
Abstract
Background:
Dysregulated DNA methylation in lymphocytes has been linked to autoimmune disorders. The aims of this study were to identify global DNA methylation patterns in patients with autoimmune thyroid diseases and to observe methylation changes after treatment for these conditions.
Methods:
A cross-sectional study was conducted, including the following patients: 51 with newly diagnosed Graves' disease (GD), 28 with autoimmune hypothyroidism (AIT), 29 with positive thyroid autoantibodies, and 39 matched healthy volunteers. Forty GD patients treated with radioiodine or antithyroid drugs and 28 AIT patients treated with L-thyroxine were followed for three months. Serum free triiodothyronine, free thyroxine, thyrotropin, thyroid peroxidase antibodies, thyroglobulin antibodies, and thyrotropin receptor antibodies were assayed using electrochemiluminescent immunoassays. CD3+ T and CD19+ B cells were separated by flow cytometry for total DNA and RNA extraction. Global DNA methylation levels were determined by absorptiometry using a methylation quantification kit. DNA methyltransferase (DNMT) expression levels were detected by real-time polymerase chain reaction.
Results:
Hypomethylation and down-regulated DNMT1 expression in T and B lymphocytes were observed in the newly diagnosed GD patients. Neither the AIT patients nor the positive thyroid autoantibodies patients exhibited differences in their global DNA methylation status or DNMT mRNA levels compared with healthy controls. Antithyroid drugs restored global methylation and DNMT1 expression in both T and B lymphocytes, whereas radioiodine therapy affected only T cells. L-thyroxine replacement did not alter the methylation or DNMT expression levels in lymphocytes. The global methylation levels of B cells were negatively correlated with the serum thyroid peroxidase antibodies in patients with autoimmune thyroid diseases.
Conclusions:
Hyperthyroid patients with newly diagnosed GD had global hypomethylation and lower DNMT1 expression in T and B lymphocytes. The results provide the first demonstration that antithyroid drugs or radioiodine treatment restore global DNA methylation and DNMT1 expression with concurrent relief of hyperthyroidism.
Introduction
A
DNA methylation, the most studied epigenetic modification pattern, occurs at position 5 of the cytosine ring in the context of CpG dinucleotides and is catalyzed by DNA methyltransferases (DNMTs). Three DNMTs determine the methylation status of DNA methylation in humans: DNMT3a and DNMT3b are responsible for de novo methylation, whereas DNMT1 is constitutively expressed and required for maintenance of methylation (11). In general, DNA methylation results in the downregulation of gene transcription (12). DNA methylation plays essential roles in the maintenance of normal lymphocyte function (13). Aberrant DNA methylation patterns or abnormal DNMT expression has been linked to various autoimmune disorders (14,15), including systemic lupus erythematosus (SLE) (16,17), rheumatoid arthritis (RA) (17,18), vitiligo (19), and type 1 diabetes (20). In these disorders, the methylation status of genomic DNA and specific autoimmunity-related gene foci is reduced, which results in the upregulation of gene expression and leads to autoreactivity. DNA methylation and DNMTs might be possible indicators of disease progression and therapeutic targets (21). The expression of DNMT1 mRNA in peripheral blood mononuclear cells (PBMCs) is reported to be associated with disease activity in patients with RA (18), and treatment with methotrexate can restore defective regulatory T-cell function by reducing methylation of the FoxP3 locus accompanying increased FoxP3 expression (22). Overexpression of DNMT1 in SLE patient-sourced CD4+ T cells can inhibit the autoimmune response and reduce immunoglobulin G production by reversing the abnormally low DNA methylation levels (23).
It was hypothesized that aberrant DNA methylation in lymphocytes might contribute to the pathogenesis of AITD. Therefore, the present study aimed to identify changes in the global DNA methylation status and DNMT mRNA levels in T and B cells from untreated AITD patients. It also aimed to analyze whether treatment-induced disease remission is associated with changes in methylation and DNMT expression. Given that thyroid hormones regulate gene expression and immune response in lymphocytes through both genomic and non-genomic mechanisms (24), as well as the fact that thyroid hormones are reported to induce changes in epigenetic modifications (25), the potential association between thyroid function and global DNA methylation was also explored in the present study.
Materials and Methods
Study groups and prospective cohorts
This study was conducted at the First Affiliated Hospital of China Medical University from August 2013 to October 2014. All research was approved by the Medical Ethics Committee of the First Affiliated Hospital of China Medical University, and all participants provided informed consent. The study recruited 51 patients with newly diagnosed GD, 28 patients with untreated AIT, and 29 euthyroid patients with positive thyroid antibodies (pTAb) in the endocrine clinic. Thirty-nine age- and sex-matched healthy volunteers were recruited from the medical examination center. The inclusion criteria for the participants were applied as previously described (3). Briefly, patients with overt hyperthyroidism, positive thyrotropin (TSH) receptor antibodies (TRAbs), positive thyroid peroxidase antibodies (TPOAbs) and/or thyroglobulin antibodies (TgAbs), and a diffuse goiter or normal thyroid volume on B-mode ultrasonography or thyroid palpation were enrolled in the GD group. Patients with overt hypothyroidism, positive TPOAbs and/or TgAbs, and diffuse goiter on B-mode ultrasonography or thyroid palpation were included in the AIT group. Euthyroid subjects with positive TPOAbs and/or TgAbs were recruited to the pTAb group, whereas those with normal thyroid function and negative TPOAbs and TgAbs were enrolled in the healthy control (HC) group. The following exclusion criteria were applied: acute or chronic infection or inflammatory illness, such as SLE, RA, psoriasis, vitiligo, diabetes mellitus, gout, viral hepatitis, or malignancy, and current medication use. Diagnosis and treatment decisions were made by qualified endocrinologists according to relevant clinical guidelines and references (26,27). None of the included patients received any treatment at baseline. L-thyroxine was given to the AIT patients to correct hypothyroidism immediately after diagnosis. The GD patients received either antithyroid drugs (methimazole or propylthiouracil) or radioiodine therapy. The subjects in the AIT group and GD group were followed after treatment for three months.
