
Abstract
Background:
Thyroid hormones are essential for normal development of the central nervous system (CNS). Experimental rodents have shown that even a subtle thyroid hormone insufficiency in circulating maternal thyroid hormones during pregnancy may adversely affect neurodevelopment in offspring, resulting in irreversible cognitive deficits. This may be due to the persistent reduced expression of the hippocampal brain-derived neurotrophic factor gene Bdnf, which plays a crucial role in CNS development. However, the underlying molecular mechanisms remain unclear.
Methods:
Thiamazole (MMI; 0.025% [w/v]) was administered to dams from two weeks prior to conception until delivery, which succeeded in inducing mild maternal hypothyroxinemia during pregnancy. Serum thyroid hormone and thyrotropin levels of the offspring derived from dams with mild maternal hypothyroxinemia (M offspring) and the control offspring (C offspring) were measured. At 70 days after birth, several behavior tests were performed on the offspring. Gene expression and DNA methylation status were also evaluated in the promoter region of Bdnf exon IV, which is largely responsible for neural activity-dependent Bdnf gene expression, in the hippocampus of the offspring at day 28 and day 70.
Results:
No significant differences in serum thyroid hormone or thyrotropin levels were found between M and C offspring at day 28 and day 70. M offspring showed an impaired learning capacity in the behavior tests. Hippocampal steady-state Bdnf exon IV expression was significantly weaker in M offspring than it was in C offspring at day 28. At day 70, hippocampal Bdnf exon IV expression at the basal level was comparable between M and C offspring. However, it was significantly weaker in M offspring than in C offspring after the behavior tests. Persistent DNA hypermethylation was also found in the promoter region of Bdnf exon IV in the hippocampus of M offspring compared to that of C offspring, which may cause the attenuation of Bdnf exon IV expression in M offspring.
Conclusions:
Mild maternal hypothyroxinemia induces persistent DNA hypermethylation in Bdnf exon IV in offspring as epigenetic memory, which may result in long-term cognitive disorders.
Introduction
T
On the other hand, subclinical hypothyroidism, defined as normal serum free T4 (fT4) and elevated TSH levels, or hypothyroxinemia, defined as normal serum TSH and low fT4 levels, in pregnancy may be associated with adverse outcomes, including a lower-than-normal intelligence quotient (IQ) in offspring. However, this remains controversial (7 –9). Animal studies previously demonstrated that maternal subclinical hypothyroidism or hypothyroxinemia had a significant negative impact on neurodevelopment. However, the molecular mechanisms by which mild maternal hypothyroid disorders, such as subclinical hypothyroidism and hypothyroxinemia, during pregnancy affect CNS development and cognitive functions in offspring remain unclear.
Brain-derived neurotrophic factor (BDNF) and its high-affinity receptor, tropomyosin-related receptor kinase B (TrkB), are widely expressed in the mammalian brain (10) and play a crucial role in the development, maintenance, and functioning of the CNS (11). In adulthood, BDNF modulates synaptic plasticity and regulates cognitive brain function, including learning and memory. The BDNF gene consists of nine exons (I–IX) in rodents and humans, and Bdnf exon IV is largely responsible for neural activity-dependent Bdnf expression in rodents (12). BDNF gene expression is positively regulated by thyroid hormone in vivo (13 –15), and hippocampal BDNF expression was previously shown to be reduced in offspring derived from dams with mild hypothyroidism during pregnancy, suggesting that reduced hippocampal BDNF expression induced by thyroid hormone insufficiency in the developing brain contributes to adverse functional and structural events in the CNS (13,16,17). However, the underlying molecular mechanisms of persistent reductions in hippocampal BDNF expression are poorly understood.
Since DNA methylation is a major epigenetic modification known to suppress gene transcription in the long term, and prenatal stress has been reported to increase DNA methylation and reduce the expression of Bdnf exon IV (12), it was hypothesized that mild maternal hypothyroidism during pregnancy induces DNA hypermethylation of Bdnf exon IV in the fetal brain, thereby reducing Bdnf expression in the long term, which may cause cognitive disorders in adulthood.
This study reports that mild maternal hypothyroxinemia during pregnancy in mice induced an impaired learning capacity and persistent DNA hypermethylation in the transcriptionally relevant region of Bdnf exon IV, thereby resulting in the attenuation of hippocampal Bdnf exon IV expression in the offspring.
Materials and Methods
Animals
All animal experiments were performed in accordance with the guidelines of the Tokyo Medical and Dental University Committee on Animal Research (#A2017032). Eight-week-old C57BL6 mice were purchased from Sankyo Lab Service (Tokyo, Japan). Female adult C57BL/6J mice were maintained at 22°C in a room with an automatic 12-hour light/dark cycle. These mice were randomly separated into two groups, and the mean body weight (BW) of each group was similar. Thiamazole (1-methy-2-merucaptoimidazole; MMI) was orally administered to one group, referred to as Dam M (18) (Fig. 1A). The MMI dose (0.025% [w/v]) administered to dams employed in this study has been reported to induce mild hypothyroidism without morphological changes in mouse offspring (19,20). Water without MMI was administered to control females (Dam C). All female mice were mated with wild-type male mice two weeks after the MMI treatment. The offspring from Dam M and Dam C were referred to as “M offspring” and “C offspring,” respectively. Male offspring were weaned 21 days after birth and subjected to behavioral tests at day 70 (Fig. 1A). Serum free T3 (fT3) and fT4 levels were assessed using an electrochemiluminescence immunoassay (ECLIA; SRL, Tokyo, Japan). Serum TSH levels of dams were measured by a specific mouse TSH radioimmunoassay, as previously described (21,22), with mouse TSH/luteinizing hormone (LH) reference (AFP51718MP), mouse TSH antiserum (AFP98991), and rat TSH antigen (NIDDK-rTSH-I-9), all of which were obtained from Dr. A.F. Parlow (Harbor–University of California at Los Angeles Medical Center, Torrance, CA). The detection range of the assay was 0.078–10 ng TSH/mL. The intra-assay variation was <6%, and all samples were measured in one series to prevent inter-assay variation. Serum TSH levels of offspring were measured with the Mouse TSH ELISA Kit (#MBS762347; MyBioSource, Inc., San Diego, CA).
