
Abstract
Background:
Anaplastic thyroid carcinoma (ATC), the most aggressive form of thyroid cancer, is unresponsive to radioiodine therapy. The current study aimed to extend the diagnostic and therapeutic application of radioiodine beyond the treatment of differentiated thyroid cancer by targeting the functional sodium–iodide symporter (NIS) to ATC.
Methods:
The study employed nanoparticle vectors (polyplexes) based on linear polyethylenimine (LPEI), shielded by polyethylene glycol (PEG) and coupled to the synthetic peptide GE11 as an epidermal growth factor receptor (EGFR)-specific ligand in order to target a NIS-expressing plasmid (LPEI-PEG-GE11/NIS) to EGFR overexpressing human thyroid carcinoma cell lines. Using ATC xenograft mouse models, transfection efficiency by 123I scintigraphy and potential for systemic radioiodine therapy after systemic polyplex application were evaluated.
Results:
In vitro iodide uptake studies in SW1736 and Hth74 ATC cells, and, for comparison, in more differentiated follicular (FTC-133) and papillary (BCPAP) thyroid carcinoma cells demonstrated high transfection efficiency and EGFR-specificity of LPEI-PEG-GE11/NIS that correlated well with EGFR expression levels. After systemic polyplex injection, in vivo 123I gamma camera imaging revealed significant tumor-specific accumulation of radioiodine in an SW1736 and an Hth74 xenograft mouse model. Radioiodine accumulation was found to be higher in SW1736 tumors, reflecting in vitro results, EGFR expression levels, and results from ex vivo analysis of NIS staining. Administration of 131I in LPEI-PEG-GE11/NIS-treated SW1736 xenograft mice resulted in significantly reduced tumor growth associated with prolonged survival compared to control animals.
Conclusions:
The data open the exciting prospect of NIS-mediated radionuclide imaging and therapy of ATC after non-viral reintroduction of the NIS gene. The high tumor specificity after systemic application makes the strategy an attractive alternative for the treatment of highly metastatic ATC.
Introduction
T
Cloning of NIS in 1996 was the first step in the development of a novel cytoreductive gene therapy strategy based on targeting functional NIS to both thyroidal and extrathyroidal malignancies, thus extending an established and highly effective anticancer strategy beyond the treatment of differentiated thyroid cancer (8 –10). It has been shown that selective NIS gene transfer into tumor cells allows systemic application of radionuclides, resulting in tumor-specific radionuclide concentration that can be used both for non-invasive imaging as well as therapy (11 –24). Clinical development of the NIS gene therapy concept is dependent on highly specific and efficient tumor targeting at low toxicity and requires systemic application of the therapeutic gene, especially for the treatment of disseminated cancer. However, the biostability and delivery efficiency of “naked” nucleic acids are very low, limiting their systemic applicability (25). Cationic polymers, such as linear polyethylenimine (LPEI), are a class of non-viral gene delivery vehicles that form submicrometer complexes with nucleic acids called polyplexes and thus stabilize the DNA by eliminating unwanted interactions with blood components and other non-target sites (25). After intravenous application, nanoparticle concentration builds up in the plasma with a long half-life, as nanoparticles escape both renal clearance and penetration of normal endothelium due to their size (26). LPEI-based polyplexes are internalized with high efficiency and exhibit intrinsic endosomolytic activity, allowing efficient release into the cytoplasm followed by nuclear import of the therapeutic gene. Tumor targeting is based on the so-called enhanced permeability and retention (EPR) effect that drives passive accumulation of polyplexes due to the leaky vasculature prevalent in tumors in conjunction with a tumor's slow venous return and poor lymphatic clearance (25 –27). Due to inter- and intratumoral heterogeneity, nanoparticle-based drugs that rely on the EPR effect alone for efficient delivery have their limitations in the clinical setting, and efficiency will vary from patient to patient (28). However, tumor specificity and delivery efficacy can be further increased using ligands that bind to receptors overexpressed on tumor cells. To this end, the epidermal growth factor receptor- (EGFR-) specific ligand GE11 was attached to LPEI via a polyethylene glycol (PEG) link that facilitates surface shielding of polyplexes and thus reduces cytotoxicity (29). GE11 polyplexes undergo efficient clathrin-mediated endocytosis and are quickly transported to the nucleus (30). In previous work, LPEI-PEG-GE11 polymers were shown to be highly efficient NIS gene delivery vehicles in a high EGFR expressing liver cancer xenograft mouse model, as well as an advanced genetically engineered mouse model of pancreatic ductal adenocarcinoma, as evidenced by tumor-specific radioiodine accumulation and therapeutic efficacy of 131I (13,24).