Venous blood samples were obtained from every subject between 8.00am and 10.00am after an overnight fast. Serum was separated, stored at −80°C, and assayed as soon as possible. Fresh whole-blood samples (approximately 10 mL) were collected in heparinized tubes for cell isolation.
Flow cytometry cell sorting
PBMCs were separated by Ficoll-Hypaque (Dakewei) density gradient centrifugation immediately after collection and washed twice with precooled phosphate-buffered saline. FITC-conjugated anti-CD3 (#561806; BD Pharmingen) and PE-conjugated anti-CD19 (#561741; BD Pharmingen) were used for cell surface staining according to the manufacturer's instructions. Cell sorting was performed on a FACS Arial™ II instrument (BD Pharmingen) as soon as possible to maintain cell viability. Experienced technicians were responsible for gating. The isolated CD3+ T- and CD19+ B-cell subpopulations were stored at −80°C until total DNA and RNA extraction.
Biochemical measurements
The serum levels of free triiodothyronine (fT3), free thyroxine (fT4), TSH, TPOAb, TgAb, and TRAb were measured through electrochemiluminescent immunoassays using a Cobas Elesys 601 instrument (Roche Diagnostics Ltd.). Reference ranges were obtained from the manufacturer and are presented in Tables 1 and 2.
The normal reference ranges were obtained from the manufacturer: fT3, 3.10–6.80 pmol/L; fT4, 12.00–22.00 pmol/L; TSH, 0.27–4.20 mIU/L; TPOAb, 0–34 IU/mL; TgAb, 0–115 IU/mL.
The variables are presented as the means ± SD.
The variables are presented as the medians with interquartile ranges. The p-values were obtained through statistical analyses using one-way ANOVA or Kruskal–Wallis test.
p < 0.05 compared to HCs; ** p < 0.01 compared to HCs.
AITD, autoimmune thyroid disease; F, female; M, male; GD, Graves' disease; AIT, autoimmune thyroiditis; pTAb, positive antithyroid autoantibodies; HC, healthy control; fT3, free triiodothyronine; fT4, free thyroxine; TSH, thyrotropin; TPOAb, thyroid peroxidase antibodies; TgAb, thyroglobulin antibodies; SD, standard deviation; ANOVA, analysis of variance.
The normal reference ranges were obtained from the manufacturer: fT3, 3.10–6.80 pmol/L; fT4, 12.00–22.00 pmol/L; TSH, 0.27–4.20 mIU/L; TPOAb, 0–34 IU/mL; TgAb, 0–15 IU/mL; TRAb, 0–1.75 IU/L.
The variables are presented as the means ± SD.
The variables are presented as the medians with interquartile ranges. A paired parametric test was used for comparisons of fT3, fT4, TPOAb, TgAb, and TRAb before and after treatment, and a paired nonparametric test was performed for comparing TSH before and after treatment.
p < 0.05 compared to before treatment; ** p < 0.01 compared to before treatment.
GD, Graves' disease; TRAb, TSH receptor antibodies.
DNA extraction and global DNA methylation measurements
Total DNA from the thawed CD3+ T and CD19+ B cells was extracted using a TIANamp Genomic DNA kit (Tiangen Biotech). The concentration and quality of the purified DNA were analyzed using a NanoDrop 2000C spectrophotometer (NanoDrop Technologies). Samples with an OD260/OD280 ratio between 1.8 and 2.0 were considered satisfactory and were used for experiments. Global DNA methylation was assayed by colorimetric means using a DNA methylation quantitative assay kit (A-P-1034-48; A&D Technology). Briefly, 100 ng of DNA was added and fixed on a strip well with a specific affinity for DNA. The DNA methylation levels were then quantified by a 5-methylcytosine capture antibody and a detection antibody. The amount of methylated DNA was proportional to the optical density (OD) values obtained from the enzyme-linked immunosorbent assays and is presented according to the calculated percentage of 5-mC. According to the manufacturer's instructions, the percentage of 5-mC was calculated as follows:
where S represents the initial input quantity of DNA samples, and P represents the quantity of the positive control. The mean OD450 for ME3 and ME4 are 0.075 and 0.675, respectively. Every sample was tested in duplicate. The standard curve samples and the positive and negative controls were assayed on the same plate.
