
Abstract
Background:
MicroRNAs (miRNAs) play a unique role in posttranscriptional regulation of gene expression and control different aspects of skin development, homeostasis, and disease. Although it is generally accepted that thyroid hormone signaling is important in skin pathophysiology, the role of their nuclear receptors (TRs) in cutaneous miRNA expression has yet to be explored.
Methods:
RNAseq was used to compare the skin miRnome of wild-type mice and genetically modified mice lacking both TRα1 and TRβ, the main thyroid hormone binding isoforms. Changes in miRNAs with a crucial role in skin physiopathology were confirmed by stem-loop quantitative polymerase chain reaction in both total skin and isolated keratinocytes, and the levels of their target mRNAs were evaluated by real-time polymerase chain reaction.
Results:
The skin of TRα1/TRβ knockout mice displays altered levels of >50 miRNAs. Among the downregulated species are several miRNAs, including miR-21, miR-31, miR-34, and miR-203, with crucial roles in skin homeostasis. TRα1 appears to be the main isoform responsible for their regulation. Increased levels of gene transcripts previously shown to be bona fide targets of these miRNAs are also found in the skin and keratinocytes of TR-deficient mice. This suggests that multiple miRNAs that are downregulated in the absence of TRs cooperate to regulate gene expression in the skin.
Conclusions:
The miRNAs reduced in TRα1/TRβ knockout mice are known to play crucial roles in epidermal proliferation, hair cycling, wound healing, stem-cell function, and tumor development, all processes altered in the absence of TRs. These results suggest that their regulation could contribute to the skin defects found in these mice and to the skin disorders associated with altered thyroid status in humans.
Introduction
M
The actions of the thyroid hormones are mediated by binding to nuclear receptors (TRs), which act as ligand-dependent transcription factors. TRs are encoded by two distinct genes, and TRα1, TRβ1, and TRβ2 are the main thyroid hormone binding isoforms (12,13). Observations both in patients and in experimental animal models show that the skin is an important target for thyroid hormones (13 –16). Thyroid hormones can induce epidermal thickening (17,18), and it has previously been shown that genetically modified mice lacking TRα1 and TRβ (TR KO mice) display reduced keratinocyte proliferation and decreased hyperplasia in response to topical application of 12-O-tetradecanolyphorbol-13-acetate (TPA) or retinoids (19,20). These effects suggested an altered susceptibility of TR-deficient mice to develop skin tumors. Indeed, these animals develop fewer tumors than normal mice when exposed to a chemical carcinogenesis protocol (21). Interestingly, the thyroid hormones appear to play a repressive role in basal-cell carcinoma (BCC) development (22,23), and depletion of deiodinase 3 (DIO3), a thyroid hormone-inactivating enzyme, strongly reduces BCC formation by a mechanism that involves the control of miR-21 oncogenic action (24).
Furthermore, in agreement with the frequent alopecia found in thyroid hormone–deficient patients (25 –27), TR KO mice display impaired hair cycling associated with a decrease in follicular-cell proliferation and develop alopecia after serial depilation (28). The thyroid hormones also play a positive role in wound healing (29 –31), and TR KO mice present a wound-healing defect, with retarded re-epithelialization, associated with impaired proliferation. Taken together, these results show that binding to their nuclear receptors is required for the thyroid hormone effects in the skin. Furthermore, TRα1 and TRβ1 expression has been observed in the skin. Both TRα1 and TRβ1 receptors appear to contribute to skin homeostasis (19,21,28). Since TRβ2 has a highly restricted expression pattern (12,13), TRβ1 is the expressed isoform in skin.
Stem cells located at the bulge of the hair follicles are involved in forming not only the follicle but also the epidermis and contribute to the re-epithelialization after wounding and to skin tumor development (32 –34). Thus, the skin phenotype of TR KO mice was compatible with a defect in the number or function of the bulge stem cells. Indeed, in TR KO mice, bulge stem cells present a clear defect in their mobilization (exit of their quiescent state and migration out of the niche) (35). This is associated with increased activation of SMAD signaling, which can lock follicle stem cells in a quiescent state (36 –38).
This work analyzed the pattern of miRNA expression in the skin of TR-deficient mice. It was found that TRs are required for full expression of an important number of miRNAs. Among them, miRNAs with important roles in skin homeostasis, including miR-21, miR-31, miR-34, and miR-203, were significantly reduced in the absence of TRs. Epidermal keratinocytes from TR KO mice showed a similar decrease in miRNA expression as total skin, demonstrating that these cells are the primary targets of the receptors for the regulation of miRNA abundance. This decrease was observed in both sexes, independently of the hair cycle stage and was accompanied by increased expression of target mRNAs. Furthermore, the reduction in miRNA levels was much less marked in the skin of TRβ KO mice than in TRα1 KO mice, suggesting a predominant role of TRα1 in their regulation.
Methods
Animals
All animal studies were done in agreement with the European Community Law (86/609/EEC) and the Spanish law (R.D. 1201/2005), with approval of the Ethics Committee of the Consejo Superior de Investigaciones Científicas. TRα–/–/TRβ–/– double KO mice have a deletion of TRα1 but not TRα2 and lack all TRβ isoforms (39). These animals, as well as single TRα1–/–/TRβ+/+ mice (KOα1), TRα1+/+/TRβ–/– mice (KOβ), and wild-type (Wt) TRα1+/+/TRβ+/+ animals with the same genetic background were genotyped (39) and used for the studies. Experiments were performed with dorsal skins of adult male and female mice. Age-matched animals were always used in the experiments. Dorsal skins were shaved and depilated 24 hours before treatment with TPA (Sigma–Aldrich, St. Louis, MO). TPA (12 μg) was applied topically twice (on days 1 and 3), and mice were sacrificed on day 4. In the control group, animals were treated with vehicle (acetone) only.
Histological analysis
Dorsal skin samples were fixed with paraformaldehyde and embedded in paraffin. Skin sections (4 μm) were stained with hematoxylin and eosin to determine the stage of the hair cycle. The anagen phase is the active growth phase in which bulb hair penetrates deep into the dermis. This is followed by a regression phase or catagen, in which the hair follicle moves upwards to the epidermal surface, and the telogen phase is a resting phase, in which the hairs are shorter reaching only the dermal fat. The anagen and not-anagen phases of the hair cycle were visually recognized by histological morphology and penetration of hairs into the dermis.