As EGFR is typically overexpressed on ATC cells (31), the LPEI-PEG-GE11 nanoparticle vectors were used for NIS gene transfer to ATC in the present study in order to capitalize on the dual diagnostic and therapeutic function of NIS.
Materials and Methods
EGFR and NIS immunohistochemistry of human anaplastic tumor tissue
In total, nine different tumors stored in the database of the Institute of Pathology from ATC patients, who had undergone surgery between 2003 and 2012, were examined. Immunohistochemical staining of EGFR was performed on a BenchMark XT slide staining system (Ventana Medical Systems, Tucson, AZ) using a monoclonal (IgG1) mouse antibody (clone: 3C6; Ventana Medical Systems). Evaluation of the staining pattern was performed by visual scoring. No staining resulted in a score of 0; little non-circumferential staining in >10% of the tumor cells was scored as 1+; weak but thin circumferential staining in >10% of the tumor cells was rated with a score of 2+; and a score of 3+ signified an intense, thick circumferential staining in >10% of the tumor cells. Scores of 0 were rated as negative, while scores of 1+, 2+, and 3+ indicated a weak, moderate, and strong positive expression of EGFR, respectively (32). NIS staining was performed as described previously using a mouse monoclonal antibody (Millipore, Darmstadt, Germany) diluted 1:500 (33).
Cell lines
ATC cell lines SW1736 and Hth74, follicular thyroid carcinoma cell line FTC-133, and papillary thyroid carcinoma cell line BCPAP were grown in RPMI (Thermo Fisher Scientific, Waltham, MA) supplemented with 10% (v/v) fetal bovine serum (FBS; PAA, Colbe, Germany) and 100 IU/mL of penicillin/100 μg/mL streptomycin (Thermo Fisher Scientific). Cells were maintained at 37°C and 5% (v/v) CO2 in an incubator with 95% humidity. Cell culture medium was replaced every second day, and cells were passaged at 70% confluence.
Flow cytometry
For analysis of cell surface EGFR expression by flow cytometry, cells were trypsinized, re-suspended in FACS buffer (phosphate-buffered saline supplemented with 10% [v/v] FBS), and diluted to a density of 6 × 105 cells/100 μL. Cells were incubated with a mouse monoclonal anti-human EGFR antibody (Dako, Hamburg, Germany) or control antibody (mouse IgG; Dako) for 1.5 h at 4°C. Cells were washed with FACS buffer and incubated with an Alexa488 conjugated polyclonal goat anti-mouse antibody (1:400; Dianova, Hamburg, Germany) for 1 h at 4°C. To discriminate live and dead cells, propidium iodide (Sigma–Aldrich, Taufkirchen, Germany) was added before acquisition. Analysis was performed on a BD FACSCanto II flow cytometer (BD, Heidelberg, Germany). Cell aggregates and debris were excluded from analysis by appropriate gating.
Polyplex formation
LPEI-based polymers were condensed with plasmid DNA at specific conjugate to plasmid ratios (c/p) in HEPES-buffered glucose, as described previously (34), and incubated on ice for 20 min before use. Final DNA concentrations were 2 μg/mL for in vitro studies and 200 μg/mL for in vivo studies.
Transient transfection
For in vitro transfection experiments, cells were grown to 70–80% confluency, and washed and incubated with LPEI-PEG-GE11/NIS polyplexes at c/p 0.8 in the absence of serum and antibiotics for 4 h. As controls, cells were transfected with LPEI-PEG-Cys/NIS, a polyplex without the active EGFR targeting ligand or non-coding LPEI-PEG-GE11/antisenseNIS (NIS sequence back to front). LPEI-PEG-GE11/NIS-transfected cells treated with the NIS-specific inhibitor perchlorate (1 mM of potassium perchlorate; Merck, Darmstadt, Germany) served as an additional control. After further incubation in full growth medium for 24 h, transfection efficiency was measured by iodide uptake activity.