Total RNA extraction and real-time polymerase chain reaction
Total RNA from the thawed CD3+ T and CD19+ B cells was extracted with the TRIzol reagent (Invitrogen). The RNA concentrations were determined using a NanoDrop 2000C spectrophotometer (NanoDrop Technologies), and sufficient quality was represented by an OD260/OD280 ratio between 1.8 and 2.0. A 20 μL reverse transcription system was constructed by adding 500 ng of RNA and 4 μL of 5 × PrimeScript RT reagent kit (#036A; TaKaRa), and cDNA was synthesized on a Veriti™ 96-well Thermal Cycler (AB Applied Biosystems). Subsequently, 2 μL of cDNA was added after fivefold dilution, and primers at a final concentration of 0.4 μM and SYBR Premix Ex Taq™ II (#820A; TaKaRa) were then added to obtain a total volume of 20 μL. Real-time polymerase chain reaction (PCR) amplification was performed on a LightCycler 480 Real-Time PCR System (Roche). GAPDH was used as an internal reference. Specific primers were designed and synthesized by TaKaRa Biotech as follows: DNMT1-F, 5′-CGG TTC CTC CTG GAG AAT GTC A-3′; DNMT1-R, 5′-CAC TGA TAG CCC ATG CGG ACC A-3′; DNMT3a-F, 5′-CAA TGA CCT CTC CAT CGT CAA C-3′; DNMT3a-R, 5′-CAT GCA GGA GGC GGT AGA A-3′; DNMT3b-F, 5′-CCC ATT CGA GTC CTG TCA TT-3′; DNMT3b-R, 5′-GGT TCC AAC AGC AAT GGA CT-3′; GAPDH-F, 5′-ACA TCG CTC AGA CAC CAT G-3′; and GAPDH-R, 5′-TGT AGT TGA GGT CAA TGA AGG G-3′. Each sample was tested in triplicate. A melting curve was generated after amplification to verify the absence of primer dimers or incorrect product formation. The mRNA expression level of the target genes was normalized to GAPDH and calculated using the 2–ΔCp method.
Statistical analysis
IBM SPSS Statistics for Windows v21.0 (IBM Corp.) was used for all statistical analyses. Normally distributed quantitative variables are described as the means ± standard deviations. Non-normally distributed variables are expressed as the medians with interquartile ranges. Categorical variables are expressed by frequencies. Comparisons for fT3, fT4, TPOAb, and TgAb among all groups at baseline were conducted by one-way analysis of variance followed by Bonferroni correction for paired comparisons. Comparison for TSH between all groups at baseline was conducted by a Kruskal–Wallis test. A paired parametric or nonparametric test was conducted to analyze the changes in fT3, fT4, TSH, autoantibodies, global methylation, and DNMT expression that occurred due to treatment. Student's t-test was used to compare DNA methylation and DNMT expression after treatment to that of HCs. Pearson's chi-square test was applied for comparisons of the sex ratios. A bivariate correlation analysis was performed using the Spearman rank correlation method. A p-value of <0.05 was considered statistically significant, and all calculated p-values were two-sided.
Results
Global DNA methylation and DNMT mRNA expression levels in CD3+ T cells and CD19+ B cells from AITD patients and HCs