miRNA analysis by RNAseq
Total RNA was obtained from dorsal skin of nine Wt and nine TRα1/TRβ KO adult male mice treated with acetone and from the same number of animals treated with TPA using a miRNeasy Mini kit (cat. no. 217004; Qiagen, Valencia, CA) following the manufacturer's protocol. Samples from three animals from each group were pooled for analysis. Sample RNA integrity numbers were analyzed on an Agilent 2100 Bioanalyzer (Agilent, Santa Clara, CA). RNA samples were processed as described in the “TruSeq Small RNA Sample Preparation Guide” (Illumina part # 15004197 Rev. A of November 2010). Briefly, 1 μg of purified total RNA containing the small RNA fraction was sequentially ligated to 3′ and 5′ adapters, with the truncated form of T4 RNA ligase 2 and the T4 RNA ligase, respectively. Reverse transcription (RT) with SuperScript II Reverse Transcriptase followed, yielding cDNA adapter-ligated libraries that were amplified by polymerase chain reaction (PCR) with Phusion DNA Polymerase and Illumina RNA PCR primers. cDNA amplified libraries were pooled and separated by polyacrylamide gel electrophoresis, and a fraction of 145–160 bases was extracted. The purified fraction constituted the multiplexed purified libraries that were applied to an Illumina flow cell for cluster generation, and sequenced on the Illumina Genome Analyzer IIx with SBS TruSeq v5 reagents following the manufacturer's protocols. Image analysis and per-cycle base calling was performed with Illumina Real Time Analysis software (RTA1.9; Illumina, San Diego, CA). Conversion to FASTQ read format was performed by CASAVA-1.8 (Illumina). Then, sequenced reads were analyzed using the bioconductor package “oneChannelGUI.” Briefly, Bowtie was used for primary mapping, resulting in a SAM file subsequently converted in a BAM file using Picard Java tools. Reads were aligned to mouse genome assembly (GRCm38/mm10 mouse genome). Then, the 5′ and 3′ end linkers were removed by trimming. Quality control was performed, and those samples with low quality were removed for further analyses. Finally, differential expression was characterized using edgeR and baySeq.
Quantitative real-time PCR assays of miRNAs and mRNAs
miRNAs were quantified in triplicate by the stem-loop real-time PCR using a TaqMan MicroRNA RT kit followed by the TaqMan Universal PCR Master Mix No AmpERASE (Applied Biosystems, Foster City, CA). The primers purchased from Applied Biosystems are listed in the Supplementary Material (Supplementary Data are available online at
Western blotting
For preparation of whole-cell extracts, dorsal skins were grounded with a mortar in liquid nitrogen and homogenized in lysis buffer composed of 200 mM of Hepes, pH 7.9, 25% glycerol, 400 mM of NaCl, 1 mM of EDTA, 1 mM of EGTA, 1.25 mM of DTT, 100 mM of PiPNa, 200 mM of sodium orthovanadate, 500 mM of NaF, 0.1 mM of PMSF, 10 mg/mL of aprotinin, and 10 mg/mL of leupeptin, NP-40 1%. The amount of protein was measured by the Bradford method. Thirty-fifty micrograms of protein was boiled in Laemmli buffer, separated on 12% polyacrylamide gels, and then transferred to Amersham™ nitrocellulose blotting membranes. Blots were blocked with 4% bovine serum albumin milk in TBS/0.5% Tween 20, and incubated with the antibodies indicated in the Supplementary Material at 4°C overnight. After washing, membranes were incubated with a peroxidase-conjugated secondary antibody for two hours at room temperature and analyzed using the enhanced chemiluminescence method (Immun-Star HRP; Bio-Rad Laboratories), according to the manufacturer's recommendations.
Isolation of keratinocytes
Keratinocytes were derived from the total skin of four-day-old Wt controls and double TRα1/TRβ1 KO mice. Tails from each pup were genotyped, the skin was removed, and the epidermis was obtained by treatment overnight with 0.25% trypsin at 4°C.
Cell culture and transfections
The mouse keratinocyte cell line MCA3D was grown in HAM's F12 medium containing 10% fetal calf serum. Before the experiments, cells were shifted to a medium with thyroid hormone–depleted serum by treatment with resin AG-1-X8 (Bio-Rad) and were transfected in 24-well plates with 1 μg of an empty vector (pSG5-0) or with TRα1 or TRβ1 cloned in the same vector using Lipofectamine 2000. Cells were incubated with or without triiodothyronine (T3; 5 nM) for 36 h, and miRNAs were isolated and quantitated as described above.
Statistical analysis
Differences with respect to Wt animals were assessed using Student's t-test when two groups were compared or by analysis of variance when more than two groups were compared, with statistical significance set at p < 0.05.
Results
Deregulated miRNA expression in the skin of TR-deficient mice
miRNAs have been shown to control skin homeostasis (10). To investigate whether TR deletion could alter the skin miRNA profile, miRNA-Seq was conducted of the dorsal skin from Wt and genetically modified male mice lacking TRα1 and TRβ (TR KO) and therefore lacking the main thyroid hormone binding isoforms. The analysis showed that irrespectively of the phase of the hair cycle, 53 miRNAs showed a differential expression (p < 0.05) between normal skin and the skin of TR KO mice, with 20 miRNAs being upregulated and 33 downregulated in the KO animals (Supplementary Table S1). These differences were more marked when only animals that were not in the anagen phase were considered, finding 82 miRNAs (56 downregulated and 26 upregulated) with statistically significant differences (adjusted p < 0.05; Supplementary Table S2). Interestingly, among the miRNAs reduced in the skin of TR-deficient mice, species of the miR-21, miR-31, and miR-34 families, with crucial roles in skin pathophysiology, were identified (40 –49).
Since an altered response to TPA in TR-deficient mice has previously been observed (19,28,35), the miRNome of Wt and TR KO mice was also compared after TPA treatment. This treatment resulted in 61 differentially expressed miRNA species in the Wt animals (44 downregulated and 17 upregulated; p < 0.05; Supplementary Table S3), while 84 miRNAs were affected by TPA in the animals lacking TRs (adjusted p < 0.05; Supplementary Table S4). Analysis of differences between Wt and TR KO mice after TPA treatment revealed the existence of 30 miRNAs with differential expression (p < 0.05) in normal skin and in the skin of animals lacking TRs. Of these miRNAs, 12 were increased and 18 were decreased in the absence of TRs (Supplementary Table S5). MiR-31 and miR-34 were again among the reduced miRNAs after TPA treatment in TR KO mice versus the Wt mice.
Reduced expression of key miRNAs in the skin of TRα1/TRβ KO mice
As the miRNome analysis showed that miRNAs with well-known functions in the skin were among the deregulated species in TR-deficient mice, attention was centered on miR-21, miR-31, and miR-34 that were downregulated in the absence of the receptors. In addition, the levels of Let-7, with important roles in development, stem cells, and cancer (50), were evaluated. These showed an apparent increase in the TRα1/TRβ-deficient animals. miR-203, with an important role in stemness and epidermal differentiation, was also analyzed (6,51). To confirm the results of the miRNA-Seq analysis further, the levels of the selected miRNAs were quantitated by RT-qPCR in the skin of a different group of normal and KO male mice. Confirming the data obtained by sequencing, the levels of miR-21, and less strongly of miR-31, were reduced in the skin of TR null mice. The miR-34 family comprises three members: miR-34A, miR-34B, and miR-34C. The latter two are generated from a single bi-cistronic transcript (40), and their levels were also significantly lower in the KO animals. Although differences in miR-203 expression were not detected by sequencing, there was a statistically significant reduction in this miRNA. In addition, even though the levels of Let7b were slightly elevated in the KO mice, the difference was not statistically significant (Fig. 1). The comparison of Ct values for the individual miRNAs with those for the U6 RNA, used as an internal control, confirmed that miR-203, Let7, and miR-21 were highly abundant in the skin, whereas the numbers for the miR-31 and miR-34A were intermediate, and miR-34B and miR-34C showed a low expression (Supplementary Table S6).
