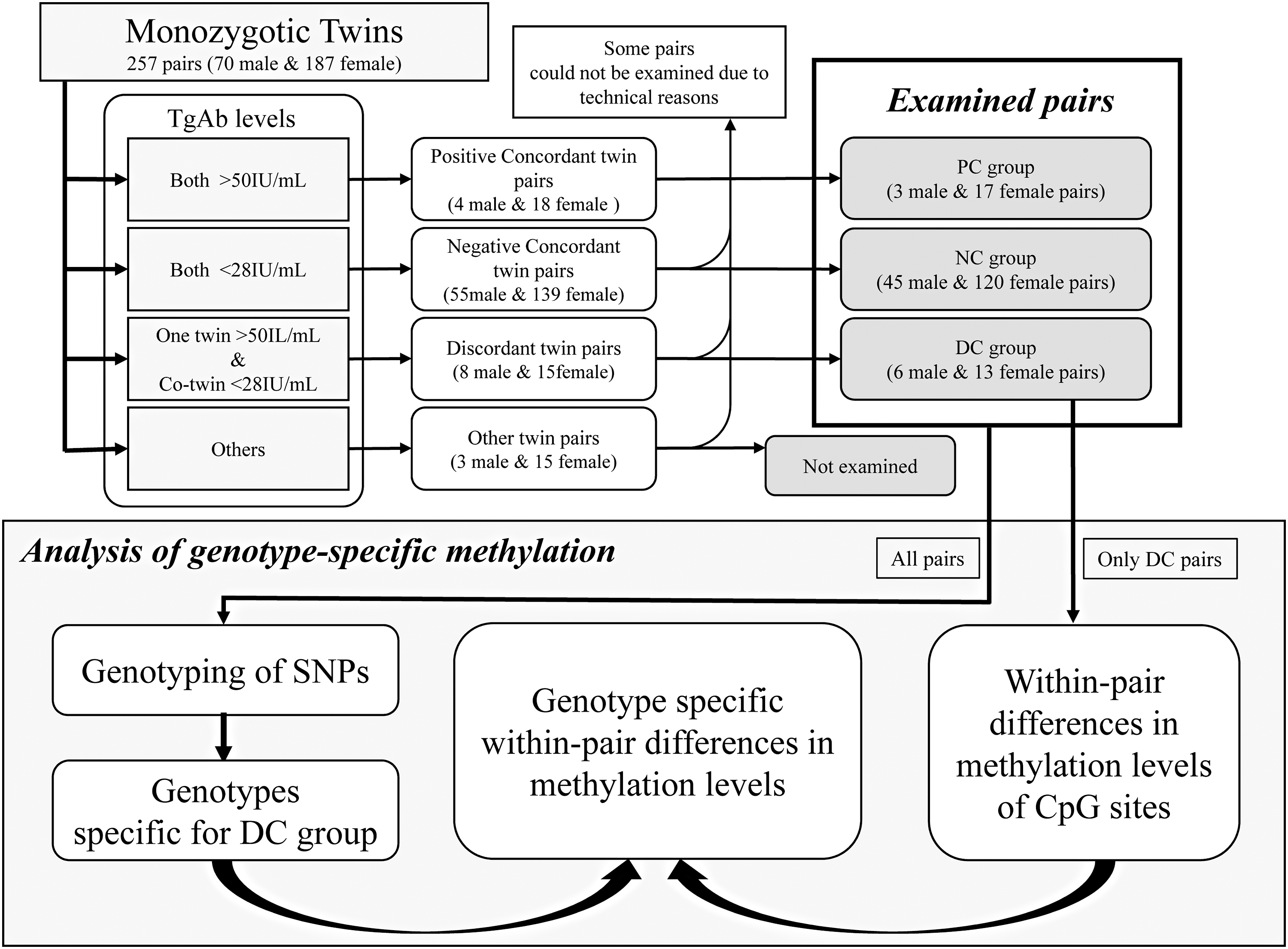
The criteria to categorize twin subjects and the protocol of analysis.
p < 0.05 vs. female of discordant group.