125I uptake assay
Iodide uptake was analyzed, as described previously (35,36).
Establishment of SW1736 and Hth74 xenografts
Female CD1 nu/nu mice (Charles River, Sulzfeld, Germany) were maintained under pathogen-free conditions with free access to standard nude mouse diet containing 2.2 mg/kg iodine (ssniff, Soest, Germany) and water ad libitum. Tumors were established by subcutaneous injection of 5 × 106 tumor cells in 100 μL of phosphate-buffered saline into the flank region. Tumors were measured twice a week, and tumor volumes were calculated using the equation: tumor volume =length × width × height × 0.52. To reduce thyroidal iodide uptake and thus enhance tumoral uptake, animals were orally pretreated with 5 mg/L of L-thyroxine (Sigma–Aldrich) for 10 days prior to radioiodine (123I, 131I) application, as described previously (14,15,18). The experimental protocol was approved by the regional governmental commission for animals (Regierung von Oberbayern, Munich, Germany).
Radioiodine uptake in vivo
Once subcutaneous tumors reached a diameter of 8–10 mm (approximately 10–12 weeks after tumor cell injection), two rounds of LPEI-PEG-GE11/NIS (n = 3 per tumor model) were injected intravenously (i.v.) via the tail vein 48 h and 24 h prior to intraperitoneal (i.p.) injection of 18.5 MBq (0.5 mCi) 123I (GE Healthcare, Braunschweig, Germany). As a control, a subset of mice (n = 1 per tumor model) received 2 mg/mouse of the competitive NIS inhibitor sodium perchlorate (Sigma–Aldrich) 30 min before 123I injection. Radioiodine biodistribution was assessed using a gamma camera equipped with a UXHR collimator
Indirect immunofluorescence assay
Immunofluorescence staining of frozen tissue sections from LPEI-PEG-GE11/NIS-treated mice was performed, as described previously (36), using a rabbit polyclonal NIS-specific primary antibody (Acris Antibodies, Herford, Germany; 1:1000) and a Alexa488-conjugated secondary antibody (Jackson ImmunoResearch, Suffolk, United Kingdom; 1:400). Sections were imaged at 40 × magnification on an Axiovert 135 TV fluorescence microscope equipped with an AxioCam MRm CCD camera and AxioVision Rel. 4.8 software (Carl Zeiss, Munich, Germany).
In vivo 131I therapy study
Therapy trials were initiated in SW1736 xenografts eight weeks after tumor cell injection at tumor diameters of approximately 5–10 mm. A cycle consisting of polyplex administration on two consecutive days and a single radioiodine application 24 h later was repeated a total of three times: polyplexes LPEI-PEG-GE11/NIS or, as controls, LPEI-PEG-GE11/antisenseNIS or NaCl were administered systemically via the tail vein on days 0/1, 3/4, and 7/8 followed by i.p. application of 55.5 MBq (1.5 mCi) 131I (GE Healthcare) or, as control, saline on days 2, 5, and 9 (LPEI-PEG-GE11/NIS +131I, n = 10; LPEI-PEG-GE11/antisenseNIS + 131I, n = 10; LPEI-PEG-GE11/NIS + NaCl, n = 15; NaCl + NaCl, n = 11). Mice were sacrificed when at least one endpoint criterion was reached, that is, when a tumor volume >1500 mm3 was reached or when tumors started to exulcerate.
Statistics
All in vitro experiments were performed in triplicate. Results are expressed as mean ± standard error of the mean (SEM), mean fold change ± SEM, or, for visual EGFR scoring of human ATC samples and survival plots, percent. Statistical significance was tested by two-tailed Student's t-test or, for tumor volumes, one-way analysis of variance followed by Tukey's Honestly Significant Difference test. Statistical significance of Kaplan–Meier plots was analyzed by log-rank test. p-Values of <0.05 were considered statistically significant (*p < 0.05; **p < 0.01; ***p < 0.001).