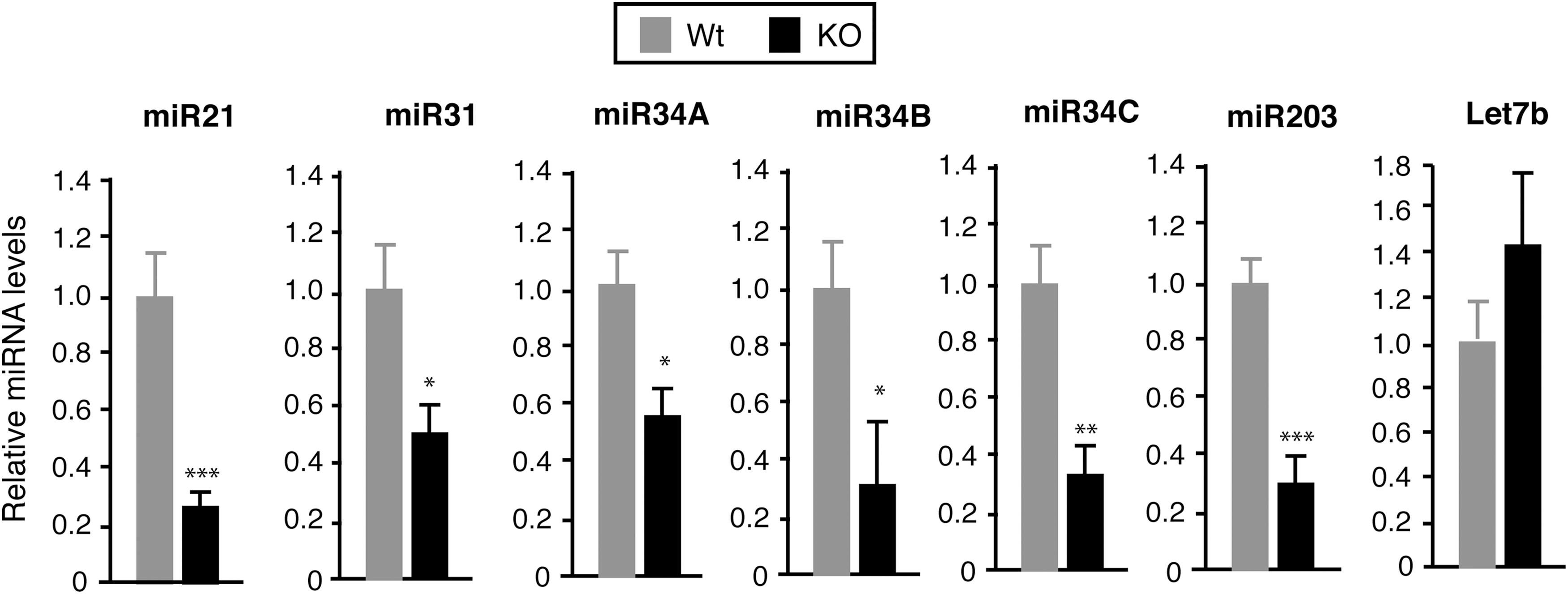
Altered microRNA (miRNA) expression in the skin of nuclear receptors (TR)-deficient mice. The levels of the indicated miRNAs were analyzed using quantitative real-time polymerase chain reaction (PCR) in the dorsal skin of wild-type (Wt) and double knockout (KO) adult male mice (n = 8), lacking TRα1 and TRβ. miRNA expression was normalized to U6 RNA. Results are expressed relative to the amount of each miRNA in Wt animals. Data are shown as mean values ± standard error (SE), and asterisks denote statistically significant differences relative to Wt mice. *p < 0.05; **p < 0.01; ***p < 0.001.
In order to analyze whether the absence of TRs could affect the pattern of cutaneous miRNA expression in a sexually dimorphic manner, the expression of the same miRNAs was next examined in control and TRα1/TRβ-deficient female mice. The results obtained were strikingly similar to those observed in males (Supplementary Fig. S1), showing the lack of sexual dimorphism and corroborating again the role of TRs as regulators of the expression of miRNAs with a key role in skin homeostasis.
As the pattern of miRNA expression can vary depending of the stage of the hair cycle (10,41) and TRs can affect hair growth (28), next the levels of the selected miRNAs were compared in male mice in the anagen phase and not-anagen phase, defined by visual inspection of the penetration of the hair bulbs into the dermis. Figure 2 shows that in Wt mice, the levels of miRNAs were generally lower when animals were in the anagen phase. This was not observed in TR KO mice, and as a consequence, differences with respect to the Wt animals in the levels of miR-21, miR-31, and miR-34 were attenuated when mice were in the anagen phase. In addition, the levels of Let7b were again unchanged. Furthermore, no differences in miR-31 and miR-34A levels between Wt and TR null female mice were found in the anagen phase, while miR-21, miR-34B, and miR-34C were strongly reduced at this stage of the hair cycle (Supplementary Fig. S2), unmasking some dissimilarities in the regulation of miRNA expression by TR depending on sex.