TgAb, antithyroglobulin autoantibodies; TSH, thyrotropin; fT4, free thyroxine; ND, Not detected.
Methylation levels of CpG sites in TgAb discordant pairs
As shown in Figure 2 and Supplementary Table S1 (Supplementary Data are available online at
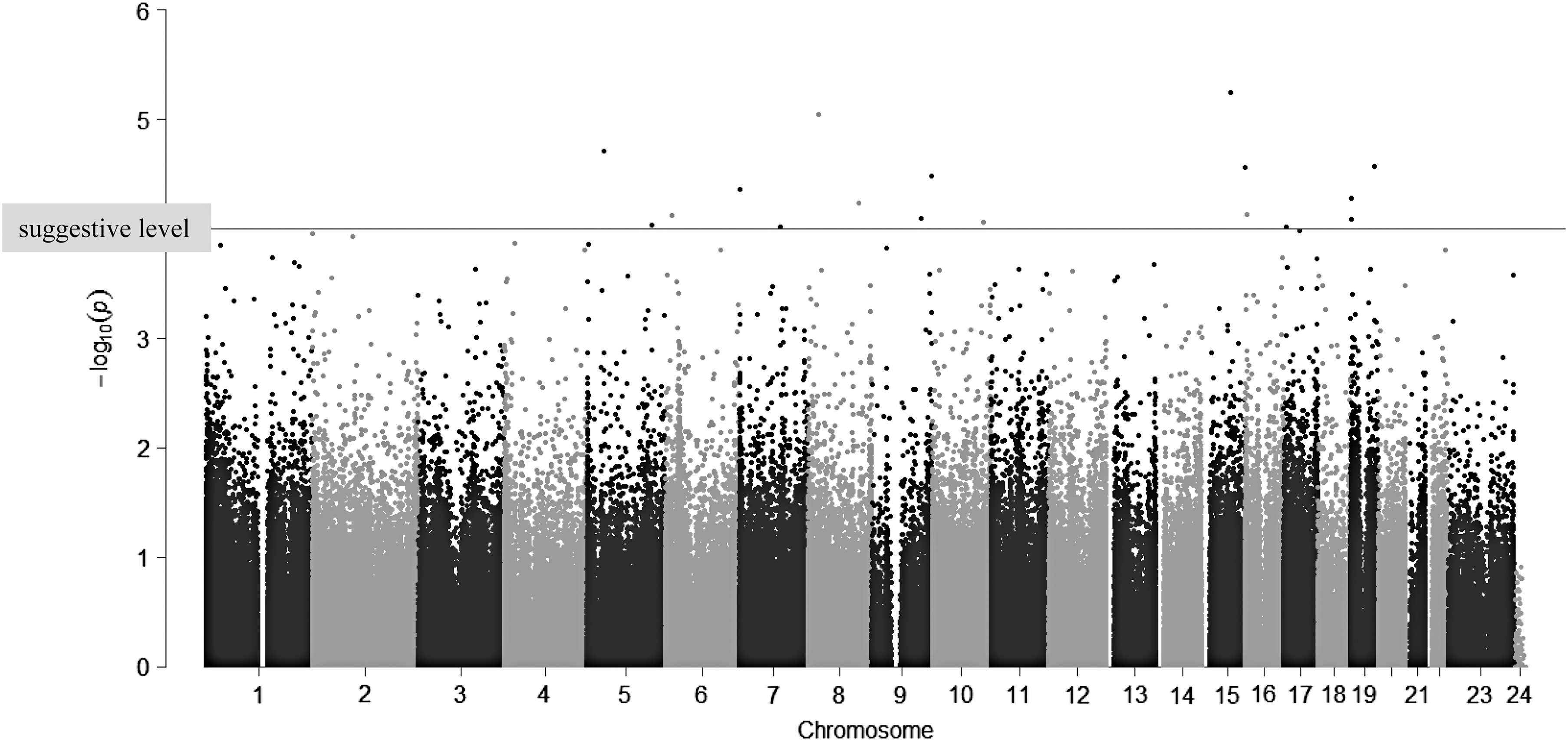
The differences in methylation levels between twins positive for antithyroglobulin autoantibodies (TgAb) and co-twins negative for TgAb in discordant twin pairs. A paired t-test was used for within-pair analysis of methylation rate. Bonferroni correction was used to reduce the risk of obtaining false-positive results (type I errors). A p-value of <5.0E-7 was the threshold for significance (before Bonferroni correction). The horizontal line indicates a suggestive but not significant level (1.0E-5) of the p-value after Bonferroni correction.