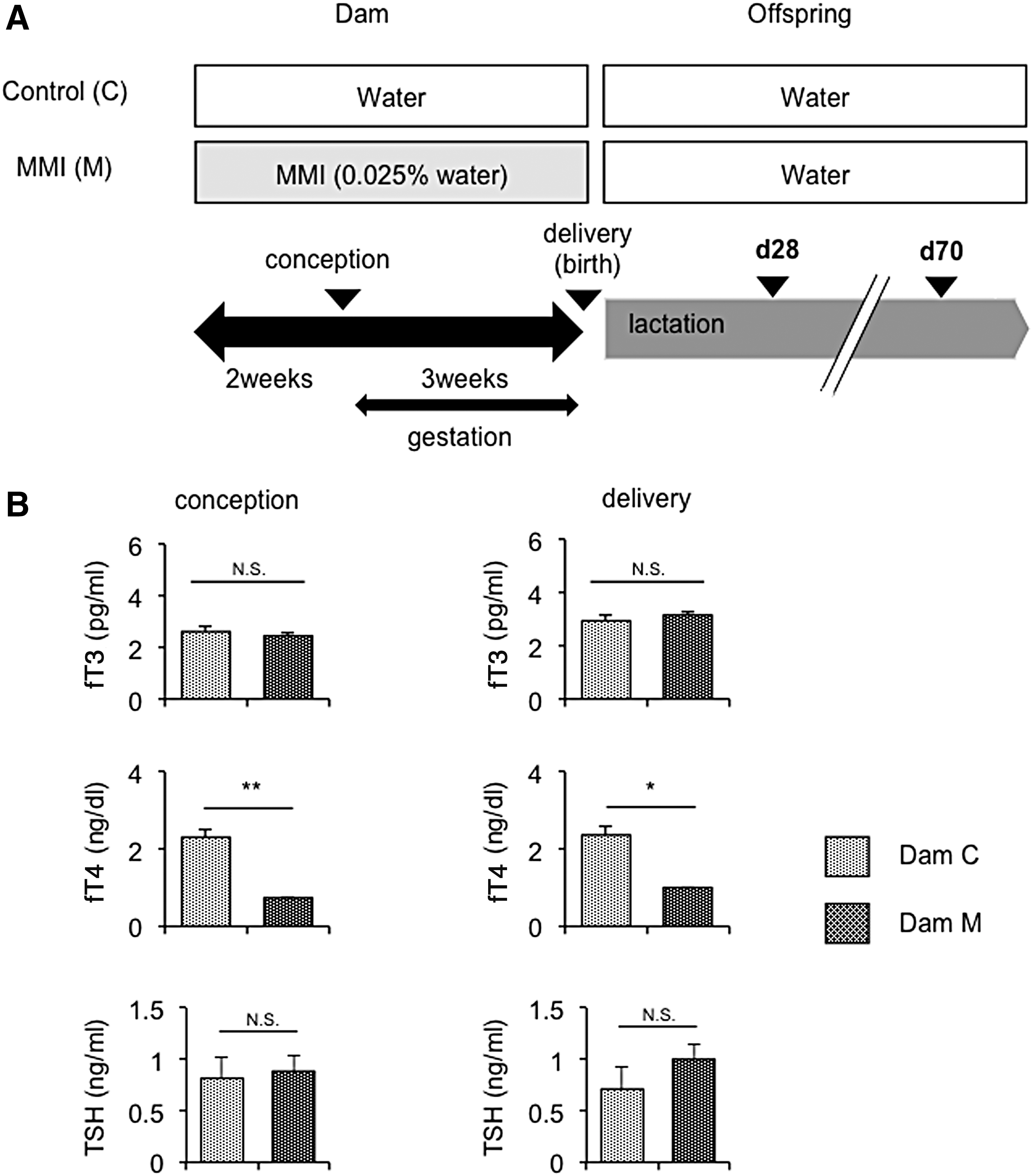
Induction of mild maternal hypothyroxinemia. (
Rotarod test
The rotarod test was performed, as previously described (23). Briefly, mice were placed on a rotating rod (diameter 3 cm), and the rotating speed was linearly increased from 3.5 rounds per minute (rpm) to 35 rpm in 300 s and continued at 35 rpm until 360 s. Mice were subjected to three trials (one trial per day for three consecutive days), with a 30- to 60 min rest interval between trials. The amount of time for each mouse to fall from the rod was recorded for each trial. The mean latency to fall off the rotarod was recorded and used in subsequent analyses.
Morris water maze test
The Morris water maze test was conducted in a circular pool that was 1 m in diameter and filled with water at a temperature of 22.0 ± 1°C. The water was colored using white paint, which prevented the mice from seeing the platform (20 cm high, 10 cm in diameter; 1 cm below the surface of the water) or other cues under the water. There were some extra-maze landmark cues that were visible to mice in the maze. Mice were subjected to four trials per day for five consecutive days. Each acquisition trial was initiated by placing an individual mouse into the water facing the outer edge of the maze at one of four designated starting points quasi-randomly, while the submerged platform remained constant for each mouse throughout testing. A trial was terminated when the mouse reached the platform, and the latency and distance swam were measured. The cutoff time of the trial was 60 s, and mice that did not reach the platform within 60 s were removed from the water and placed on the platform for 30 s before being toweled off and placed back into their home cage. The inter-trial interval was approximately 6 min. The distance swam, number of crossings, position of the target platform and other three platforms, and time spent in the quadrants of the four platforms were measured.
Contextual fear conditioning test
The contextual fear conditioning test was performed, as previously described (23), and was conducted one day after the Morris water maze test. This test consisted of two parts: conditioning and cued test trials. Fear conditioning was performed on a clear plastic chamber equipped with a stainless-steel grid floor (34 cm × 26 cm; 30 cm high). A CCD camera was equipped on the ceiling of the chamber and connected to a video monitor and computer. The grid floor was wired to a shock generator. White noise (80 dB) was supplied from a loudspeaker as an auditory cue (the conditioned stimulus [CS]). A continuous 0.4 mA foot shock (the unconditioned stimulus [US]) for 2 s was administered at the end of the 30 s CS period. The conditioning trial consisted of a 2 min exploration period followed by two CS–US pairings separated by 1 min each. A cued test was performed in the same conditioning chamber (80 dB white noise, 30 s) in the absence of the foot shock 24 h after the conditioning trial. The rate of the freezing response and moving distance of the mouse were measured as an index of fear memory.
Histological analysis
On day 28 and day 70, mice were deeply anesthetized and sacrificed. Brains, livers, and blood were removed from each mouse. Mouse brains were fixed with 4% paraformaldehyde in 0.1 M of phosphate buffer, pH 7.4, embedded in paraffin, and sectioned into slices 5 μm thick. Sections were stained with hematoxylin and eosin. For immunohistochemistry, paraffin-embedded mouse sections were deparaffinized, rehydrated, and then microwaved in 10 mM of citrate buffer, pH 6.0, for 15 min. These sections were incubated with primary antibodies at 4°C overnight and finally with Alexa Fluor 546-labeled anti-IgGs (#A-11035; Invitrogen, Paisley, United Kingdom) at room temperature for 1 h. Primary antibodies of anti-BDNF (#AB1534SP; Chemicon, Temecula, CA) were diluted at a ratio of 1:300. Sections were then washed in phosphate-buffered saline, dried, and mounted with Vectashield Mounting Medium with DAPI (#H-1200, Vector Laboratories, Burlingame, CA).