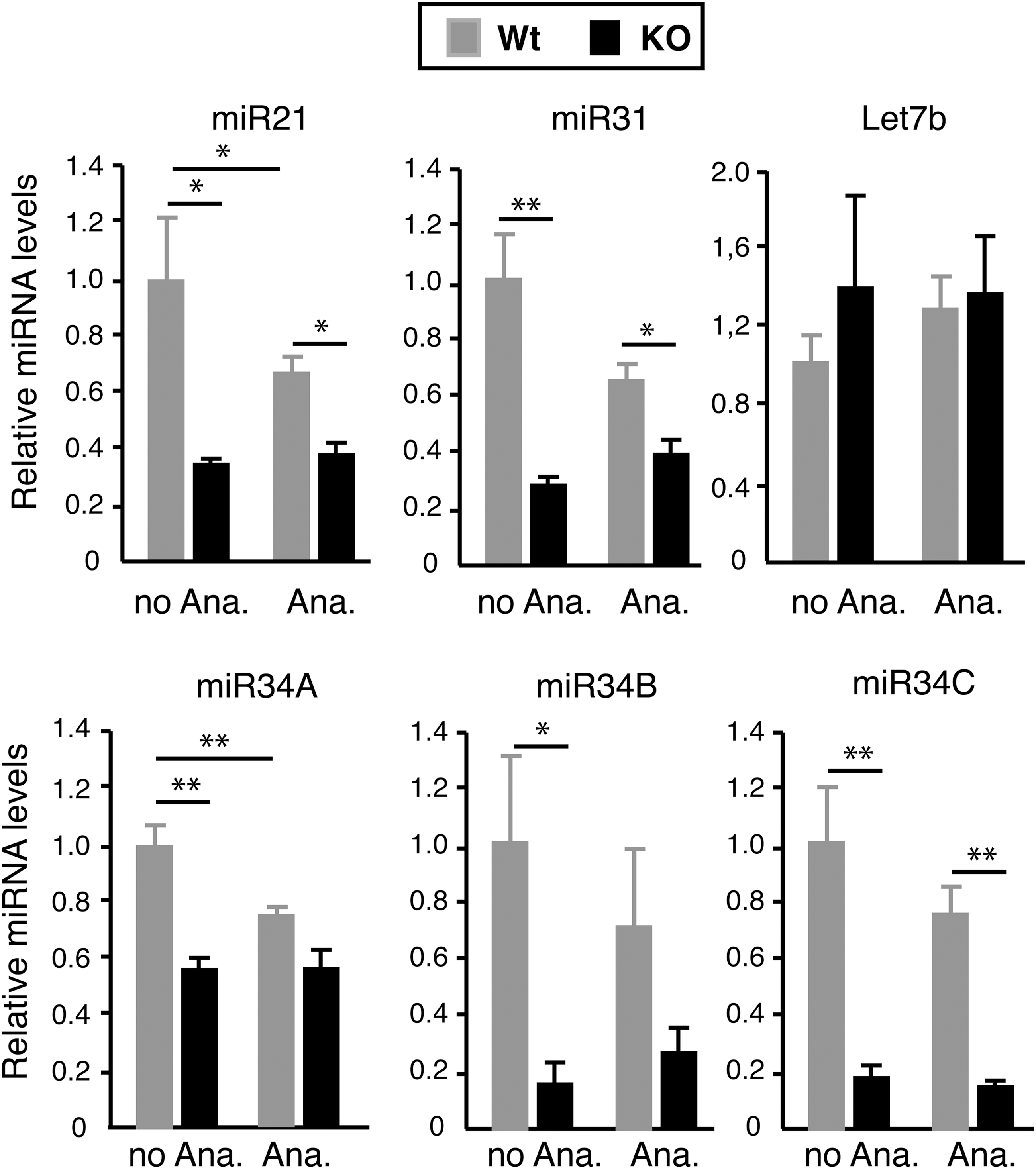
miRNA expression is reduced in the skin of KO mice in the anagen phase and not-anagen phase. The levels of miRNAs were quantitated in the dorsal skin of Wt (n = 6) and TRα1/TRβ KO male mice (n = 7) in the anagen phase (Ana.) and in other phases of the hair cycle (no Ana.). Results are expressed relative to those obtained in Wt mice that are not in the anagen phase. Data are means ± SE. Statistically significant differences between the anagen phase and not-anagen phase and Wt and KO mice are indicated with asterisks (*p < 0.05; **p < 0.01).
Altered miRNA response to TPA in mice lacking TRα1 and TRβ
The changes in abundance of the selected miRNAs obtained by RNA-seq in TPA-treated animals were also confirmed by RT-qPCR. Figure 3 shows the results obtained in male mice. TPA treatment of Wt mice resulted in a decrease of miR-34A, miR-34C, miR-203, and Let7b, and no statistically significant changes in the levels of miR-21, miR-31, and miR-34B. This pattern was different in TR-deficient mice where, with the exception of Let-7, the response to TPA was in general less marked and, as a consequence of the reduced expression of miR-21, miR-31, miR-203, and miR-34A with respect to the Wt mice observed in the untreated animals, was no longer found after TPA treatment. These results confirm the smaller number of altered miRNAs in TPA-treated TR null with respect to the normal mice obtained by RNA-seq. On the other hand, similar results were obtained when female mice were subjected to the same treatment. In this case, the phenotype was even more pronounced, since all miRNAs that were reduced in the untreated KO mice were not further decreased or were even increased after TPA treatment (Supplementary Fig. S3). TPA treatment induces anagens. Therefore, it is possible that regulation by TPA might simply reflect this change. However, its effects appear to be specific, since the miRNA profile obtained was different from that found in the untreated mice in the anagen phase, and differences between Wt and TR KO mice in the anagen phase were still observed (see Fig. 3 and Supplementary Fig. S2).
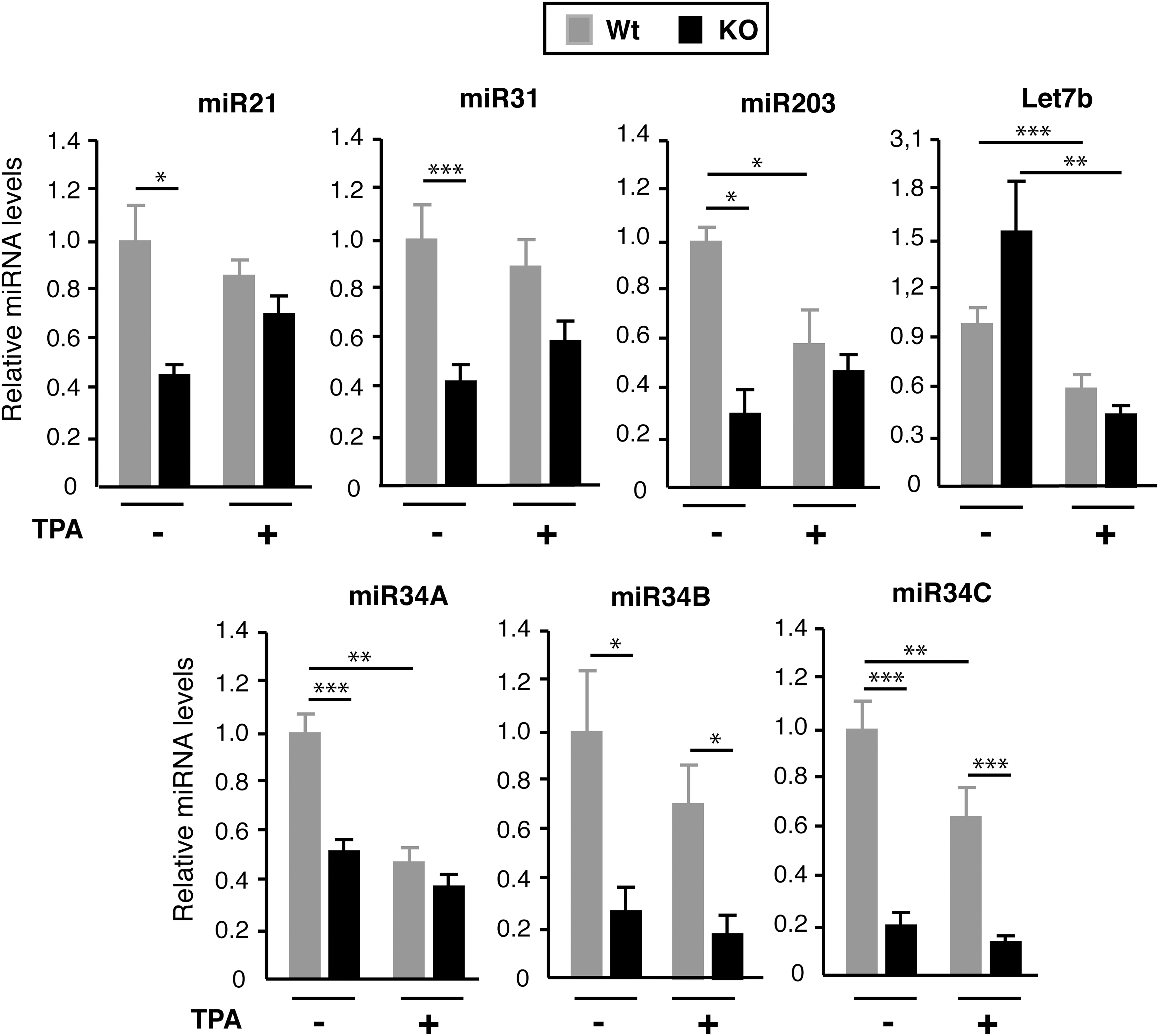
TR KO mice show a reduced miRNA response to 12-O-tetradecanolyphorbol-13-acetate (TPA) administration. Wt and TRα1/TRβ KO mice were sacrificed after topical treatment of the dorsal skin with vehicle (acetone; n = 13) or TPA (n = 9) for five days. miRNA expression was analyzed, and results are expressed relative to those obtained for each miRNA in the skin of Wt mice without TPA treatment. Data are means ± SE, and asterisks denote statistically significant differences between the indicated groups (*p < 0.05; **p < 0.01; ***p < 0.001).