The subjects' demographic and clinical characteristics are presented in Table 1. There were no statistically significant differences in terms of age or sex ratio between the groups, indicating a comparable baseline. The GD patients exhibited lower global methylation in CD3+ T cells (0.734 ± 0.293 vs. 1.636 ± 0.371; p < 0.001) and CD19+ B cells (0.165 ± 0.069 vs. 0.817 ± 0.172; p < 0.001) compared to the HCs. The global DNA methylation levels in the CD3+ T cells and CD19+ B cells from the AIT group (p = 0.375 for T cell and p = 0.103 for B cell) and the pTAb group (p = 1.00 for T cell and p = 1.00 for B cell) did not differ from those of the control group (Fig. 1A and B).
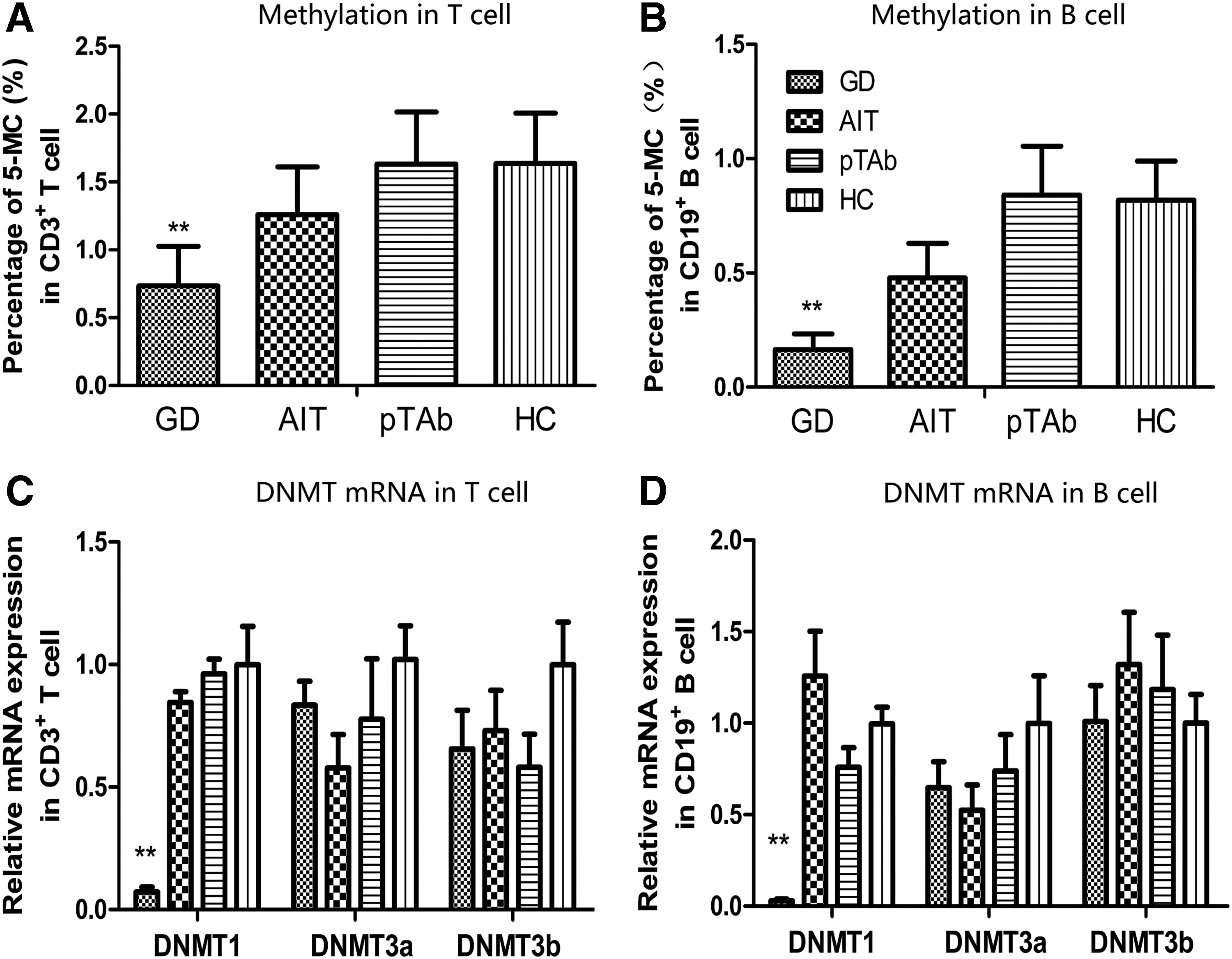
Global DNA methylation and DNMT expression in CD3+ T cells and CD19+ B cells of patients with autoimmune thyroid disease (AITD) and HCs. Global DNA methylation levels in T cells (
The DNMT1 mRNA expression levels in both CD3+ T cells (0.072 ± 0.020 vs. 0.999 ± 0.156; p < 0.001) and CD19+ B cells (0.029 ± 0.010 vs. 1.000 ± 0.091; p < 0.001) of GD patients were significantly lower compared with those of the HCs. There were no significant differences in DNMT3a (p = 1.00 for T cell and p = 0.101 for B cell) or DNMT3b (p = 0.345 for T cell and p = 1.00 for B cell) expression between the GD patients and controls. No difference in DNMT mRNA expression was found between AIT patients and HCs (p = 1.00 for DNMT1, p = 1.00 for DNMT3a, p = 0.92 for DNMT3b in T cells; p = 1.00 for DNMT1, p = 0.509 for DNMT3a, p = 0.745 for DNMT3b in B cells) or between pTAb patients and HCs (p = 1.00 for DNMT1, p = 1.00 for DNMT3a, p = 0.352 for DNMT3b in T cells; p = 1.00 for DNMT1, p = 1.00 for DNMT3a, p = 1.00 for DNMT3b in B cells; Fig. 1C and D).
Restoration of global DNA methylation and DNMT1 expression in CD3+ T cells and CD19+ B cells accompanied by relief of hyperthyroidism in the GD patients after treatment
Forty GD patients and 28 AIT patients were successfully followed after three months of clinical treatment. In the GD group, 15 patients received antithyroid drugs, and 25 were treated with radioiodine therapy. All individuals in the AIT group were given L-thyroxine replacement therapy. The corresponding changes in thyroid function and serum antithyroid antibodies are summarized in Table 2. In brief, hyperthyroidism and hypothyroidism were effectively treated in the entire GD and AIT cohorts, respectively. However, the levels of serum autoantibodies were unchanged after treatment.
As shown in Figure 2A and B, a significant increase in global DNA methylation levels in both CD3+ T cells (1.314 ± 0.618 vs. 0.739 ± 0.268; p < 0.001) and CD19+ B cells (0.486 ± 0.287 vs. 0.166 ± 0.059; p = 0.004) was found in GD patients after treatment compared to the pretreatment levels. After treatment, the DNMT1 mRNA levels in CD3+ T cells (0.775 ± 0.023 vs. 0.064 ± 0.018; p < 0.001) and CD19+ B cells (1.075 ± 0.114 vs. 0.031 ± 0.024; p < 0.001) were significantly elevated in all of the GD patients (Fig. 2C and D). Moreover, compared to the HCs, there was no significant difference in global DNA methylation (p = 0.124 in T cell and p = 0.071 in B cell) and DNMT1 expression (p = 0.247 in T cell and p = 0.956 in B cell) in T and B lymphocytes from GD patients after treatment. In contrast, no significant changes in DNMT3a (p = 0.365 for T cell and p = 0.268 for B cell) or DNMT3b (p = 0.486 for T cell and p = 0.815 for B cell) mRNA expression were detected after treatment.