Results
EGFR and NIS immunoreactivity of human ATC tissue
In total, nine anaplastic thyroid tumors from different patients were examined and ranked according to their EGFR expression. Seven (78%) of the nine examined tumors were categorized as 3+ (Fig. 1A and C), while one (11%) tumor was classified as 2+ and one (11%) showed no EGFR-specific immunoreactivity (Fig. 1B and C). Immunohistochemical staining for NIS revealed no differences in NIS staining between EGFR-positive (score 3+) and EGFR-negative (score 0) ATC tissue (Fig. 1D and E). Graves' tissue served as a positive control for NIS staining (Fig. 1F).

Epidermal growth factor receptor (EGFR)-targeted NIS gene transfer in vitro. Human anaplastic thyroid carcinoma (ATC) samples (n = 9) were ranked according to their EGFR expression. Highly EGFR-positive sections were classified as 3+ (
EGFR-targeted NIS gene transfer in vitro
Two different human ATC cell lines and, for comparison, more differentiated follicular and papillary thyroid carcinoma cell lines were examined for their EGFR expression levels by flow cytometry. SW1736 cells showed high EGFR expression, while intermediate EGFR levels were measured on Hth74, FTC-133, and BCPAP cells (Fig. 1G).
Twenty-four hours after transient transfection with LPEI-PEG-GE11/NIS, NIS-specific iodide uptake was detected in all four cell lines. SW1736 cells showed a 25-fold increase, Hth74 and BCPAP cells a 10-fold increase, and FTC-133 cells a sevenfold increase in 125I uptake compared to background levels measured in untransfected cells. The NIS-specific inhibitor perchlorate completely blocked iodide uptake in all cell lines. After transfection with unspecific LPEI-PEG-Cys/NIS, iodide uptake was significantly reduced compared to EGFR-targeted polyplexes LPEI-PEG-GE11/NIS. Cells transfected with LPEI-PEG-GE11/antisenseNIS showed no gain in uptake compared to untransfected cells (Fig. 1H). The amount of iodide uptake correlated well with EGFR levels measured by flow cytometry. Comparing the two ATC cell lines to each other, SW1736 cells showed a significantly higher iodide uptake than Hth74 cells (Fig. 1H).
EGFR-targeted NIS gene transfer in vivo
Twenty-four hours after two rounds of systemic NIS gene transfer via i.v. application of LPEI-PEG-GE11/NIS, 123I uptake was evaluated on a gamma camera. Serial scanning revealed maximum tumoral iodide accumulation 1 h post 123I injection based on NIS-mediated iodide influx from the blood circulation. Over time, iodide content in the circulation decreased due to renal clearance. In the tumor, the radioiodine recirculated to a certain extent, as iodide lost by one tumor cell was taken up by neighboring NIS-transduced cells. Due to lack of iodide organification, however, tumoral iodide content slowly decreased over time, as 123I was completely cleared from the circulation (Fig. 2A–C). A maximum 123I uptake of 5.6–7.8% ID/g was observed for SW1736 tumors, whereas Hth74 tumors accumulated 4.5–5.8% ID/g (Fig. 2A–C). For 131I, this translates to a tumor-absorbed dose of 35.1 mGy/MBq/g tumor, with an effective half-life of 3.1 h in the SW1736 model and 25.0 mGy/MBq/g tumor with an effective half-life of 2.7 h in the Hth74 model. Pretreatment of LPEI-PEG-GE11/NIS transfected xenografts with perchlorate inhibited tumoral accumulation of radioiodine in both models (Fig. 2A and B). Physiological iodide uptake in the thyroid, salivary glands, and stomach was also blocked upon perchlorate treatment. Radioiodine accumulation in the bladder is due to renal elimination of radioiodine (Fig. 2A and B).