Upregulation of miRNA targets upon TRα1/TRβ deletion
Since microRNAs predominantly act to decrease target mRNA levels (4), next the study analyzed whether the observed reduction in miRNA expression in the skin of TR null mice was reflected in an opposite enhancement in the levels of their target mRNAs. With this aim, transcript levels of Pten, Timp3, and Spry2, which have been shown to be miR-21 targets both in the epidermis and fibroblasts (28,42,45,48,52), were measured. As miR-31 targets, the mRNA levels were examined for Fgf10 and for the components of Wnt and BMP signaling pathways Bambi, and Dlx3, which have been demonstrated to be direct targets of this small RNA in the skin (41). The cell-cycle regulator Cdk4, as well as Notch 1 and Wnt1, were also used as well-established targets of the miR-34 family (53,54) and Tap 63 and Dp63 as miR-203 targets, since regulation of p63 by miR-203 is also direct and mediated by a specific 3′-UTR target site (7,51). As shown in Figure 4, the levels of most of these miRNAs were higher in the skin of the double TR KO than in Wt mice, suggesting that the observed changes in miRNA expression could alter expression of genes involved in the regulation of key signaling pathways involved in skin proliferation and differentiation.

Effect of TR deletion on expression of target mRNAs. Transcript levels of bona fide targets of the analyzed miRNAs were evaluated by quantitative PCR in the dorsal skin of Wt and double TRα1/TRβ KO male mice (n = 9). mRNA expression was normalized to GusB mRNA. Data are expressed relative to the amount of each mRNA in Wt mice. Data are shown as mean values ± SE. Asterisks denote statistically significant differences between Wt and KO mice (*p < 0.05; **p < 0.01; ***p < 0.001).
Genes encoding miRNAs in the miR-34 family are direct transcriptional targets of p53 (55,56), and miRNA-203 expression in keratinocytes is also directly dependent on the regulation of p53 levels (57). Therefore, it was possible that increased p53 activity could underlie the reduction in these miRNAs in the skin of TR KO mice. In addition, p53 signaling is affected by Hairless (Hr) (58), a TR target gene with important effects on the skin and hair (59), which is reduced in KO mice (Fig. 5B). However, p53 activation was similar in Wt and TRα1/TRβ KO mice (Fig. 5A), suggesting that p53 does not play an important role in miRNA regulation by the receptors. On the other hand, in epidermal keratinocytes, miR-21 is strongly downregulated by BMP4, which induces Smad activation (42), and it has been previously shown that Smad1,5,8 is aberrantly activated in the skin of TR-deficient mice (35). This, as well as the finding that miR-21 target genes such as Pten or Timp3 are upregulated in these animals, suggested that increased BMP4 expression could be involved in miR-21 repression. Indeed, Bmp4 mRNA levels were significantly increased in the skin of TR-deficient mice (Fig. 5B). Upregulation of Spry and Pten in TR KO mice where miR-21 levels are low should lead to reduced activation of the Erk/MAPK and