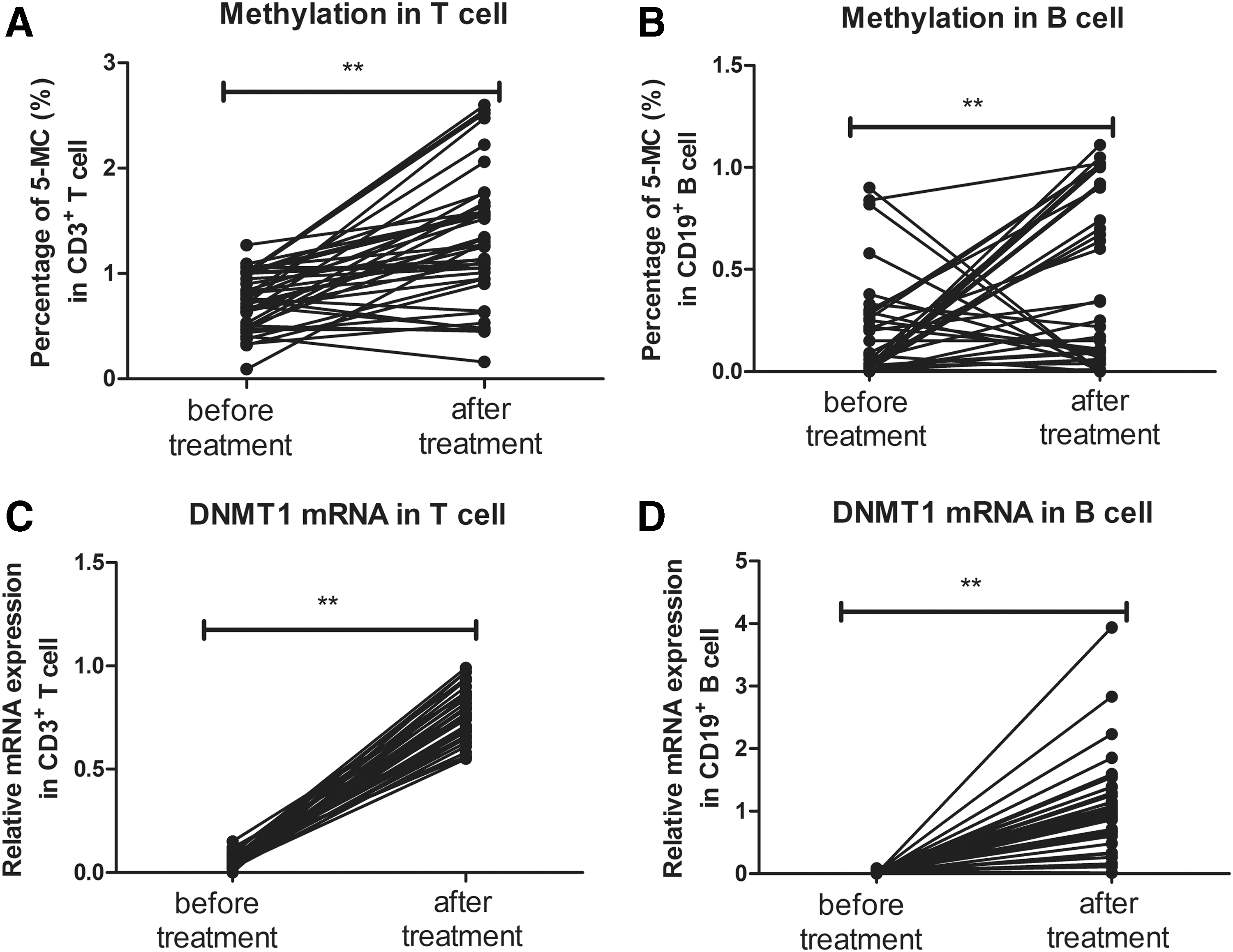
Changes in global DNA methylation and DNMT1 mRNA expression in CD3+ T cells and CD19+ B cells from GD patients before and after treatment. Global DNA methylation levels in T cells (
Compared to the pretreatment global DNA methylation status, there were no significant changes in T cells (p = 0.772) or in B cells (p = 0.192) from AIT patients after L-thyroxine replacement. Furthermore, the mRNA expression levels of DNMTs in both T cells (p = 0.362 for DNMT1, p = 0.219 for DNMT3a, and p = 0.484 for DNMT3b) and B cells (p = 0.695 for DNMT1, p = 0.187 for DNMT3a, and p = 0.732 for DNMT3b) did not change significantly.
Changes in global DNA methylation and DNMT1 expression in CD3+ T cells and CD19+ B cells after antithyroid drug therapy and radioiodine therapy
An upward trend in DNMT1 expression was observed in GD patients after treatment (Fig. 2). However, global methylation increased in some patients but decreased in others. The question was raised of whether antithyroid drugs and radioiodine might exert different effects on global methylation in lymphocytes. Both therapies significantly reduced fT3 and fT4 levels, whereas TSH levels were rising. Antithyroid drugs significantly decreased the serum TRAb levels (before 18.69 ± 6.33 vs. after 8.43 ± 8.00; p < 0.01), whereas radioiodine therapy increased these levels (before 21.44 ± 8.59 vs. after 27.52 ± 13.38; p = 0.058; Fig. 3A and B).