Extraction of genomic DNA and total RNA
Mouse brain and liver genomic DNA and total RNA samples were extracted on day 28 and day 70 by the All DNA/RNA preparation kit (Qiagen, Hilden, Germany). Different mice were used to extract RNA samples at day 70 without and after the last behavior test.
Quantitative real-time PCR analysis
Quantitative real-time polymerase chain reaction (qPCR) was performed, as described below. Total RNA was prepared using Sepazol™ (Nacalai Tesque, Kyoto, Japan). cDNA was synthesized from 5 μm of total RNA using Superscript II reverse transcriptase (Invitrogen) with random primers. Gene expression levels were measured with ABI PRISM 7700 using SYBR Green PCR Core Reagents (Applied Biosystems, Foster City, CA). mRNA levels were normalized to that of 36B4 and analyzed by the comparative CT method. The quantification data were analyzed with ABI StepOne software v2.3 (Applied Biosystems). The primer sets used for qPCR are described in Supplementary Table S1 (Supplementary Data are available online at
Bisulfite DNA methylation analysis
The sodium bisulfite treatment of genomic DNA was performed with a BisulFast™ DNA modification kit (Toyobo, Tokyo, Japan) according to the manufacturer's instructions. Bisulfite-treated genomic DNA was amplified by PCR using primers listed in Supplementary Table S2 for the promoter region of mouse Bdnif gene exon IV (24). PCR conditions were 96°C for 1 min and 40 cycles of 96°C for 15 s, 56°C for 30 s, and 72°C for 1 min, followed by 7 min for 72°C.
Bisulfite DNA methylation analysis of the promoter region of mouse glutamic acid decarboxylase 1 (Gad1) and reelin (Reln) genes was performed, as described in a previous study using PCR primers listed in Supplementary Table S2 (25). Twenty-four bacterial colonies were picked up per PCR amplification. A web-based tool, a quantification tool for the methylation analysis, was used for the bisulfite sequencing analysis of CpG methylation (
Statistical analysis
Data are expressed as the mean ± standard error of the mean. Data were compared using Student's t-test or two-way analysis of variance with Bonferroni post hoc test. A p-value of <0.05 was considered to be significant. Statistical analysis was performed using GraphPad Prism v6 (GraphPad Software, Inc., La Jolla, CA).
Results
MMI treatment induced mild maternal hypothyroxinemia during pregnancy
MMI (0.025% [w/v]; Dam M) or water (Dam C) was administered to dams from two weeks prior to conception until delivery (for approximately five weeks; Fig. 1A). Serum fT4 levels were significantly lower in Dam M than in Dam C at conception and delivery, whereas no significant difference in serum fT3 levels was observed between Dam C and Dam M at these time points (Fig. 1B). Serum TSH levels were also similar between the two dams at both conception and delivery (Fig. 1B). Thus, the MMI treatment induced mild hypothyroxinemia in Dam M. Since T3 is the primary biologically active form of thyroid hormone that binds to and activates thyroid hormone receptors (27), normal serum fT3 and low fT4 levels suggested the successful induction of mild maternal hypothyroxinemia in Dam M.
Mild maternal hypothyroxinemia did not affect growth or thyroid function in offspring
Body weight, whole brain weight, and thyroid function were examined in M and C offspring. No significant differences were observed in body weight at birth and thereafter or whole brain weight between M and C offspring at day 28 and day 70 (Fig. 2A and B). No significant differences were found in serum fT3, fT4, or TSH levels between M and C offspring at day 28 and day 70. However, serum fT4 and fT3 levels in M offspring were significantly decreased relative to those in C offspring at day 1 (Table 1). Therefore, it is conceivable that even though normalized in adulthood, thyroid hormone levels in offspring derived from dams with mild maternal hypothyroxinemia are decreased during the gestational period.
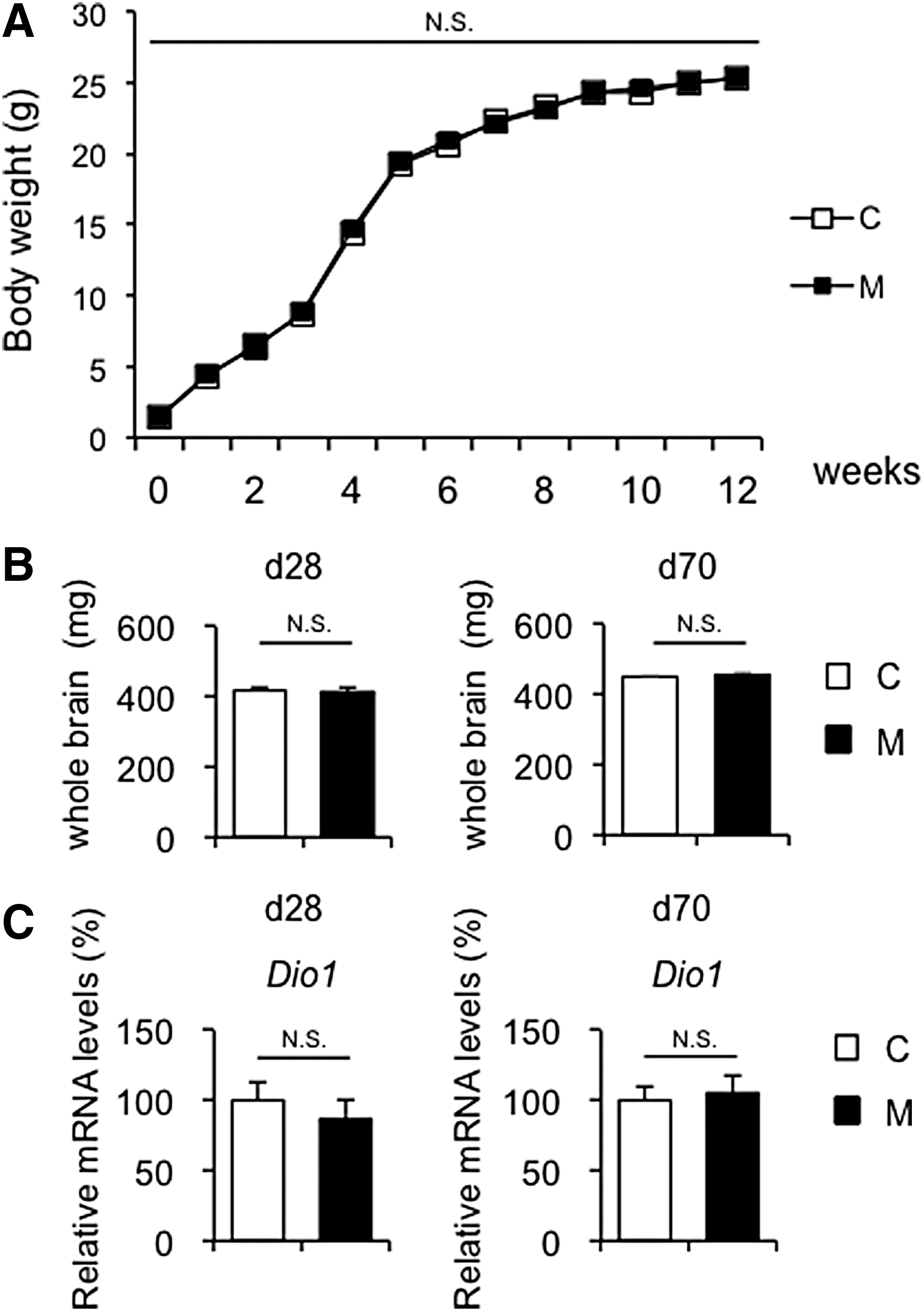
General phenotypes of offspring. (
Serum fT3, fT4, and TSH levels of C and M offspring at day 1 (C, n = 4–5; M, n = 4–5), day 28 (C, n = 3–5; M, n = 3–5), and day 70 (C, n = 3–4; M, n = 3–8). Data are mean ± SE.