PI3K/Akt axis that play a key role in skin homeostasis and carcinogenesis (45,52). However, these pathways can be stimulated by growth factors, among them Fgf10, which is upregulated in these animals. As shown in Figure 5C, and confirming previous observations (19), Erk and Akt phosphorylation were enhanced in the skin of TR null mice, suggesting a predominant role of stimulatory over inhibitory factors of the activity of these pathways in the absence of TRs.
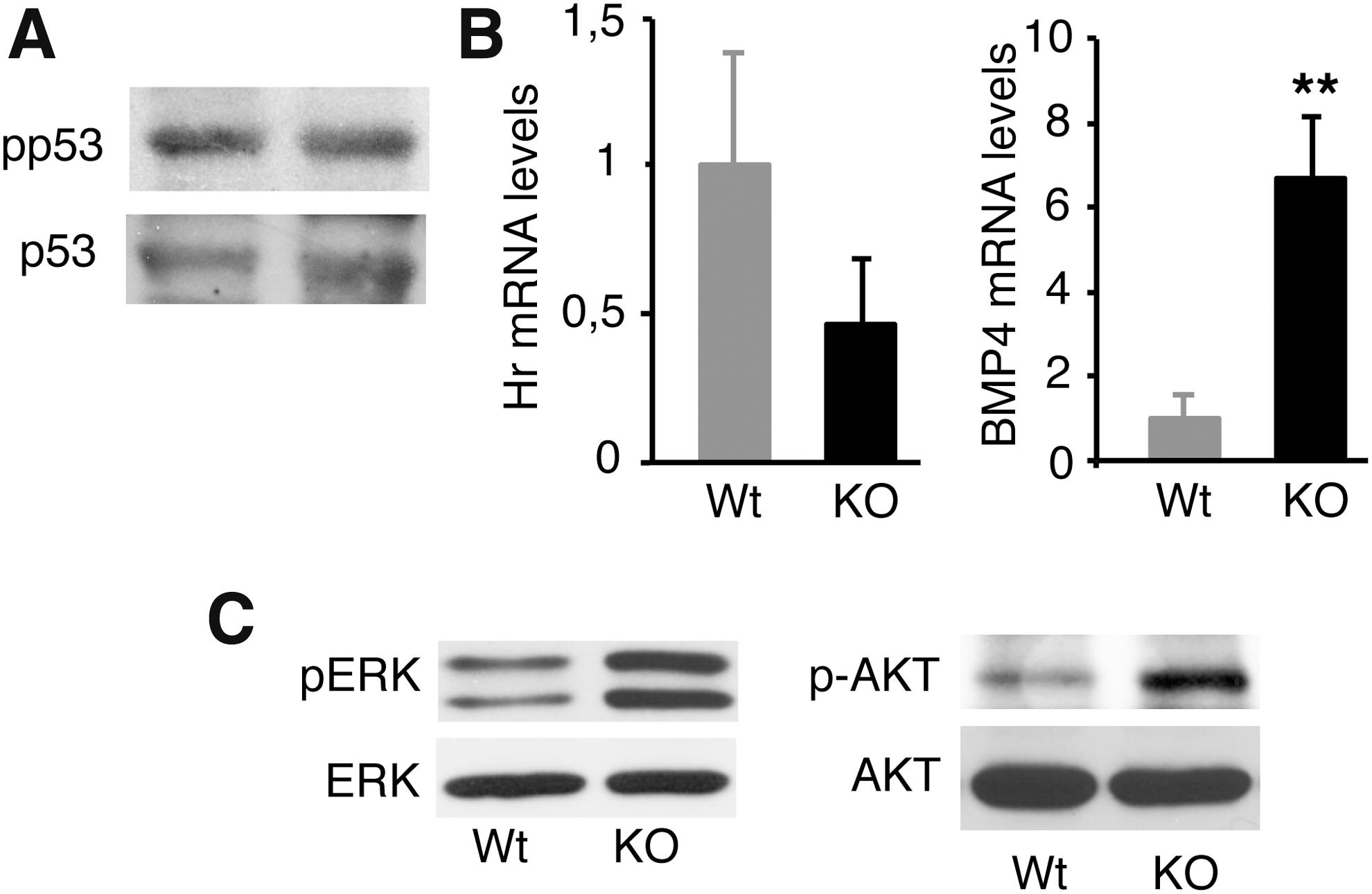
Altered signaling pathways in the skin of TR-deficient mice. (
Effect of TRα1/TRβ deletion on miRNA expression in epidermal keratinocytes
As the miRNA expression profile could differ in the different skin components (5,60), next the study analyzed whether the miRNA regulation observed in the dorsal skin of TR-deficient mice was also observed in keratinocytes. For this purpose, the levels of miRNAs were quantitated in keratinocytes isolated from the total skin of newborn Wt and TRα1/TRβ KO mice. Confirming previous observations (5,40), miR-21, miR-34A, and miR-203 were strongly expressed in mouse keratinocytes, miR-31 was expressed at medium levels, and miR-34b and miR-34c were expressed at very low levels, being essentially undetectable. In addition, the pattern of changes in miRNA expression in keratinocytes from TRα1/TRβ KO mice was almost identical to that observed in adult dorsal skin, since their levels were reduced to a similar extent with respect to those found in Wt mice (Fig. 6). Furthermore, the effect of TRα1/TRβ deletion on target gene expression in keratinocytes was similar to that observed in total skin, as miRNAs depletion was accompanied by a general increase in the levels of their target mRNAs and lower levels of Hairless mRNA (Supplementary Fig. S4).
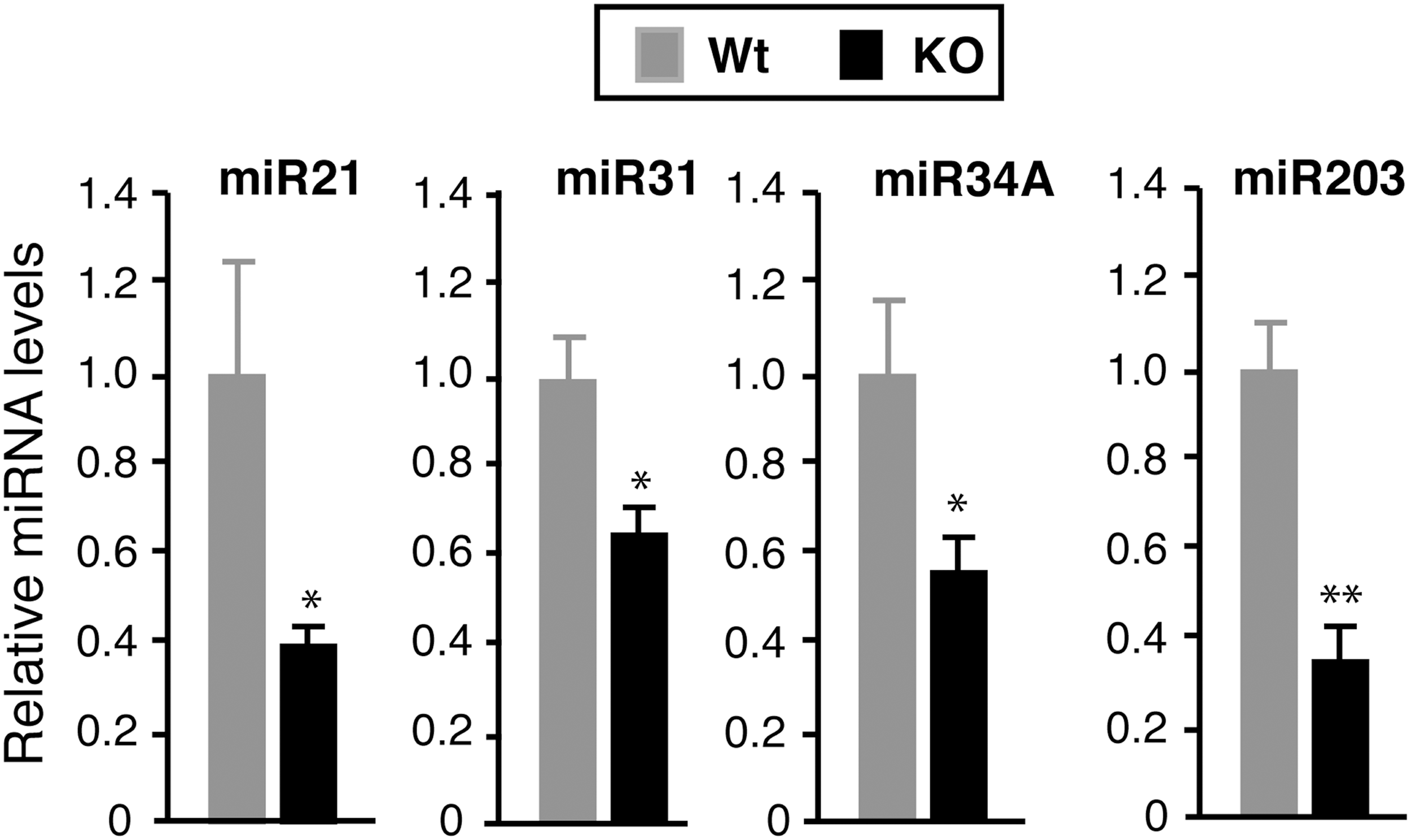
Effect of TR deletion on miRNAs expression in keratinocytes. Keratinocytes were isolated from the skin of newborn Wt (n = 5) and TRα1/TRβ KO mice (n = 7), and the levels of the indicated miRNAs were quantitated. Each miRNA is expressed relative to the value obtained in Wt mice. Data are means ± SE. Asterisks indicate statistically significant differences between Wt and KO mice (*p < 0.05; **p < 0.01).
TRα1 plays a predominant role in skin miRNA regulation
Since both TRα1 and TRβ1 are expressed in the skin (21,28,61,62), next miRNA levels were compared in the dorsal skin of adult Wt mice and in single KO mice lacking either TRα1 or TRβ. As shown in Figure 7, the reduction in the miRNAs analyzed was much more accentuated in mice lacking TRα1 than in mice lacking TRβ. This result suggests that although TRβ could contribute to miRNA regulation, TRα1 appears to be the main receptor isoform responsible for the control of miRNA expression by TRs in the skin. This was also suggested by the finding that the increase in ERK and AKT phosphorylation, as well as in expression of miRNA targets, observed in the double TRα1/TRβ KO mice was also observed in TRα1 mice, while TRβ deletion did not induce this change (Supplementary Fig. S5).