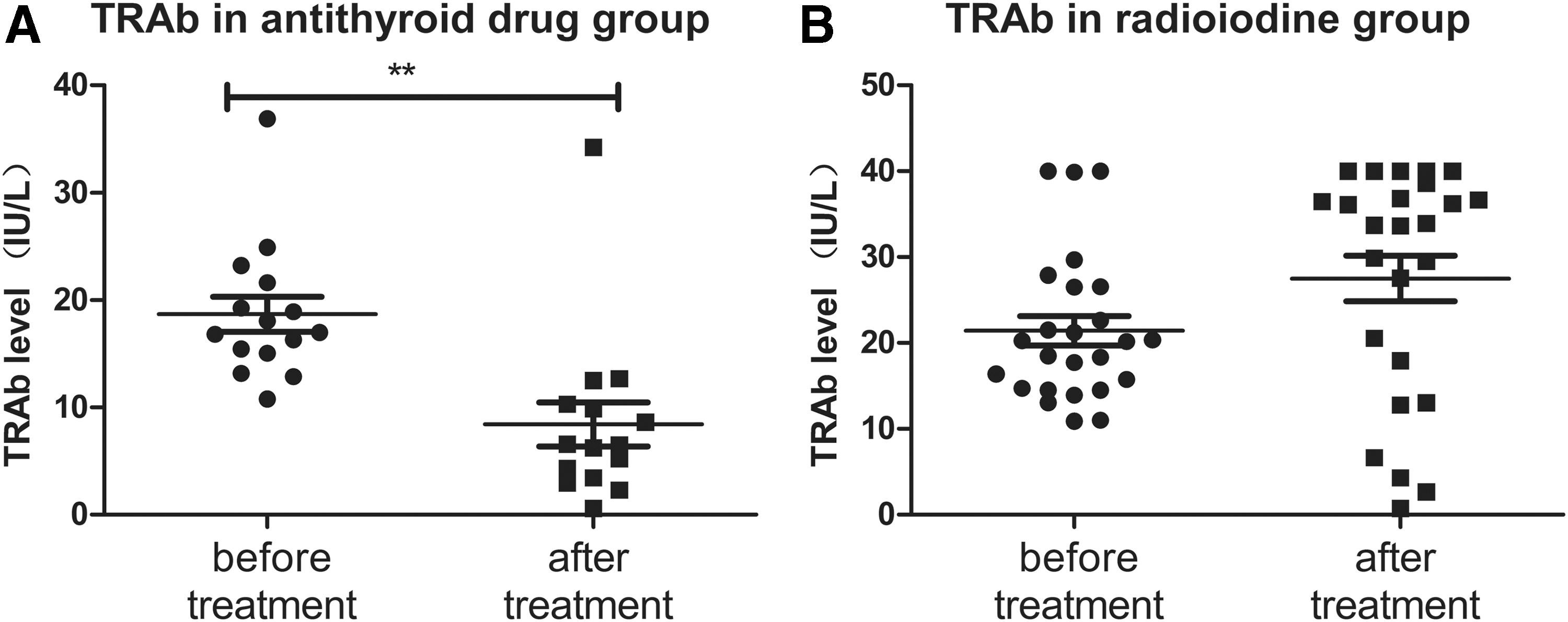
Changes in thyrotropin receptor antibodies (TRAb) in GD patients before and after treatment with antithyroid drugs or radioiodine. TRAb levels in GD patients administered with antithyroid drugs (
The global DNA methylation levels in CD3+ T cells (before 0.739 ± 0.249 vs. after 1.289 ± 0.776; p = 0.007) and CD19+ B cells (before 0.122 ± 0.057 vs. after 0.851 ± 0.126; p = 0.006) increased significantly after treatment with antithyroid drugs (Fig. 4A and B). The post-treatment levels did not differ from those measured in HCs (p = 0.181 for T cells and p = 0.362 for B cells). Radioiodine treatment increased the global DNA methylation levels in CD3+ T cells (before 0.739 ± 0.284 vs. after 1.330 ± 0.519; p = 0.001) significantly but had little effects on CD19+ B cells (before 0.202 ± 0.051 vs. after 0.343 ± 0.063; p = 0.230; Fig. 4C and D). The global methylation levels in B cells post radioiodine treatment remained lower than those in HCs (p = 0.034). Both antithyroid drugs and radioiodine treatment increased DNMT1 mRNA expression in T and B cells (Fig. 4E–H).