p < 0.05; ** p < 0.01.
M, off spring of dams with mild maternal hypothyroxinemia; C, control offspring; fT3, free triiodothyronine; fT4, free thyroxine; TSH, thyrotropin; N/A, not applicable.
Furthermore, type 1 iodothyronine deiodinase (Dio1) mRNA levels were evaluated in the liver, which reflects the peripheral thyroid status in the mouse (28). It was found that hepatic Dio1 mRNA levels were similar between M and C offspring at day 28 and day 70 (Fig. 2C). These results suggest that mild maternal hypothyroxinemia did not affect thyroid hormone action in the offspring.
Mild maternal hypothyroxinemia induced spatial learning impairments in adult offspring
In order to evaluate whether mild maternal hypothyroxinemia causes cognitive and behavioral alterations in offspring, several behavior tests were performed at day 70 in adulthood.
Previous studies showed that maternal hypothyroidism induced hippocampal function impairments in the offspring (29 –32). Accordingly, the Morris water maze test and fear conditioning test were performed, which are commonly used to assess hippocampal memory function in rodents. In the Morris water maze test, latency at the fourth and fifth trials was significantly higher in M offspring than in C offspring, indicating memory dysfunction in M offspring (Fig. 3A).
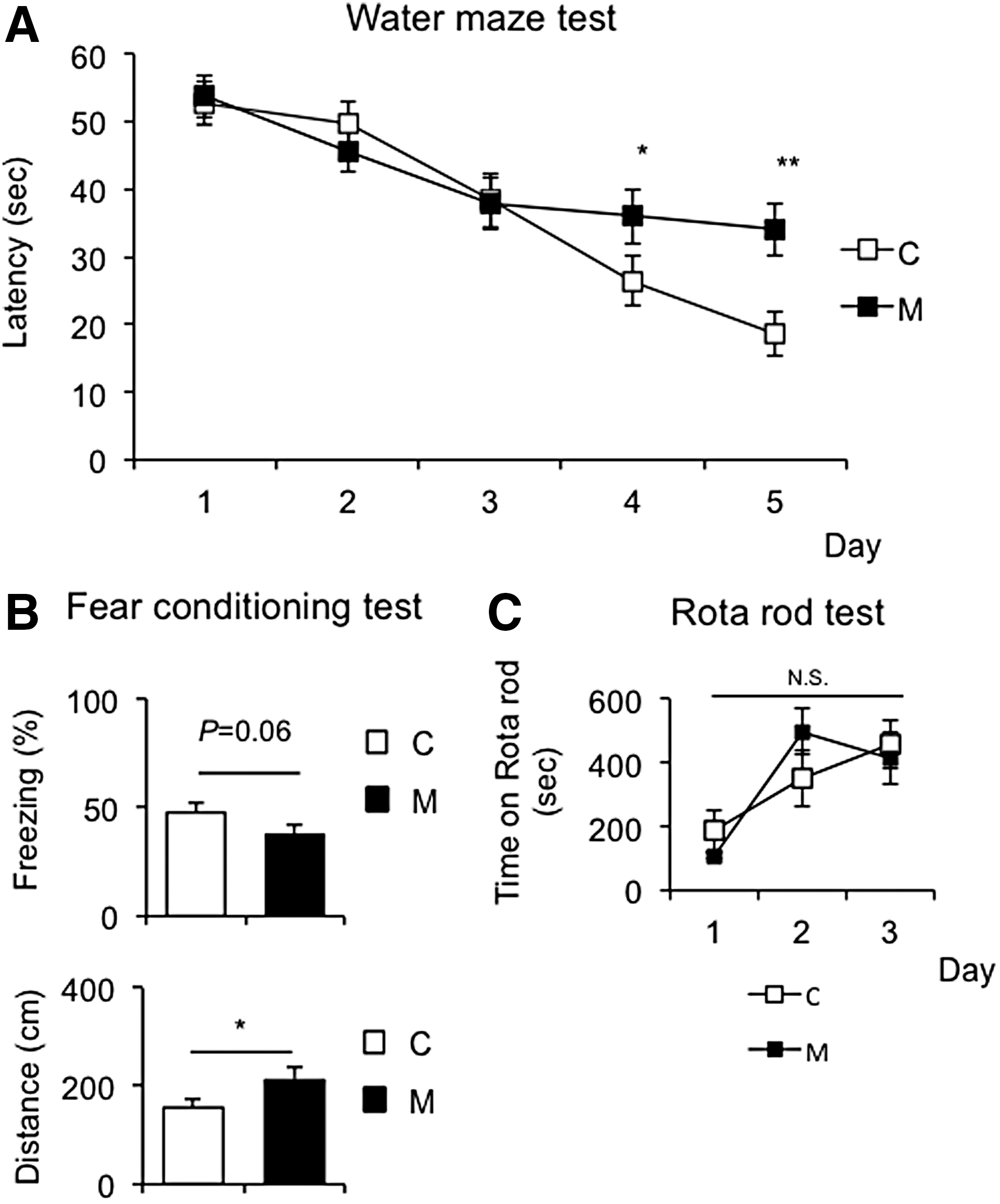
Behavioral analysis of offspring at day 70. Several behavior tests were performed at day 70 (C, n = 8; M, n = 8). (
The fear conditioning test was performed to assess the ability of C57BL6J mice to learn and remember an association between environmental cues and aversive experiences (33,34). The cued fear conditioning test demonstrated that the duration of freezing was slightly shorter in M offspring than in C offspring (Fig. 3B, upper panel) and also that the distance traveled was significantly longer in M offspring than in C offspring (Fig. 3B, lower panel). These results indicate that M offspring have weaker fear memory formation than C offspring.