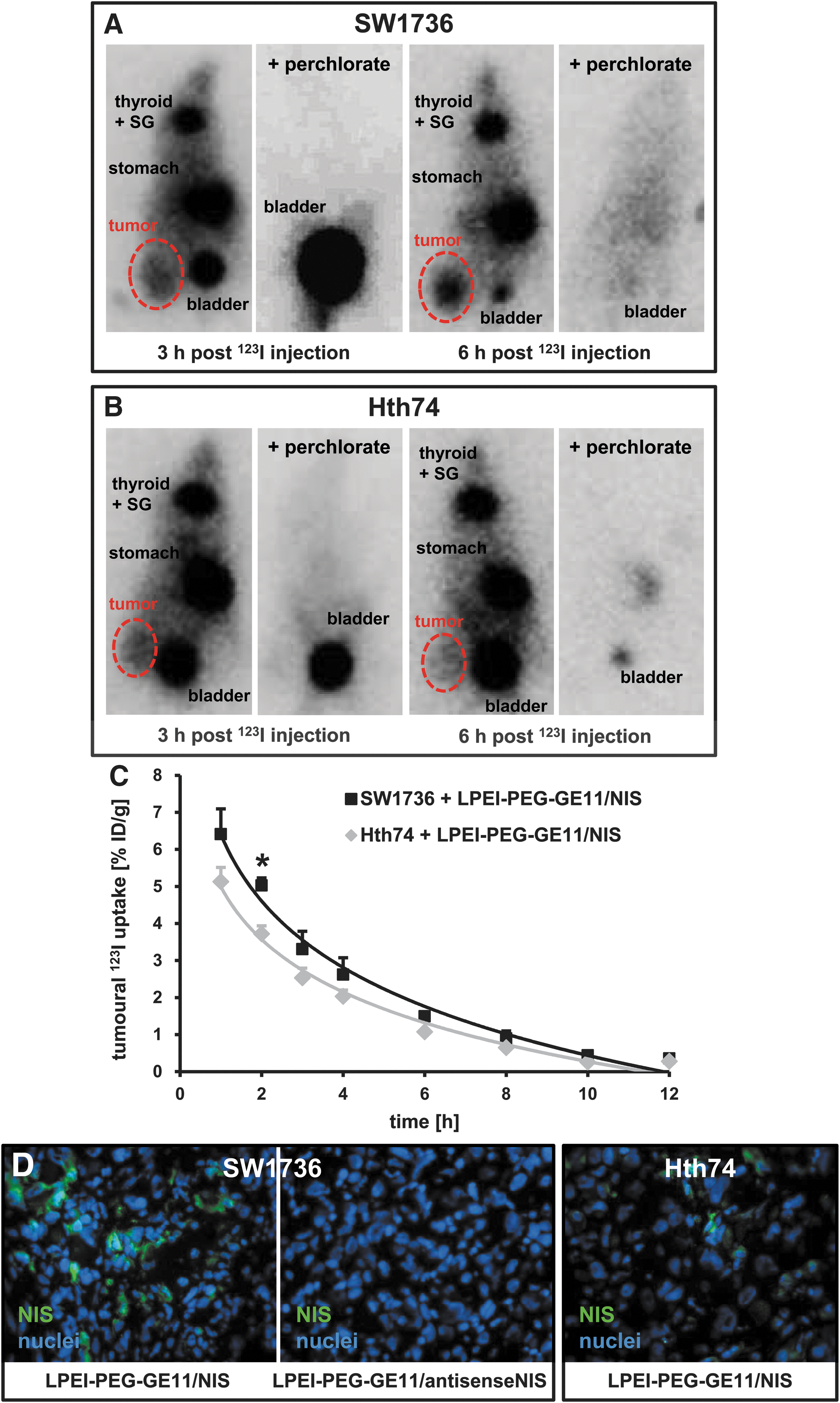
In vivo imaging of NIS-mediated iodide uptake. 123I scintigraphy revealed tumor-specific radioiodide uptake in SW1736 (
After gamma camera imaging, mice were sacrificed and dissected. In both SW1736 and Hth74 tumors from LPEI-PEG-GE11-treated mice, immunofluorescence staining revealed areas of NIS-specific staining, and the amount of NIS-positive cells in SW1736-derived tumors seemed higher than in Hth74 tumors (Fig. 2D). SW1736 tumors transfected with LPEI-PEG-GE11/antisenseNIS, in contrast, showed no NIS-specific immunoreactivity (Fig. 2D).