Differences in distributions of genotypes between TgAb concordant and discordant pair groups
As shown in Figure 3A, the distributions of some genotypes, mainly in chromosomes 6 and 14, were significantly different between PC and NC pair groups (significant markers are summarized in Supplementary Table S2). On the other hand, as shown in Figure 3B and C, the distributions of some genotypes in other chromosomes were significantly different between NC and DC pair groups and between concordant (CC; PC and NC) and DC pair groups. As shown in Table 2, 155 polymorphisms showed significant differences in the frequencies of genotypes between in TgAb DC and CC pair groups after correction for multiple comparisons.
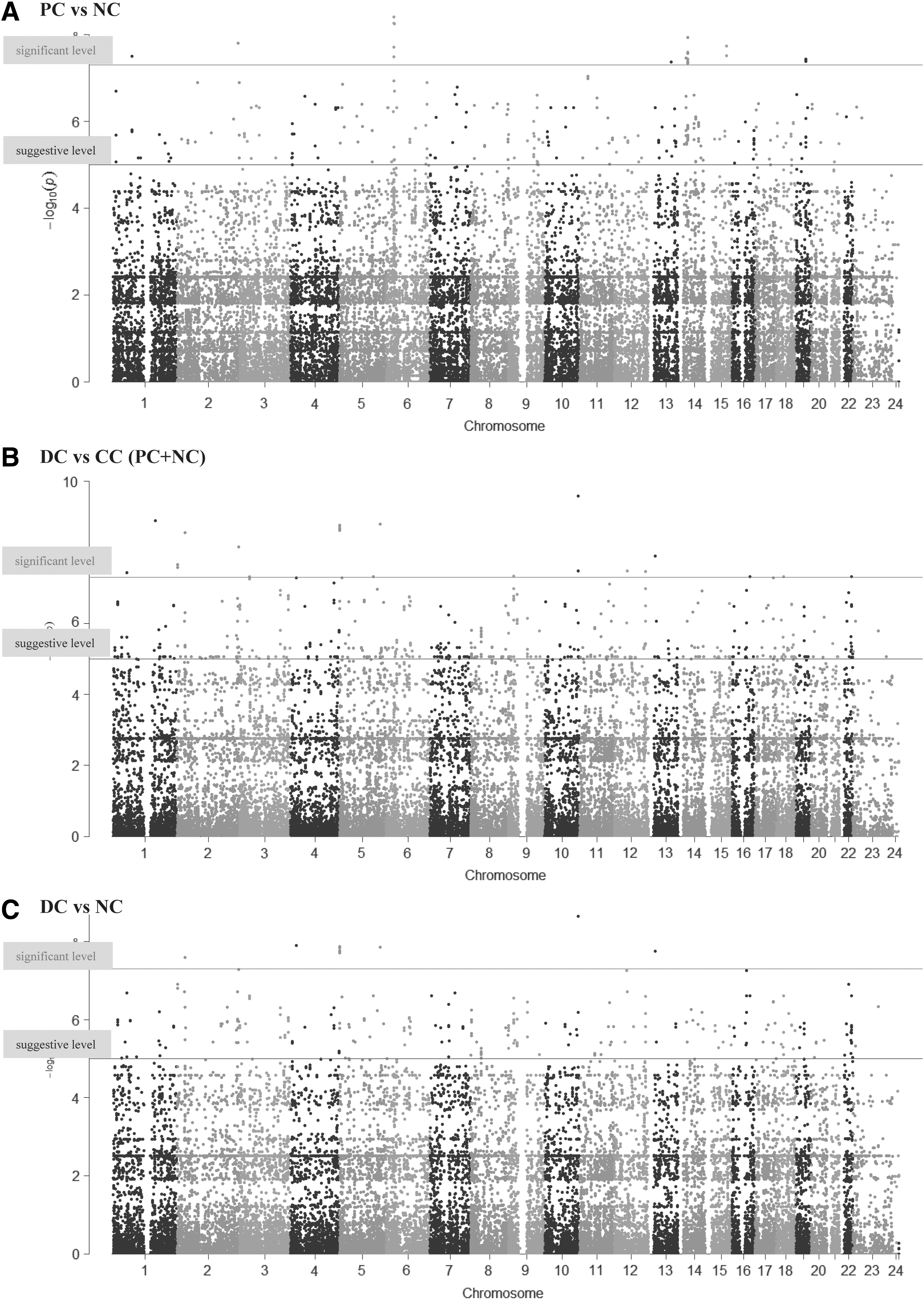
Manhattan plot of p-values in genome-wide association study between (
SNP names in parentheses are code used in Illumina kit. SNP used to analyze the genotype-specific methylation are indicated in bold.
p-Value tested by chi-square test; **corrected p-value by Bonferroni's correction.
Methylation levels of CpG sites in genotype-based TgAb discordant pairs
Among 155 polymorphisms showing significant differences in the genotype frequencies between TgAb DC and CC pairs (Table 2), the study focused on 10 polymorphisms with the following criteria: (i) polymorphisms must show the typical difference between DC and CC pairs, (ii) sample number of the specific genotype for the DC pair is enough to analyze, and (iii) one representative polymorphism among linked polymorphisms in each region must be selected.
In DC pairs with DC-specific genotypes of each of the representative polymorphism, within-pair differences in methylation levels of each CpG site were compared using a paired t-test (Supplementary Table S3). Among these representative polymorphisms, significant differences were found in DNA methylation levels of CpG sites between TgAb positive and negative twins of DC pairs with five specific polymorphisms (rs78946853T/C, rs56211683T/G, rs35707110C/T, rs31489C/A, and rs7290176G/A). In the DC pairs with rs35707110TT, there was a significant CpG site (cg10261589; p = 0). Because DNA methylation of this CpG site we could only be evaluated in two pairs, however, this site was excluded to increase reliability.
As shown in Figure 4 and Table 3, methylation levels of four CpG sites (cg17082856 in rs78946853CC, cg23898213 in rs56211683GG, cg00334293 in rs31489AA, and cg14990605 in rs7290176AA) were significantly different after correction for multiple comparisons between TgAb positive and negative co-twins within the DC-specific genotype. Figure 5 shows the parallel plots of methylation levels of these four CpG sites between the TgAb positive twin and the negative co-twin categorized into three DC-specific polymorphisms.