Since maternal severe hypothyroidism is known to cause morphological abnormalities and functional defects in the cerebellum (35), the rotarod test was employed to evaluate motor coordination. However, no significant differences were found between M and C offspring in the rotarod test, suggesting intact motor function in M offspring (Fig. 3C).
Since perinatal stress is associated with psychiatric disorders, such as anxiety, and schizophrenia (36), the open-field test (Supplementary Fig. S2A), light–dark exploration test (Supplementary Fig. S2B), and the elevated plus maze test (Supplementary Fig. S2C) were employed to evaluate anxiety-related behaviors in M and C offspring. In these behavioral tests, no significant differences were observed between the two groups.
Collectively, these results indicate that M offspring have impaired memory formation in adulthood. Since the hippocampus is responsible for memory formation, mild maternal hypothyroxinemia may induce hippocampal impairments in offspring.
Mild maternal hypothyroxinemia did not induce morphological changes in the hippocampus of offspring
Since M offspring showed impaired hippocampal function based on the results of the performed behavioral tests, morphological changes in the hippocampus of M and C offspring were examined at day 70 after the behavior tests. M offspring exhibited no gross morphological abnormalities in the CA1, CA2, or CA3 pyramidal cell layers (Fig. 4A). A similar localization of Bdnf expression was also found between M and C offspring (Fig. 4B). Moreover, no gross morphological abnormalities or changes in localization of Bdnf expression in the hippocampus of M offspring compared to those of C offspring were found either at day 28 (Supplementary Fig. S3A and B) or at day 70 without the behavior tests (Supplementary Fig. S4A and B). In the hippocampus, Bdnf was observed in the neuronal cell bodies in both the granule and pyramidal cell layers. In the cells, Bdnf was distributed throughout the cytoplasm but was not found in the nucleus at either day 28 or day 70, as previously reported (37,38).

Histological analysis of the hippocampus of offspring at day 70. (
Mild maternal hypothyroxinemia affects hippocampal Bdnf exon IV expression and the DNA methylation status
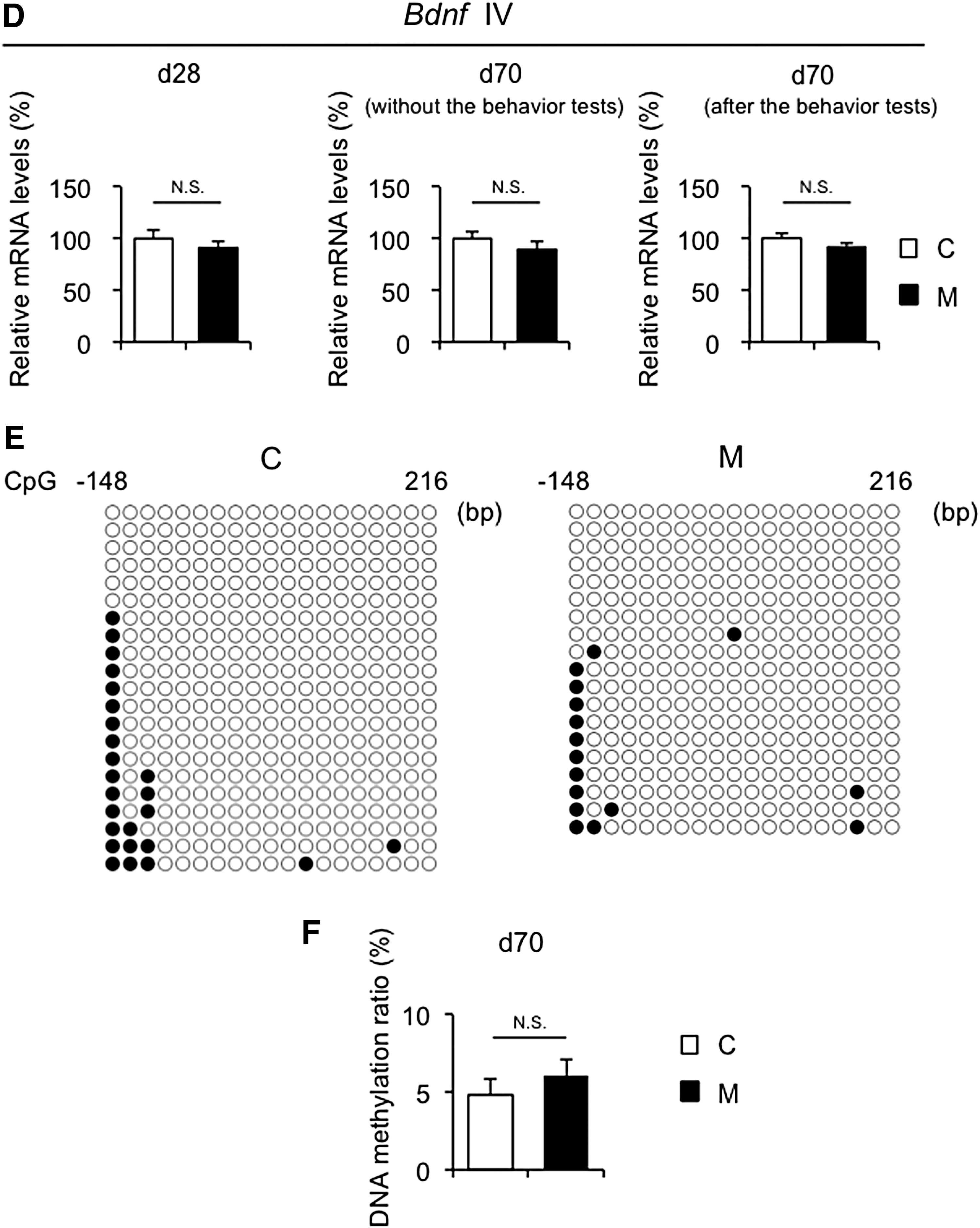
Hippocampal Bdnf exon IV expression is associated with neuronal plasticity and psychopathology and is epigenetically regulated in response to environmental stresses (12). Therefore, hippocampal Bdnf exon IV expression and the DNA methylation status were evaluated at day 28 and day 70 (Fig. 5A). Genes that play important roles in neural activity and development were also examined, such as glutamic acid decarboxylase 1 (Gad1), reelin (Reln), and myelin basic protein (Mbp) (15). At day 28, Bdnf exon IV expression was significantly weaker in M offspring than in C offspring (Fig. 5A, left panel), whereas no significant changes were noted in Gad1, Reln, or Mbp mRNA levels between the two groups (Supplementary Fig. S5A). At day 70, although Bdnf IV expression was similar between the two groups without the behavior tests (Fig. 5A, center panel), it was significantly weaker in M offspring than in C offspring 24 hours after the behavior tests (Fig. 5A, right panel). On the other hand, no changes in Gad1, Reln, or Mbp mRNA levels were found between the two groups at day 70 (Supplementary Fig. S5B–D). In addition, Gad1 and Reln mRNA levels were comparable between M and C offspring without and 24 hours after the behavioral tests at day 70 (Supplementary Fig. S5C and D).