131I therapy after in vivo NIS gene transfer
Mice harboring SW1736 xenograft tumors were treated with three cycles of LPEI-PEG-GE11/NIS followed by 131I 24 h later (Fig. 3A). Mice injected with LPEI-PEG-GE11/NIS and saline, LPEI-PEG-GE11/antisense-NIS and 131I, or saline only served as controls and exhibited a continuous, exponential tumor growth (Fig. 3B). Mice injected with LPEI-PEG-GE11/NIS and 131I, in contrast, showed a significant delay in tumor growth, which resulted in prolonged survival (Fig. 3B and C). Thirty-five days after therapy started, the last control mouse was sacrificed, while 60% of mice injected with LPEI-PEG-GE11/NIS and 131I were still alive (Fig. 3C). Median survival times were 42 days for the therapy group, 28 days for both LPEI-PEG-GE11/antisenseNIS + 131I- and NaCl + NaCl-treated animals, and 18 days for the LPEI-PEG-GE11/NIS + NaCl group.
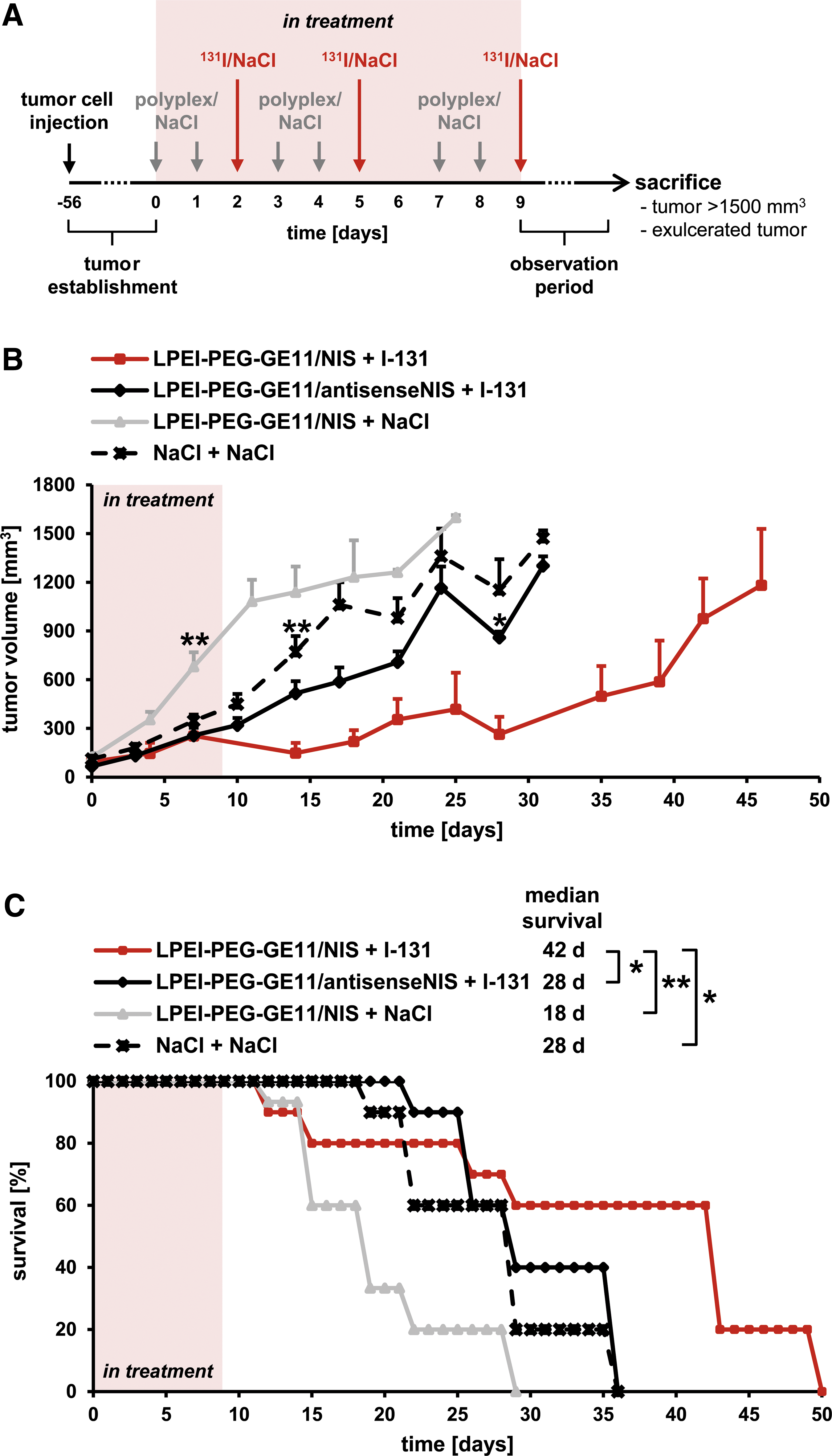
Therapeutic application of 131I after NIS gene transfer in vivo. Mice harboring SW1736 xenograft tumors were treated with three cycles of dual intravenous injections of polyplexes followed by intraperitoneal injection of 55.5 MBq 131I (
Discussion
Besides its key role in thyroid physiology, NIS-mediated iodide uptake provides the molecular basis for diagnostic imaging and targeted radionuclide therapy for the treatment of differentiated thyroid carcinomas. This concept, however, has proven ineffective in poorly differentiated and undifferentiated forms of thyroid cancer that lose the ability to accumulate iodide due to reduced or absent expression of functional NIS (37). A very elegant and obvious therapy approach is to reinstate NIS expression and thus make it susceptible to radioiodine therapy either by re-differentiation aimed at recovery of endogenous NIS expression or by transfection with exogenous NIS.