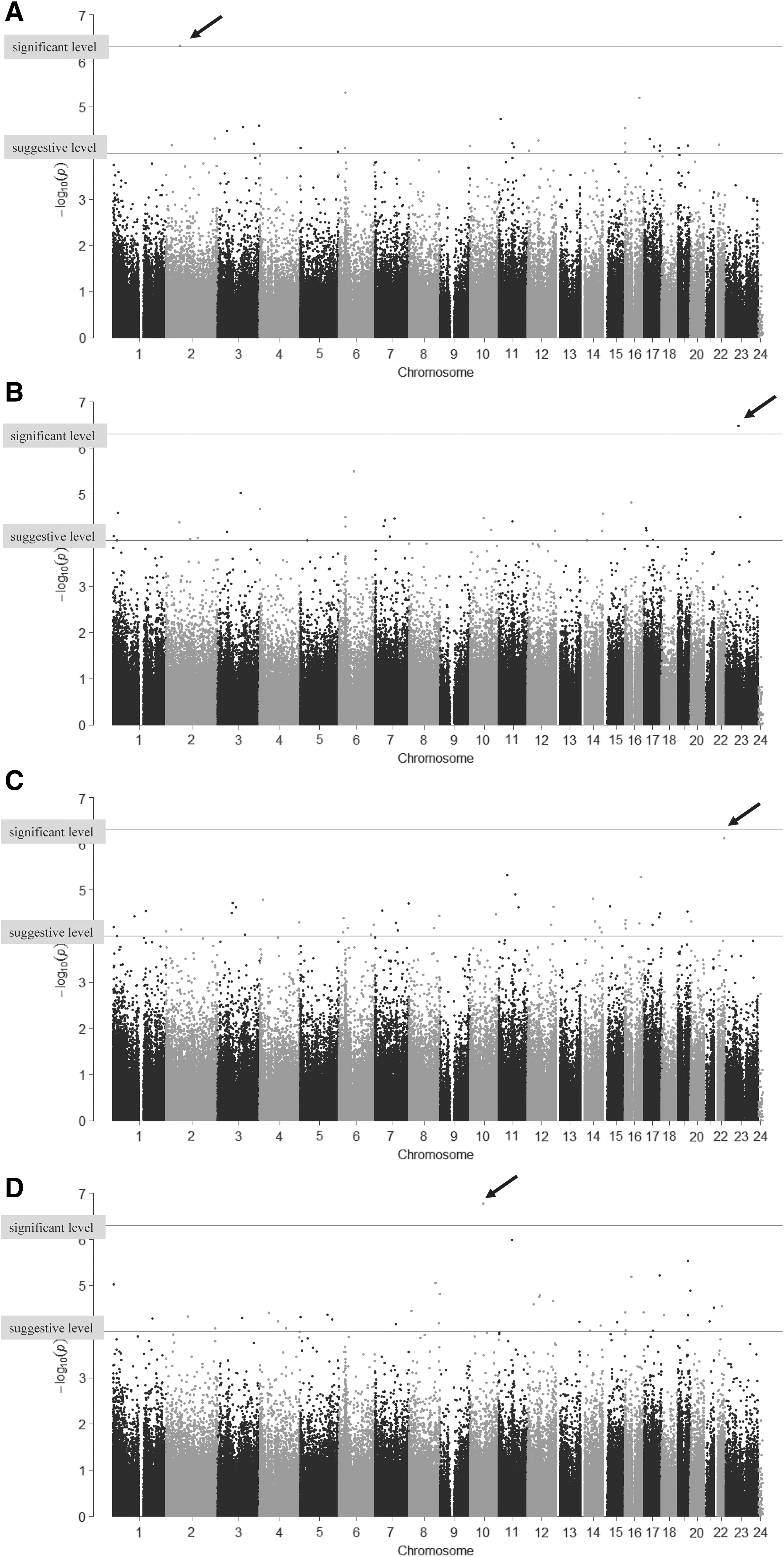
Manhattan plot of p-values in paired t-tests of differences in genotype-specific methylation levels between twins positive for TgAb and co-twins negative for TgAb in discordant twin pairs. (
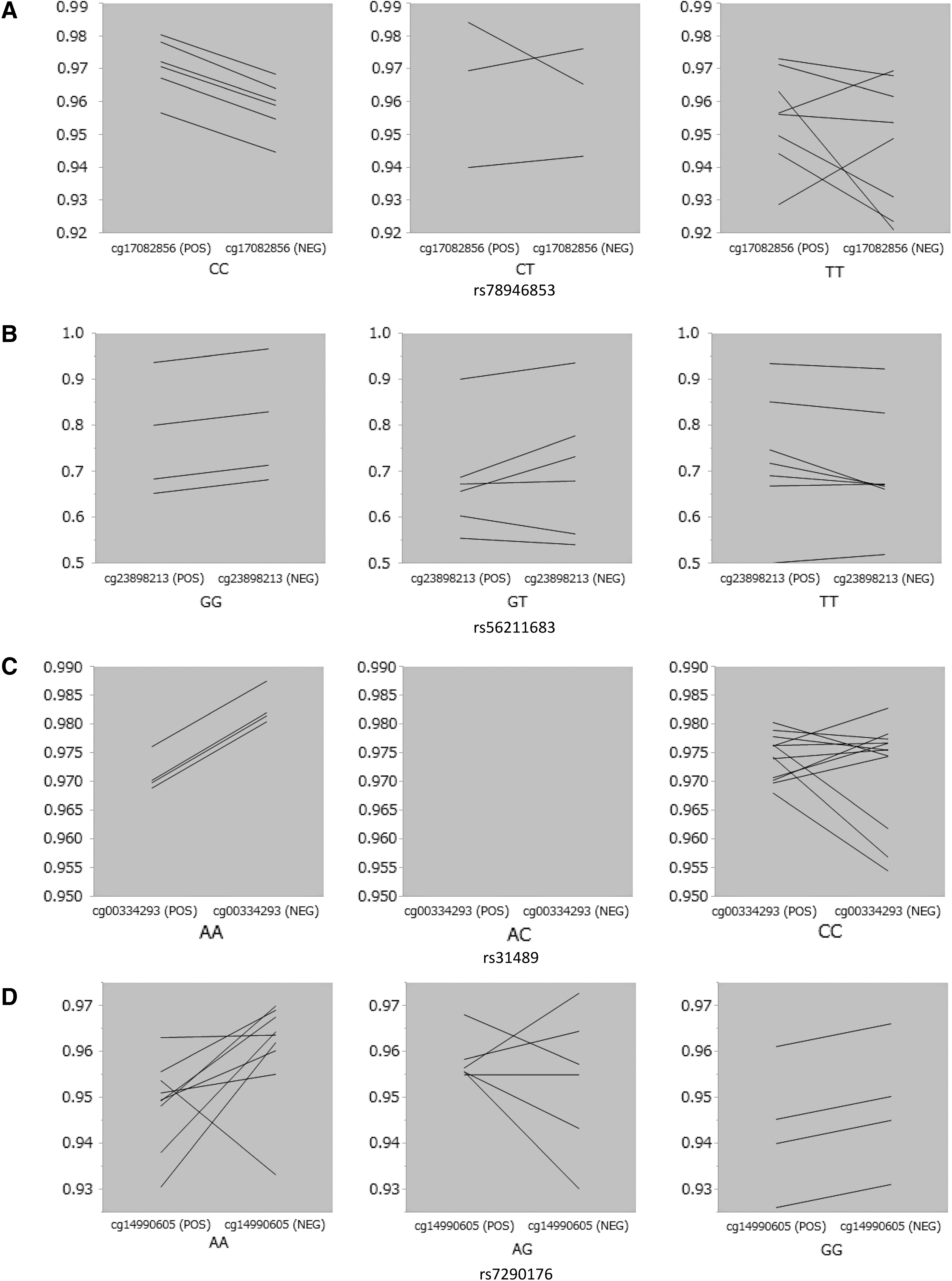
Parallel plots of genotype-specific methylation levels between twins positive for TgAb and co-twins negative for TgAb in discordant twin pairs. (
Significant p-values after correction for multi-comparison are shown in bold.
Discussion
Using recommended cutoff values for TgAb, the prevalence of TgAb in this study (13.6% of all male and 19.3% of all women) was slightly different from a previous report in Japanese (13.1% in male and 27.4% in female subjects) (16), probably because of differences in the method to measure the antibodies (ECLIA in this study vs. radioimmunoassay in the previous study).