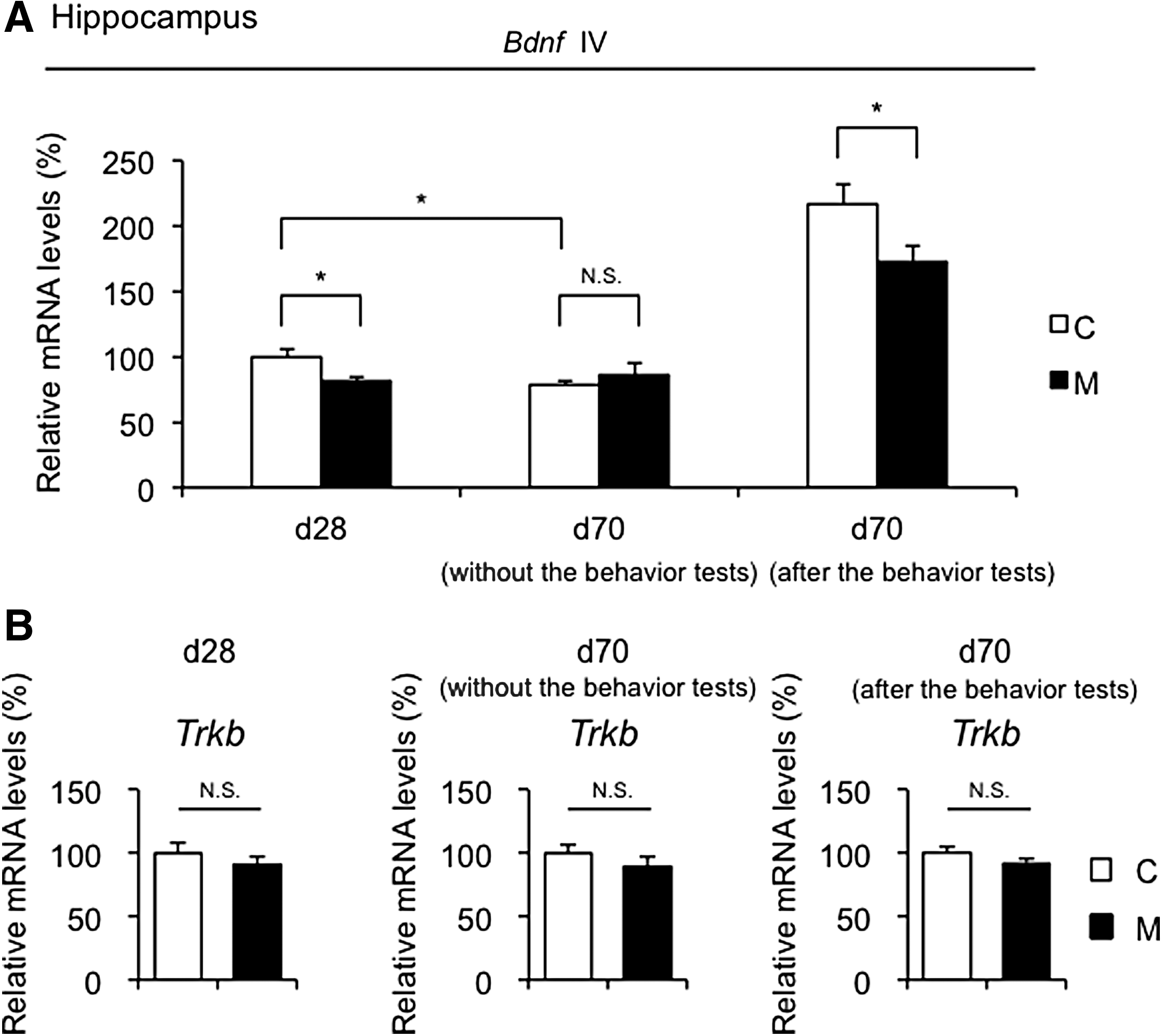
Gene expression analysis of Bdnf exon IV (Bdnf IV) and Trkb in the hippocampus of offspring at day 28 and day 70. Bdnf exon IV (Bdnf IV) (
Recently, it has been reported that exercise induces hippocampal fibronectin type III domain containing 5 (FNDC5) gene expression, which is positively regulated by peroxisome proliferator-activated receptor gamma coactivator 1-alpha (PGC-1α), resulting in an increase in BDNF gene expression in the hippocampus (39). Fndc5 and Pgc1a mRNA levels were significantly increased after the behavioral tests relative to mice without the behavioral tests at day 70 in both M and C offspring (Supplementary Fig. S5E and F), suggesting that the behavioral tests in the current study provided an exercise stimulus, which was sufficient to induce hippocampal Bdnf gene expression.
To confirm Bdnf exon IV expression by qPCR, two different sets of primers were employed (40,41), and similar results were obtained with both sets of primers (Supplementary Fig. S6).
The high affinity receptor for BDNF is tropomyosin receptor kinase B (TrkB). The binding of BDNF to TrkB is required to promote neuronal survival, differentiation, and synaptic plasticity (42,43). No significant differences in Trkb expression were observed between the two groups at day 28 and day 70, even after the behavior tests (Fig. 5B).
We also examined the gene expression of DNA methylation modifiers such as Ten eleven translocations (TETs), DNA methyltransferases (DNMTs), and Methyl-CpG binding protein 2 (MeCP2) at day 28 and day 70. Tet1, Tet2, Tet3, Dnmt1, Dnmt3a, Dnmt3b, and Mecp2 mRNA levels in the hippocampus of the two groups were similar at day 28 and day 70 (Supplementary Fig. S5A and B).
In order to investigate differences in the DNA methylation status of Bdnf exon IV between M and C offspring, the DNA methylation status of 19 CpG sites (i.e., cytosine followed by guanine) was evaluated in the flanking region around the transcriptional start site (Fig. 6A). Bisulfite sequencing revealed that DNA hypermethylation in the promoter region of Bdnf exon IV in the hippocampus was significantly greater in M offspring than in C offspring at day 28 and day 70 (Fig. 6B and C). Moreover, no changes in DNA methylation status in the promoter region of Bdnf exon IV in the hippocampus were found between without and after the behavior tests at day 70 for both C and M offspring (Supplementary Fig. S7). On the other hand, no significant differences in Bdnf exon IV mRNA levels (Fig. 6D) or the DNA methylation status in the promoter region of Bdnf exon IV (Fig. 6E and F) were observed between the two groups in the cerebellum.