Substances for the induction of re-differentiation in dedifferentiated thyroid cancer that have been tested to date include retinoids, thiazolidinediones, such as rosiglitazone, and regulators of epigenetic modifications, that is, histone deacetylase (HDAC) inhibitors and DNA methyltransferase inhibitors (38 –43). Clinical success rates, however, have remained unexpectedly low for all of these compounds (42,44). More recently, the MAPK kinase inhibitor selumetinib and the selective BRAF inhibitor dabrafenib were shown to induce clinically relevant levels of radioiodine uptake in patients with radioiodine-refractory differentiated thyroid cancer (45,46).
The alternative strategy of exogenously introducing NIS was made available by the cloning of the NIS gene in 1996 (8,9). Early work in tumorigenic Fisher rat thyroid follicular cells transfected stably with rat NIS demonstrated iodide uptake capacity in vitro and in vivo (47). Similarly, after transfection of human ATC cells with NIS and subsequent injection into nude mice, tumors could be imaged by scintigraphy and were susceptible to radionuclide therapy (48 –50). Intratumoral injection of NIS using adenoviral vectors in a medullary thyroid carcinoma xenograft mouse model and measles virus vectors in an ATC xenograft mouse model restored radioiodine uptake, as shown by gamma camera or SPECT imaging, respectively, and therapeutic efficacy of 131I (33,51). For the development of this concept toward clinical application, especially in metastatic disease, however, systemic application of NIS followed by safe, efficient, and tumor-specific delivery are essential.
Previous work has focused on systemic delivery of the NIS gene to extrathyroidal tumors using viral vectors, mesenchymal stem cells, and synthetic polymers as gene transfer vehicles (12 –24). Based on the high efficiency of EGFR-targeted LPEI-PEG-GE11 polymers for tumor-specific delivery of functional NIS to a liver cancer xenograft mouse model, as well as an advanced genetically engineered mouse model of pancreatic ductal adenocarcinoma in earlier studies and the high expression levels of EGFR in human ATC tissue samples, these synthetic polymers were chosen as delivery vehicles for the NIS gene to ATC. High EGFR expression on ATC was confirmed by FACS analysis of human ATC cell lines SW1736 and Hth74. Both cell lines express wild-type EGFR protein, though Hth74 harbor a heterozygous silent polymorphism in the EGFR gene (52,53). Transient in vitro transfection of these cells with LPEI-PEG-GE11/NIS led to significant perchlorate sensitive and therefore NIS-mediated radioiodine uptake. In comparison, iodide uptake was significantly lower after transfection with non-targeted LPEI-PEG-Cys/NIS, demonstrating improved transfection efficiency using the peptide ligand GE11. NIS dependency was further confirmed by the absence of iodide uptake activity using non-coding antisenseNIS DNA. The higher EGFR levels detected on SW1736 cells by FACS were reflected in higher iodide uptake activity compared to Hth74 cells. Similar results were obtained in more differentiated follicular and papillary thyroid carcinoma cell lines, demonstrating the feasibility of this strategy for NIS gene transfer in EGFR-positive radioiodine refractory thyroid carcinoma in general.