In the present study, methylation levels of many CpG sites showed significant associations with the appearance of thyroid-specific autoantibodies (Fig. 2 and Supplementary Table S1). However, these associations were not statistically significant after multiple comparisons. It was supposed that the effects of DNA methylation would vary based on genetic background; that is, some DNA methylation in individuals with a specific genetic background will affect the development of autoantibodies, whereas those with another genetic background will not. Therefore, the study tried to determine significant genetic differences between TgAb concordant and discordant pairs to find specific genotypes for discordant pairs. As shown in Table 2 and Figure 3C, 155 polymorphisms were found showing significant differences in the genotype frequencies between TgAb DC and CC pairs. These polymorphisms were located in a large variety of genes (Table 2), and these results show specific genotypes for TgAb discordant pairs. These data suggest that some individuals may have a genetic susceptibility to epigenetic or environmental factors for autoantibody production, whereas others may be resistant to developing them.
Interestingly, chromosomes containing these significant polymorphisms were not always the same as those containing established AITD susceptibility genes (Fig. 3A, such as HLA located on chromosome 6) (1). This indicates that the genes related to genetic and epigenetic susceptibility to thyroid autoimmunity are different. Supporting this, as shown in Figure 3B, the chromosomes with significant polymorphisms between NC and DC were not always located on chromosomes containing AITD susceptibility genes. These data suggest that the balance among genetic susceptibility to TgAb production, genetic susceptibilities to epigenetic changes, and environmental factors affecting epigenetic changes may be independently related to the development of TgAb.
Next, the methylation levels of each CpG site were compared between TgAb positive and TgAb negative twins of each TgAb discordant pair with five specific genotypes for TgAb discordant pairs (rs78946853CC, rs56211683GG, rs35707110TT, rs31489AA, and rs7290176AA). As shown in Figure 4 and Table 3, four CpG sites showed significant differences between TgAb positive and negative twins in discordant pairs. As shown in Figure 5, methylation levels of cg17082856 were significantly higher in the TgAb positive twin than in the TgAb negative twin of discordant pairs with the rs78946853CC genotype. cg17082856 is located on the CEP68 gene and rs78946853 is located on the CAPN13 gene, and both genes are mapped on chromosome 2. CEP68 has been reported to be associated with acute urticaria/angioedema and a risk of aspirin-intolerant asthma (17,18), but no studies suggesting a relation of this gene with thyroid disorders or immune functions could be found. The CAPN13 gene, which is part of a family of cytosolic calcium-activated proteases involved in apoptosis, cell division, modulation of integrin-cytoskeletal interactions, and synaptic plasticity (19), is associated with hypertension (20). However, there is no report showing an association between CAPN13 and autoimmune disorders. In addition, no epistatic interactions between the CEP68 and CAPN13 genes resulting in an impact on autoimmunity have been reported.
Other significant CpG sites are cg23898213 in rs56211683GG, cg00334293 in rs31489AA, and cg14990605 in rs7290176AA. The genes containing each of these CpG sites and the genes containing each specific polymorphism are not mapped to the same chromosomes, as shown in Table 3. These data suggest that epigenetic changes may not always be affected by genetic factors on the same chromosome. Some of the genes associating with these polymorphisms are CLPTM1 L and LARGE1. CLIPTM1 L encodes a membrane protein, and polymorphisms in this gene have been reported to increase susceptibility to several cancers, including lung and pancreatic cancers (21,22). LARGE1 encodes a member of the N-acetylglucosaminyltransferase gene family, and mutations of this gene cause congenital muscular dystrophy with severe mental retardation (23). However, no reports of these genes showing associations with autoantibody production could be found. Moreover, it is not possible to comment on the effect of epigenetic changes in this gene because the region containing these specific CpG sites was not close to any novel genes, which is a limitation of this study. In addition, more samples are desirable to corroborate the findings of the analysis.
In conclusion, this twin study provides some insights into the genetic background underlying the epigenetic susceptibility for developing thyroid autoimmunity. Several coding or non-coding DNA regions may be associated with the production of thyroid-specific autoantibodies through epigenetic changes based on specific genetic background. However, the effects of these epigenetic changes on gene expression must be confirmed.
Footnotes
Acknowledgments
The authors are thankful to all the consultants from the Osaka Twin Research Group (Yoshinori Iwatani [lead author:
Author Disclosure Statement
The authors report no conflicts of interest in this work.