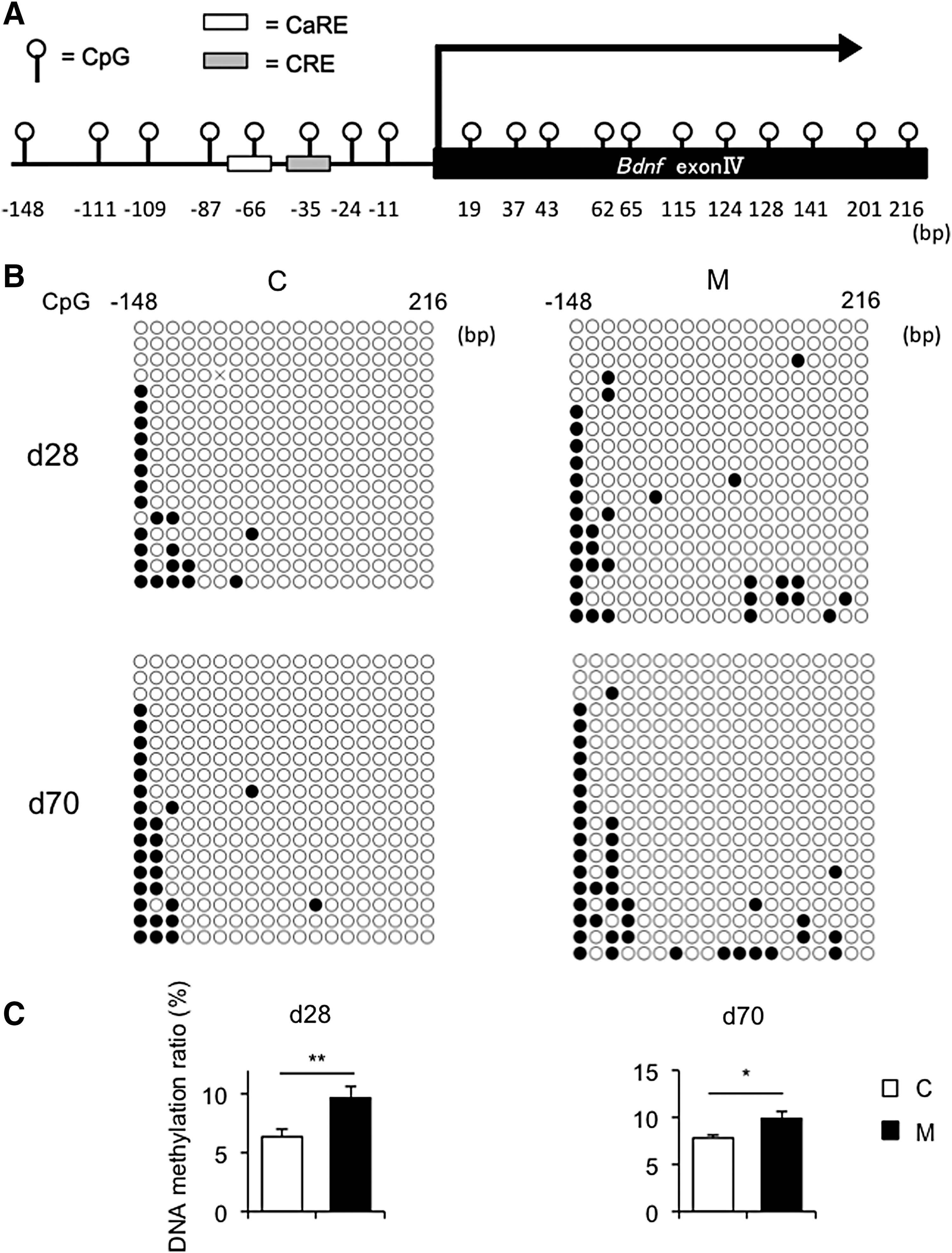
DNA methylation analysis of the promoter region of Bdnf IV in the hippocampus and cerebellum of offspring. (
Since it has been reported that DNA methylation status of Reln and Gad1 in the hippocampus are altered in several psychiatric disorders and that prenatal stress induces DNA hypermethylation of these genes in mice (44,45), the DNA methylation status of these genes was evaluated (Supplementary Fig. S8). There were no differences in DNA methylation status in Reln and Gad1 between C and M offspring. These findings suggest that Bdnf exon IV is more susceptible to maternal hypothyroxinemia in terms of DNA methylation status than Reln or Gad1.
Discussion
The impact of mild maternal hypothyroxinemia, which appears to be more frequent in pregnant women than overt or subclinical hypothyroidism and autoimmune thyroid disease, on perinatal and fetal outcomes remains controversial (46). Therefore, the present study aimed to induce mild maternal hypothyroxinemia in female mice at conception and thereafter during pregnancy by administering MMI, which led to reduced serum fT4 levels without significantly affecting serum fT3 and TSH levels in dams. Considering the previous studies, the degree of maternal thyroid hormone insufficiency depends entirely on the experimental design: the dose of MMI, timing and duration of MMI administration, species (rats or mice), and strain of test animals (47). Therefore, it is critical to optimize the dose of MMI administered to dams and to evaluate thyroid hormone levels in mouse dams in each experiment. Since no studies have ever employed the exact same protocol as that used in the present study, the dose of MMI was optimized in several runs of experiments, and it was concluded that 0.025% (w/v) of MMI was the appropriate dose to induce mild hypothyroxinemia for mouse dams in the current study.
Moreover, maternal hypothyroxinemia induced in the present study did not affect body weight, whole brain weight, or Dio1 expression in the livers of offspring, suggesting a subtle form of thyroid hormone insufficiency.
Increasing evidence supports that maternal hypothyroxinemia can cause prominent and irreversible damage in learning and memory in offspring (32,47 –49). As a region known for its high structural plasticity during development, the hippocampus is involved in learning and memory and is highly sensitive to the actions of thyroid hormone (17,50). The present study, consistent with previous findings, demonstrated that mild maternal hypothyroxinemia during pregnancy induced cognitive disorders without anthropometric changes or hippocampal morphological damage in the offspring in adulthood (6,17,51,52). Since no significant differences were observed between M and C offspring in other behavior tests, such as the open field test, light–dark test, and elevated plus maze test, maternal hypothyroxinemia induced in the present study appears to lead specifically to a deficiency in spatial learning rather than other types of behavioral alterations.