Transferring these results to systemic vector application in vivo, i.v. administration of LPEI-PEG-GE11/NIS resulted in significant perchlorate sensitive, tumor-specific iodide uptake in xenograft mouse models derived from both ATC cell lines, although again SW1736 showed higher 123I uptake, as determined by gamma camera imaging and confirmed by immunofluorescence staining. The biological half-life measured in ATC tumors was similar to what we have observed in previous studies in other tumor models, which is clearly long enough for a significant therapy effect (21 –24). The radiation dose that reaches the tumor depends on the rate of iodide uptake, its retention time in the tumor cell, that is, iodide efflux, iodide recirculation within the tumor, and iodide organification (54). Due to loss of thyroid peroxidase (TPO) and thyroglobulin expression in ATC as a result of dedifferentiation, no iodide organification was seen in any of the ATC xenografts. However, the iodide retention time is longer than one would expect without organification, as the iodide is recirculated within the tumor. A possible approach to enhance iodide retention time and the delivered dose in non-organifying tissues is co-transfection of NIS and TPO (54). In this context, co-expression of both genes decreased iodide efflux in non-small-cell lung cancer cell lines and enhanced tumor cell apoptosis compared to single transfection with NIS (55). In contrast, after co-infection of human cervix carcinoma cells with two adenoviral vectors encoding NIS and TPO, no increase in iodide retention time was detected, though active TPO was produced, and a significant increase in organification was observed (56). To date, there is no clear evidence that TPO-mediated organification does indeed enhance the efficacy of NIS-based gene therapy. No clear data exist to prove that organification is a crucial prerequisite for successful 131I therapy. On the contrary, the results in the current study, as well as in earlier studies in various non-organifying tumor entities, demonstrate that iodide organification is not required for accumulation of therapeutically effective radioiodine doses.
The differences in both in vitro and in vivo data between the two ATC cell lines used here are presumably due to differences in EGFR expression levels. Tumor heterogeneity is a typical problem seen in clinical practice. Indeed, of the nine patient samples analyzed for EGFR receptor expression in this study, one tumor showed slightly lower EGFR receptor expression, while one showed no expression at all. For this reason, the arsenal of targeting strategies is constantly being broadened, so that different targeting ligands can be applied depending on receptor status. In this context, a further interesting candidate for ATC targeting is transferrin, as the transferrin receptor is highly expressed on ATC (57 –59).
Due to the more promising results in SW1736, the subsequent evaluation of the therapeutic effectiveness of 131I after LPEI-PEG-GE11-mediated systemic NIS gene delivery was performed in this model only. After three cycles of two applications of LPEI-PEG-GE11/NIS followed by an application of 131I in SW1736 xenograft tumor carrying mice, it was possible to demonstrate a significant delay in tumor growth in therapy animals compared to control groups, which was accompanied by a significant increase in survival time.
Taken together, the data clearly show the great potential of EGFR-targeted synthetic polymers to target NIS to ATC. Systemic application is of particular importance in ATC, as it rapidly metastasizes to distant sites and, at the time of diagnosis, metastases, predominantly in the lung, liver, bone, and brain, are already present in 50% of patients (7). To the authors' knowledge, this is the first study demonstrating successful reinstatement of NIS expression and therefore susceptibility to radioiodine uptake in ATC after systemic application of the therapeutic gene, an important prerequisite for clinical development.
Footnotes
Acknowledgments
We are grateful to Gerald Assmann (Institute of Pathology, LMU Munich, Munich, Germany) for assistance with staining and scoring of human ATC sections, and Wolfgang Rödl (Department of Pharmacy, LMU Munich, Munich, Germany) for conjugate synthesis.
This work was supported by grants from the Deutsche Forschungsgemeinschaft within the Collaborative Research Centre SFB 824 (project C8) to C.S. and the Priority Programme SPP1629 to C.S. (SP 581/6-1, SP 581/6-2) and P.J.N. (NE 648/5-2), as well as a grant from the Wilhelm-Sander-Stiftung (2014.129.1) to C.S.
Author Disclosure Statement
No competing financial interests exist.