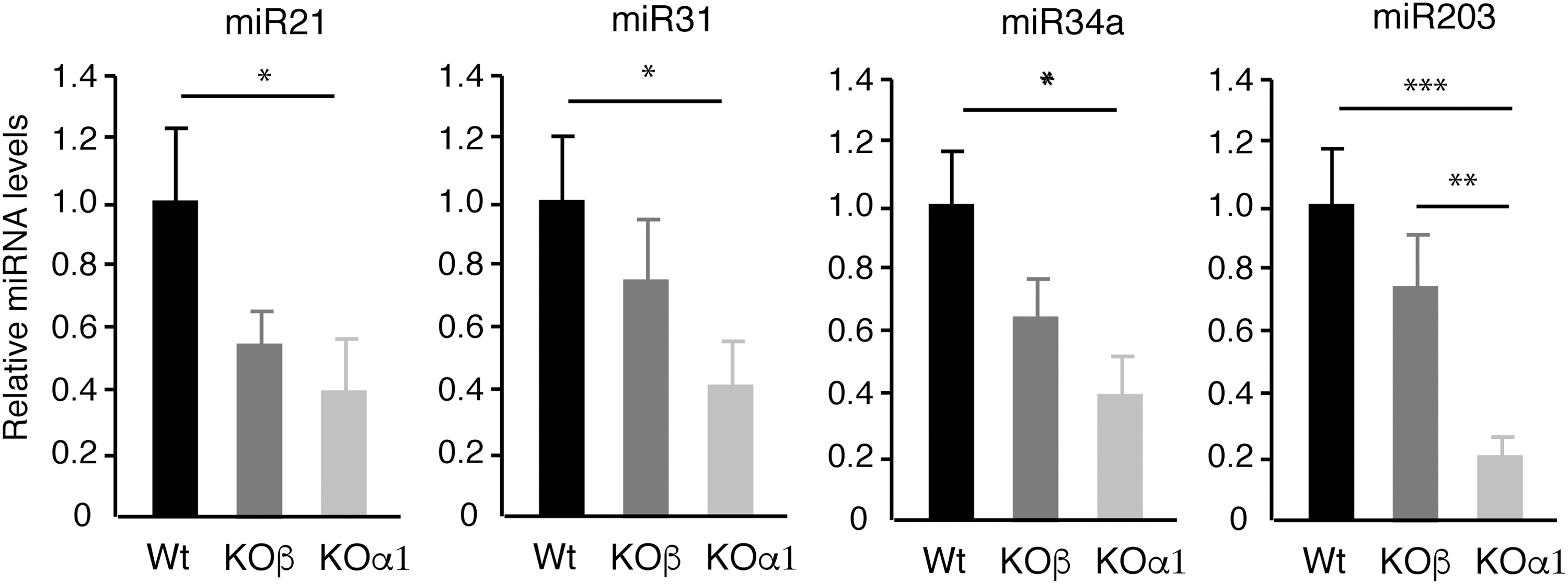
Effect of TRα1 or TRβ deletion in skin miRNA expression in mice. The levels of miRNAs were determined in the dorsal skin of Wt mice and single KO mice lacking either TRα1 (KOα1) or TRβ (KOβ; n = 6). Data are shown as mean values ± SE. *p < 0.05; **p < 0.01; ***p < 0.001 vs. Wt animals.
To analyze the role of both TR isoforms in miRNA expression further, a gain-of-function experiment was conducted by transfecting cultured mouse MCDA3 keratinocytes with an empty vector or with expression vectors for TRα1 or TRβ1. As shown in Figure 8, incubation with T3 had no major effects on miRNA levels in this cell line, indicating low receptor levels in these cells, and the same occurred in TRβ1-expressing cells. In contrast, there was a significant T3-dependent increase in the levels of miR-21, miR-34A, miR-34C, and miR-31 in TRα1-expressing cells, reinforcing the concept that TRα1 is the major regulator of these miRNAs in keratinocytes.

TRα1 but not TRβ1 enables triiodothyronine (T3)-dependent induction of miRNA expression in cultured keratinocytes. MCDA3 keratinocytes were transfected with an empty vector (–) or with vectors encoding TRα1 or TRβ1. The levels of the indicated miRNAs were determined in untreated cells and in cells treated with 5 nM of T3 for 36 hours. Data are expressed as fold change with respect to the values obtained in the corresponding T3 untreated cells and are shown as mean values ± standard deviation. *p < 0.05; **p < 0.01; ***p < 0.001.
Discussion
It has been known for decades that thyroidal status has an important impact on the skin, and previous work has shown that mice devoid of thyroid hormone signaling by genetic ablation of their nuclear receptors show a strong skin phenotype characterized by defects in epidermal proliferation, hair growth, wound healing, and stem-cell function, as well as an altered response to chemical carcinogenesis (19 –21,28,35). There is increasing evidence that all these processes are controlled by several miRNAs, which play a crucial role in epidermal homeostasis and disease (10). Unbiased analysis of the miRNome in adult mice demonstrates that TRα1/TRβ deletion leads to changes in the levels of >50 dorsal skin miRNAs, indicating that these receptors could play an important role as regulators of miRNAs expression in this organ. Given that each one of these small RNAs is predicted to bind to >100 target mRNAs, TRs are likely to posttranscriptionally regulate a large set of skin genes as a consequence of the changes in miRNA expression. Notably, among the miRNA species that were downregulated in these double TR KO mice were miR-21, miR-31, the miR-34 family, and miR-203. Although future studies on the function of other miRNAs differentially expressed in the skin of TR-deficient mice will be needed to define the role of miRNA regulation by the receptors fully, attention was centered in this study on the above-mentioned miRNAs with well-known roles in the skin.
A similar reduction in miR-21, miR-31, miR-34, and miR-203 levels was observed in the dorsal skin from male and female adult TR KO mice and in isolated keratinocytes from newborn mice, suggesting that these cells are the primary targets for the regulation of miRNA abundance by the TRs and demonstrating that this deficiency appears early in life, is sex-independent, and lasts through adulthood. Furthermore, TRβ null mice showed a much milder reduction in miRNA levels than the animals lacking TRα1, suggesting that the binding of the thyroid hormones to TRα1 is the main mechanism by which these hormones could regulate skin miRNA expression. This was confirmed in a keratinocyte cell line, where expression of TRα1 but not of TRβ1 mediated a significant T3-dependent increase in the levels of the analyzed miRNAs. Previous results indicated that single KO mice present a skin proliferative defect, but not as marked as that found in double KO mice, and that deletion of both TRα1 and TRβ is required to impair follicular proliferation or anagen entry, supporting that both receptors participate in this response (19,28). Those results, together with the present finding that TRα1 is the main responsible for miRNA regulation, show that both TR isoforms can play overlapping but also distinct functional roles in the skin.