The molecular mechanisms by which maternal hypothyroxinemia cause irreversible or long-term impairments in learning and memory in offspring have not yet been elucidated. The present study demonstrates persistent DNA hypermethylation in the promoter region of Bdnf exon IV, which may cause reductions in hippocampal Bdnf exon IV expression in offspring. Moreover, no significant difference was found in Bdnf exon IV expression or the DNA methylation status in the promoter region of Bdnf exon IV in the cerebellum between M and C offspring. This may be explained by an effect on the development of cerebellum in rodents in the postnatal period, during which serum thyroid hormone levels are normal in offspring derived from dams with mild maternal hypothyroxinemia, and suggests that persistent DNA hypermethylation in the promoter region of Bdnf exon IV is specific to the hippocampus in M offspring (53,54).
Sui and Li demonstrated DNA hypermethylation of the promoter region of Bdnf exon II in the hippocampus in rat offspring derived from dams with severe maternal hypothyroidism (hypothyroid offspring) at day 1 to day 15, when a significant hypothyroid status was observed relative to that in offspring derived from control rat dams (control offspring). Moreover, the DNA methylation status of the promoter region of Bdnf exon II in the hippocampus in hypothyroid offspring was not maintained in adulthood. Furthermore, both the DNA methylation status of the promoter region of Bdnf exon II and Bdnf mRNA levels were comparable between control and hypothyroid offspring at day 60 in adulthood, which may not be a cause of cognitive defects (55). Therefore, it was speculated that persistent DNA hypermethylation in the promoter region of Bdnf exon IV in the hippocampus of M offspring would be Bdnf exon IV specific.
Based on the results obtained in the present study, it is speculated that mild maternal hypothyroxinemia during pregnancy induces persistent DNA hypermethylation in the promoter region of Bdnf exon IV in the hippocampus, and, thus, Bdnf exon IV expression was suppressed in the long term, which may cause irreversible cognitive disorders in the offspring.
One of the limitations of the present study is that it was not possible to analyze expression profiles and the DNA methylation status of genes in specific subnuclei of the hippocampus. Functional differences exist among subnuclei that may be reflected by differences in DNA methylation status of genes. Additionally, the present study did not differentiate between neurons and glia cells. Since Bdnf is expressed in both cell types, elucidating whether mild maternal hypothyroxinemia-induced DNA hypermethylation in the promoter region of Bdnf exon IV occurs in neurons or glia or in both of these cell types would provide insights into the potential functional impact of alterations in the DNA methylation status.
Furthermore, the mechanisms by which mild maternal hypothyroxinemia induces DNA hypermethylation in the promoter region of Bdnf exon IV currently remain unclear. Therefore, more direct evidence of DNA hypermethylation in the promoter region of Bdnf exon IV in the hippocampus of offspring via developmental hypothyroxinemia is needed in the future. Previous studies suggested that DNA demethylation of the promoter region of Bdnf exon IV contributes to Bdnf expression in neurons, and MeCP2 and TETs may induce DNA methylation and demethylation, respectively (56 –58). The present study found no significant difference in the gene expression of MeCP2, TETs, and DNMTs at day 28 or day 70. Evaluations of the recruitment of these epigenetic modifiers to the promoter region of Bdnf exon IV will contribute to the understanding of the underlying molecular mechanisms.
It has been reported that the differences in DNA methylation status would not reflect the differences in steady-state gene expression, especially when the transcriptional activity of the gene is low (59,60). However, in a transcriptionally active status, even subtle differences in DNA methylation status would induce substantial differences in gene expression (59 –61). Since Bdnf exon IV expression in the hippocampus strongly increases from day 14 to day 21 and then decreases throughout adulthood in rodents (12,62), hippocampal Bdnf exon IV expression is more active at day 28 than at day 70, suggesting that it is transcriptionally active for hippocampal Bdnf exon IV at day 28. Therefore, Bdnf exon IV mRNA levels in the steady state were significantly lower in M offspring than in C offspring at day 28, which may be attributed to the DNA hypermethylation status in the promoter region of Bdnf exon IV in M offspring. On the other hand, even though the DNA methylation ratio in the promoter region of Bdnf exon IV was significantly higher in M offspring than in C offspring, since it is transcriptionally less active at day 70 than day 28, no significant differences were observed in Bdnf exon IV mRNA levels in the steady state, which is without behavior tests at day 70. Several studies have demonstrated that hippocampal Bdnf expression increases after exercise and the behavior tests, such as the Morris water maze test and fear conditioning test (63 –66). Thus, as shown in Figure 5A, it is important to note that Bdnf exon IV mRNA levels were significantly higher in C offspring than in M offspring after the behavior tests, which would activate the transcription of Bdnf exon IV. Collectively, these results indicate that the DNA methylation status in the promoter region of Bdnf exon IV reflects basal Bdnf exon IV mRNA levels in early postnatal days, during which transcriptional activity is high, but may also reflect the magnitude of Bdnf exon IV expression in response to environmental cues such as exercise and behavior tests in adulthood, during which basal gene expression is low.
A previous study reported that exon IV-specific Bdnf expression correlated with the degree of DNA methylation of the corresponding exon IV-associated CpGs in rats exposed to associative fear conditioning (65). Moreover, in the present study, Bdnf exon IV mRNA levels in the cerebellum were similar between M and C offspring, even after the behavior tests, and this may be attributed to the absence of a significant difference in the DNA methylation status in the promoter region.
Similar findings have been reported. A transient surge in hepatocyte type 2 deiodinase (DIO2), which activates T4 to T3 conversion during the perinatal period, changes the DNA methylation status of lipid metabolism-related genes, which persist in adulthood. The DNA methylation status established by DIO2 in early life has been shown to affect the expression of lipid metabolism-related genes in response to a high fat diet in later life (67). Similar to these findings, the present results suggest that the DNA methylation status of Bdnf exon IV in the hippocampus is susceptible to developmental hypothyroxinemia and that the DNA hypermethylation status established during the fetal period is relatively stable and persists into adulthood as epigenetic memory, which may affect Bdnf exon IV expression in response to environmental cues.
However, the direct causal relationship between DNA methylation and the mRNA expression of Bdnf exon IV in the hippocampus remains unclear. Therefore, further investigations employing the targeting of increased or decreased DNA methylation in the promoter region of Bdnf exon IV in the hippocampus during the early postnatal period are needed in order to elucidate the causal pathways.
Nevertheless, the present study demonstrates for the first time that mild maternal hypothyroxinemia induces persistent DNA hypermethylation in the promoter region of Bdnf exon IV in offspring as epigenetic memory, which may result in long-term cognitive disorders.
Footnotes
Acknowledgments
This work was supported in part by Grants-in-Aid for Scientific Research (KAKENHI) from the Japan Society for the Promotion of Science; Grant Number JP 26461376 (K.H.).
Author Disclosure Statement
The authors have nothing to disclose.