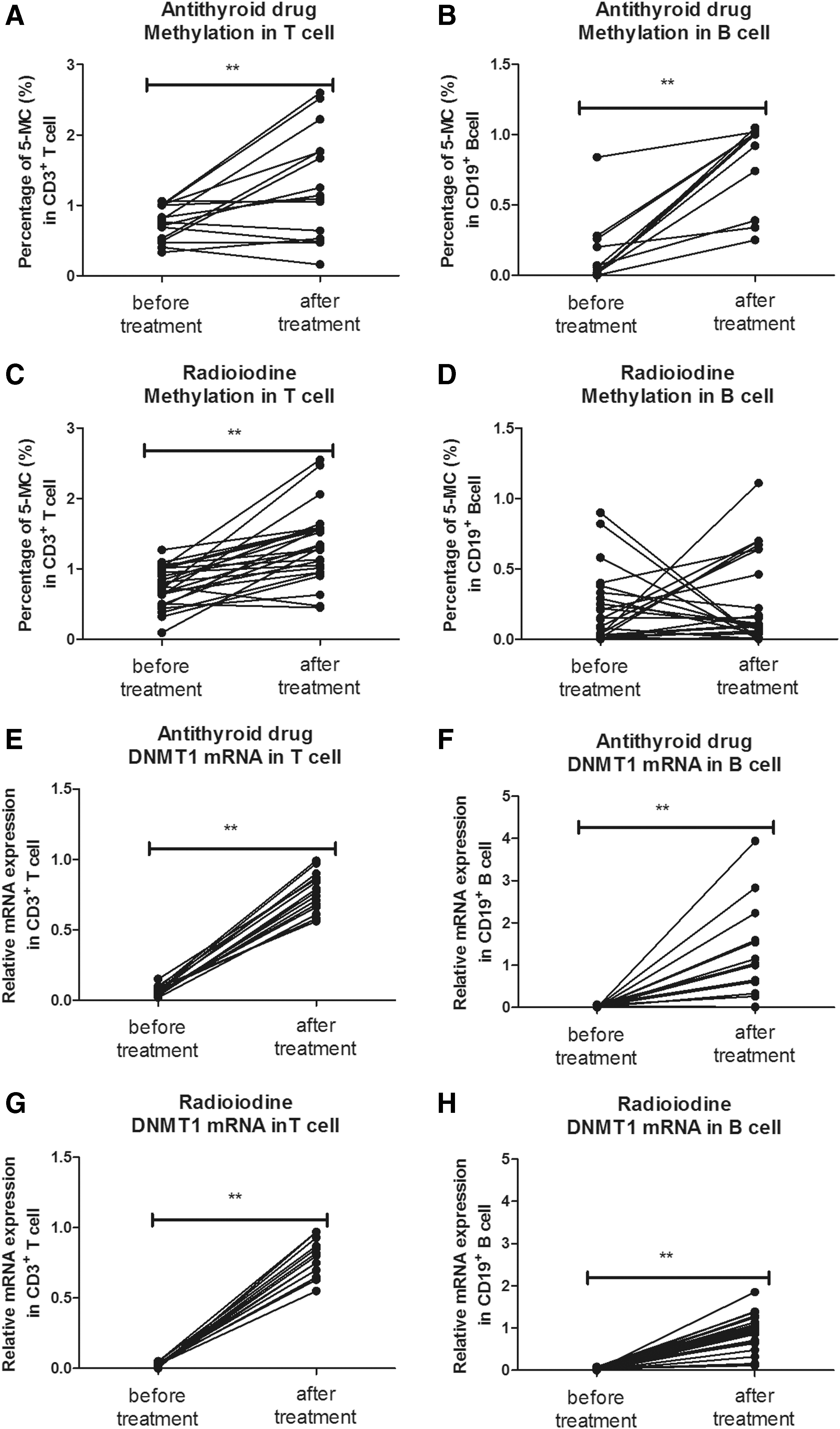
Changes in global DNA methylation and DNMT1 mRNA expression in CD3+ T cells and CD19+ B cells from GD patients after treatment with antithyroid drugs or radioiodine. Global DNA methylation levels in T cells (
Correlation among global DNA methylation, DNMT mRNA expression and serum fT3, fT4, TSH, TPOAb, TgAb, and TRAb levels
At baseline, no significant correlations were found between the global DNA methylation levels and the serum fT3, fT4, TSH, TPOAb, TgAb, or TRAb levels in any of the groups, and this remained unchanged after three months of treatment in the GD and AIT patients. The TRAb levels did not correlate with global DNA methylation in T cells and B cells, irrespective of whether the patients were treated with antithyroid drugs or radioiodine.
The results from GD, AIT, and pTAb patients at baseline revealed a negative association between global methylation in B cells and serum TPOAb (r = −0.294, p = 0.024, n = 108). Among these AITD patients, the DNMT1 mRNA levels were significantly correlated with global DNA methylation in CD3+ T cells (r = 0.476, p < 0.001, n = 147) and CD19+ B cells (r = 0.408, p < 0.001, n = 147).
Discussion
The present study demonstrates the characteristics of global DNA methylation and DNMT expression in lymphocytes from AITD patients. Global hypomethylation and downregulated DNMT1 mRNA expression were observed in T and B lymphocytes in patients with newly diagnosed GD. However, neither the AIT patients nor the pTAb patients exhibited differences in global DNA methylation status or DNMT mRNA levels compared to the HCs. Treatment with antithyroid drugs or radioiodine for three months restored the global DNA methylation and DNMT1 mRNA levels in lymphocytes from patients with GD. L-thyroxine replacement did not alter the methylation status or DNMT expression in lymphocytes from AIT patients.