Some of the miRNAs downregulated in the absence of TRs are among the most highly expressed in the epidermis (5,40). Although it is difficult to define the minimal number of miRNA copies needed to exert functional consequences, as this could also depend on other factors such as the amount of target mRNA or site efficacy, the abundance of these small RNAs regulated (with the exception of miR-34B and miR-34C) suggests that they should be sufficient to be of functional importance in the skin. Indeed, they have been described to play predominant roles in many skin processes (5,6,40,41,45,47,49,51,63), including the different aspects of the skin phenotype observed in the TR-deficient mice. Thus, it is tempting to speculate that regulation of miRNA expression could underlie some of the defects observed in these animals, as well as in patients with thyroid disorders, particularly hypothyroidism, which often present alterations in epidermal proliferation, frequent alopecia, and impaired wound healing (14,16,25 –27,31).
That the decreased expression of the analyzed miRNAs in the skin of mice lacking both TRα1 and TRβ could have functional consequences is suggested by the finding that many of their previously identified mRNA targets are elevated in the dorsal skin and keratinocytes of these animals. Interestingly, these target genes regulate key signaling pathways that will in turn regulate other downstream genes that control diverse physiological skin processes. Therefore, regulation of components of these crucial signaling pathways should further amplify the intricate circuitry of TR action in the skin. In addition, only a few specific mRNA targets have been quantified, and it is very likely that alterations in other mRNAs, as well as changes in protein output without changes in mRNA, could also occurr, as miRNAs can repress not only transcript levels but also protein translation (2). Hopefully, further studies in which global transcriptome analysis of potential target genes and signaling pathways regulated by these and other TR-regulated miRNAs are analyzed will allow the identification of the crucial miRNAs involved in the receptor effects in skin pathophysiology.
Modulation of miRNA levels by TRs could be a direct consequence of binding of these transcriptions factors to thyroid hormone response elements (TREs) in their regulatory regions to be indirectly due to the regulation of other transcription factors or signaling pathways that in turn regulate transcription of the miRNAs or both. The analyzed miRNAs could follow the first model, as chromatin immunoprecipitation experiments have demonstrated that TRs can associate with the regulatory regions of some of these miRNAs (24,64,65), strongly suggesting that they could be direct T3 target genes in the skin. For instance, TRs can directly bind recognition motifs at the miR-34A promoter and transcriptionally activate the expression of this miRNA in the presence of the thyroid hormone in human cells (65), and in silico analysis of transcription factor binding sites in the mouse miR-34A promoter shows the existence of potential TREs. This would be compatible with the reduced levels of miR-34A in the total skin and isolated keratinocytes displayed by the KO mice lacking TR-dependent thyroid hormone signaling. Whether this also occurs with miR-34A and miR-34B, which are transcribed from a distinct genomic locus, remains to be established, but examination of their promoters also indicates the existence of several putative TREs that could mediate a direct regulation by the receptors. Interestingly, miR-31 and miR-203 also contain possible TR-binding sites in their 5′-flanking sequences. The finding that potential TR recognition motifs are also present in human genes encoding these miRNAs suggests that their regulation by TRs could also presumably apply to humans, although further studies will be needed to demonstrate whether these motifs are functional.
The existence of TREs in the miRNA regulatory regions does not preclude the possibility that the analyzed miRNAs could be also regulated by TRs by an indirect mechanism. p53 is the best-characterized transcriptional regulator of the miR-34 family and of miR-203 (6,40), but p53 is not altered by TR deletion in the skin. In addition, the downregulation of miR-21 in the skin of TR-deficient mice could reflect an increased BMP4 signaling, as this pathway is a key repressor of miR-21 expression in keratinocytes (42). Interestingly, Stat3 is another transcriptional stimulator of miR-21 (52,66), but thyroid hormones can be negative regulators of Stat3 signaling (67), and consequently Stat3 activity is not reduced but rather increased in the skin of TR null mice (19). Moreover, in agreement with the reduced levels of miR-21 in the skin of TR KO mice found by the authors, it has been shown that this small RNA can be activated by thyroid hormones in human hepatoma cells through a TRE present in the miR-21 promoter region (64), but it has also been described that it is negatively regulated by these hormones in BCCs at the transcriptional level through an element located in the miR-21 enhancer region (24). Thus, a complex mechanism appears to underlie the regulation of miR-21 by the TRs, as the miRNA could be a direct target of the receptor, as well as responding to other altered signaling pathways.
The miRNAs analyzed in this study play an important role in the control of epidermal proliferation. However, it is not always easy to reconcile the downregulation of these miRNAs observed in the TR-deficient mice and their proposed role. For instance, members of the miR-34 family, first known to be components of the p53 tumor suppressor network, participate in the regulation of the cell cycle (55,56) and in the control of epidermal cell proliferation (40). On the other hand, miR-203 modulates keratinocyte proliferation and migration and contributes to a proper transition from basal to supra-basal cells, ensuring the induction of differentiation and repression of stemness (5,6,51,68). However, these miRNAs are reduced in TR KO mice with impaired proliferation, indicating the presence of other miRNAs and/or regulatory mechanisms that compensate these effects. Among them, the deficiency of miR-31 in keratinocytes inhibits their hyperproliferation (49), and miR-21-null mice show a reduction in the number of proliferating cells in the epidermis (24), which is compatible with the reduced proliferative potential of the epidermis of mice lacking TRs. Moreover, miR-203 contributes to skin re-epithelialization after skin injury (63), and a reduction in miR-21 causes a significant delay in wound closure with impaired collagen deposition (47), which is also compatible with the retarded wound healing observed in TR-deficient mice (28).
Regulation of miRNAs by TR signaling could also be involved in the observed tumor suppressor role of TRs (69). Thus, miR-21 and miR-31, oncomirs that are often overexpressed in skin tumors (24,52,70,71), are reduced in TR null mice. Moreover, loss of miR-21 significantly inhibits the number of papillomas formed in mice subjected to a two-stage skin chemical carcinogenesis protocol (45), a result also found in the animals lacking TRs with reduced miR-21 levels. Interestingly, at late stages, the tumors formed in TR-deficient mice became more aggressive, showing that these receptors can have divergent effects on cell proliferation and tumor progression (21).
Although the mechanisms underlying the effects of TRs in the skin are complex and likely to be dominated by transcriptional controls, given the diversity of roles that miRNAs can play in various skin processes, these molecules might serve as important mediators of TR functions in this organ. Future studies of miRNA expression in the TRα1/TRβ KO background in pathophysiological contexts such as wound healing, psoriasis, or tumorigenesis in which miR-21, miR-31, miR-34, and miR-203 miRNAs are deregulated, will contribute to defining the role of TRs in these processes. In addition, loss-of-function and gain-of-function studies involving the analyzed miRNAs as well as other miRNAs with altered expression in TR-deficient mice will hopefully clarify whether they play a major role in determining cutaneous phenotypes.
Footnotes
Acknowledgments
We thank B. Vennström and D. Forrest for mice. This work was supported by Grants BFU2011-28058, BFU2014-53610-P, and B2017/BMD-3724 to A.A; and SAF2015-66015-R, RD12/0036/0009, PIE 15/00076, and CB/16/00228 to J.M.P.
Author Disclosure Statement
The authors have nothing to disclose.