Two possible mechanisms might explain the findings, and they might coexist and work together. First, hypomethylation in lymphocytes might contribute to the pathogenesis of AITD. Studies by Richardson et al. have indicated that inhibition of DNA methylation in T lymphocytes causes autoreactivity in vitro (28) and lupus-like pathological changes in vivo (29). In addition, DNA hypomethylation in lymphocytes has been observed in patients with RA and active SLE (17), as well as other autoimmune disorders (15). These findings suggest a possible relationship between DNA hypomethylation, lymphocyte hyperfunction, and autoimmunity. Although complicated mechanisms are involved, it is assumed that newly diagnosed GD patients are at the stage of immune activation in which autoreactive lymphocytes respond to endogenous thyroid antigens and undergo vigorous differentiation and proliferation (5), whereas euthyroid or hypothyroid patients are in remission or at a terminal stage of immune attack (2). The global hypomethylation that only occurred in newly diagnosed GD patients might be a potential cause of immune activation. In addition, hyperthyroidism might be a simultaneous consequence of an immune attack on the thyroid tissue. Additionally, both antithyroid drugs (30) and radioiodine therapy (31) are reported to regulate immune activity and differentiation in lymphocytes. In the current study, meaningful restoration of global DNA methylation and DNMT1 expression was observed in GD patients after treatment, which might add a possible mechanism contributing to treatment-induced disease remission. It should be noted that antithyroid drugs and radioiodine therapy might work through different epigenetic modifications, such as global DNA methylation, in CD19+ B lymphocytes, which in turn leads to different effects on TRAb production.
As a ubiquitous epigenetic modification pattern, global methylation is influenced by various endogenous and exogenous factors (11). It is hypothesized that high levels of thyroid hormone might be one of these factors. This hypothesis is based on the findings that low levels of global methylation and DNMT1 expression were only observed in patients with hyperthyroidism, even though autoantibodies were present in all AITD patients. Furthermore, the increases in global methylation and DNMT1 expression in GD patients detected after treatment were accompanied by a decrease in thyroid hormones in the blood, whereas the serum TPOAb and TgAb remained at stable levels. Moreover, whether antithyroid drugs and radioiodine regulate immune activity by reducing thyroid hormones or through direct actions on lymphocytes remains controversial (30). To the best of the authors' knowledge, thyroid hormones are important enhancers of lymphocyte function (24,32). It was speculated that some epigenetic mechanisms, such as DNA methylation, might be involved. In a previous study, reactive oxygen species (ROS) generation was found to be threefold higher in PBMCs from hyperthyroid patients compared to euthyroid and hypothyroid individuals (33). It has also been reported that ROS can induce global hypomethylation in cancer cells by transforming 5-mC to 5-hmc and guanine to 8-OHdG (8-OHdG serves as a measure of DNA oxidative damage) (34). In this study, a slight change in global methylation was observed in CD19+ B cells after radioiodine treatment. This outcome might have been induced simultaneously by decreased thyroid hormones and enhanced release of autoantigens because radioiodine exerts direct damaging effects on thyroid tissue (35).
A few studies have indicated that aberrant DNA methylation or gene polymorphism of methylation-related enzymes might participate in the pathogenesis of AITD. Limbach et al. investigated the genome-wide DNA methylation of CD4+ and CD8+ T cells in GD patients with a median time after initial diagnosis of 12 years. They found that genes involved in T-cell activation and signaling were hypermethylated (36). Although the present study also investigated GD patients, the focus was on newly diagnosed patients, and a follow-up was performed to observe eventual changes. The results revealed global hypomethylation in lymphocytes at diagnosis followed by restoration after treatment. The discrepancies between the study by Limbach et al. and the present findings might be due to the differences in inclusion criteria. Similar to the current findings, a recent study by Cai et al. found a significant decrease in DNMT1 mRNA transcription in PBMCs from untreated hyperthyroid GD patients (37). The use of purified CD3+ T cells and CD19+ B cells in the present experiments likely further reduced confounding factors caused by heterogeneity of cellular blood components. Another study reported that the DNMT1 +32204GG genotype correlated with DNA hypomethylation and was more frequently observed in patients with intractable GD than in patients with GD in remission (38). Regrettably, the authors did not mention whether there was a difference in the methylation status between intractable patients and individuals going into remission (duration of treatment was approximately 12 years).
This study has several limitations. First, the three-month post-treatment reevaluation time point is early. Given that euthyroidism was not achieved in all patients, different estimates might be achieved after a longer follow-up. Second, we did not perform further studies on patients with non-autoimmune hyperthyroidism or research addressing mechanisms in vitro with cultured lymphocytes stimulated with thyroid hormone. Such studies may help to elucidate further the cross-talk between thyroid hormones and epigenetic modifications of the immune system. Moreover, the levels of inflammatory cytokines, which are related to the immune activity of lymphocytes, were not measured due to the lack of sufficient serum and RNA samples.
In conclusion, global hypomethylation and lower DNMT1 expression were observed in T and B lymphocytes of hyperthyroid patients with newly diagnosed GD. Neither AIT patients nor pTAb patients exhibited differences in the global DNA methylation status or DNMT mRNA levels compared to HCs. This study provides the first demonstration that antithyroid drugs or radioiodine restore global DNA methylation and DNMT1 expression through treatment of hyperthyroidism. This study highlights the participation of hypomethylated lymphocytes in the pathogenesis of GD and a novel epigenetic-modifying effect of antithyroid drugs and radioiodine.
Footnotes
Acknowledgments
This work was supported by the National Natural and Science Foundation of China (grant numbers: 81570708, 81430018, and U1508219) and the Liaoning Provincial Key Laboratory Foundation of the Ministry of Education (LZ2014035). We thank all of the volunteers who participated and donated samples in this study. We also thank physicians and technicians from the Institute of Endocrinology in the First Affiliated Hospital of China Medical University for the advice and assistance provided.
Author Disclosure Statement
No competing financial interests exist.