
Abstract
Background:
CD147 contributes to increased aerobic glycolysis through which it promotes tumor growth. Accumulating evidence suggests that CD147 exerts a variety of functions in thyroid cancer (TC) progression but the molecular mechanisms and therapeutic value of CD147 remain unclear.
Methods:
CD147 levels in TC tissues were analyzed to assess its relationship with prognosis and disease progression. A microRNA (miRNA) microarray and bioinformatics approach were used to identify microRNA regulators of CD147 through measurement of the expression and functions of these miRNAs in TC tissues and cell lines. Precursor miRNA-transfected cells were used to assess regulation of CD147 by miRNA. The effect of miRNA on TC cells via inhibition of glycolysis through CD147 targeting was also evaluated.
Results:
We found that miR-125a-5p regulates CD147 and is negatively correlated with its expression and function. Moreover, CD147 knockdown or increased miR-125a-5p expression significantly reduced the viability, migration, and invasion of TC cells. Our mechanistic studies demonstrate that, through directly repressing the expression of the CD147 protein, miR-125a-5p suppresses aerobic glycolysis and lactate production and subsequently reduces TC cell viability, migration, and invasion, thereby exerting tumor suppressor functions.
Conclusions:
The novel connection identified between miR-125a-5p and CD147 suggests a new diagnostic and prognostic role for miR-125a-5p and that CD147 inhibition may be a candidate therapeutic target in the therapy of for TC.
Introduction
A
Cancer cells exhibit unique metabolic phenotypes and rely on aerobic glycolysis to support their proliferation and anabolic growth, an observation known as the Warburg effect (3). The Warburg effect allows cancer cells to meet their energetic demands, provides essential carbon and nitrogen used in macromolecule synthesis, and minimizes reactive oxygen species production in the mitochondria, thereby fueling rapid tumor growth and proliferation. Cancer cell metabolism is a multistep process controlled by the aberrant regulation of both coding and noncoding genes. The switch from oxidative to glycolytic glucose metabolism is accompanied by increased expression of the glucose transporter GLUT1 and glucose uptake (4). According to several studies published over the past few decades, alterations in oncogenes and tumor suppressors are responsible for metabolic reprogramming in cancer cells (5). Hexokinase (HK) 2 is a pivotal enzyme in glucose metabolism that catalyzes the irreversible rate-limiting step in the glycolytic pathway by phosphorylating glucose to glucose-6-phosphate in an adenosine triphosphate (ATP)–dependent manner (6). However, the glycolytic phenotype results in increased lactic acid production and tumor cells must increase proton efflux by up-regulating pH regulators, such as proton-pumps and/or monocarboxylate transporters (MCTs) (e.g., MCT1 and MCT4), to prevent cellular acidosis (7).
CD147 is expressed at high levels on the surfaces of various malignant cells, including lung, breast, kidney, colon, prostate, oesophagus, liver, and TC cells (8,9). This protein significantly contributes to tumor growth, metastasis, and angiogenesis and has been described to form complexes with monocarboxylate transporters (MCTs) on the cell surface, suggesting that CD147 plays a pivotal role in regulating cell metabolism (10). Based on recent studies from our group and others, CD147 silencing dramatically reduces glycolytic rate and lactate efflux in carcinoma cell lines, indicating that CD147 is involved in tumor glycolysis; however, a mechanistic understanding of the involvement of CD147 in the reprogramming of glucose metabolism in cancer cells is lacking (10,11).
According to recent studies, microRNAs (miRNAs) play critical roles in energy metabolism, a class of naturally occurring small noncoding RNA molecules of approximately 22 nucleotides (12). Their importance in cancer metabolism has been noted over the past decade, as they regulate the expression of genes whose protein products either directly regulate the metabolic machinery or indirectly modulate the expression of metabolic enzymes, serving as master regulators (13). However, the underlying mechanism by which miRNAs modulate cancer metabolism is still largely unknown.
In this study, we aimed at exploring the molecular mechanisms that CD147 plays in TC. Our results show that CD147 overexpression in a subset of TC samples contributes to distinct clinical outcomes for patients. A comprehensive miRNA (miR) analysis found that CD147 expression is regulated by miR-125a-5p and that CD147 knockdown or increased miR-125a-5p expression significantly reduces the viability, migration, and invasion of TC cells. As shown in our mechanistic studies, miR-125a-5p exerts its tumor-suppressing function by directly repressing the expression of the CD147 protein, thereby reducing aerobic glycolysis and lactate production and subsequently reducing TC cell viability, migration, and invasion. Thus, the miR-125a-5p/CD147 axis is tightly correlated with the malignant phenotype and serves as an important regulator of glucose metabolism in human TC.
Materials and Methods
Human TC tissues
TC (n = 200) and nodular goiter (n = 20) tissues were collected from patients at the Xiangya Hospital Central South University (Changsha, Hunan, China) from January 2010 to June 2016 after receiving approval from the Research Medical Ethics Committee of Xiangya Hospital. The TC tissues included 186 PTC, 10 follicular thyroid carcinoma, 3 medullary thyroid cancer, and 1 anaplastic thyroid cancer (ATC) tissues. Clinical data were obtained from databases; patients did not undergo radioactive iodine therapy or other adjuvant treatments prior to surgery. Pathological grading was performed using standards listed in the seventh edition of the American Joint Committee on Cancer (AJCC) Cancer Staging Manual.
miRNA microarray and bioinformatics prediction
For the miRNA microarray assay, six PTC and adjacent nontumor specimens stored in liquid nitrogen were microdissected for RNA extraction. The miRNA microarray was performed using OneArray Human HmiOA6.2 (Phalanx Biotech), and 246 significantly down-regulated miRNAs were identified (tumor/adjacent nontumor ratio >1.5). Using bioinformatics analyses with DIANAmT, miRanda, Target Scan, miRWalk, and PICTAR5, we cross-compared the miRNAs targeting CD147 (BSG) genes and found that CD147 was a predicted target of miR-125a-5p in several databases. The miRNAs that were predicted by at least four algorithms were subsequently validated to reduce the number of false positives.
Cell lines and transfection
The human TC cell lines TPC-1, K1, BCPAP, FRO, and WRO were provided by the Institute of Medical Sciences at Xiangya Hospital. Cells were maintained in Dulbecco's modified Eagle's medium containing 10% fetal bovine serum, 1 μg/mL penicillin, and 1 μg/mL streptomycin (Invitrogen) at 37°C in a 5% CO2 atmosphere. To knockout or overexpress CD147, cells were grown in complete medium for 24 h and then transfected with CD147-ShRNA (GenePharma Co., Ltd) and CD147 expression vector (CD147-pEGFP-C1; GenePharma), respectively using Lipofectamine™ 2000 (Invitrogen Life Technologies) according to the manufacturer's instructions. GenePharma Co., Ltd synthesized all miRs (miR mimics) and anti-miRs (miRNA inhibitors) used in this study. The miRs were transfected into cells using Lipofectamine™ 2000 (Invitrogen) according to the manufacturer's instructions.
Immunohistochemical staining
Tissue microarray sections (5 μm) were immunostained with antibodies against CD147 (dilution, 1:300; Santa Cruz Biotechnology). Two researchers from Xiangya Hospital who were blinded to the clinical data independently performed immunohistochemical evaluations; cases with discrepant grades were re-evaluated by discussion until a consensus was achieved. A semiquantitative H-score ranging from 0 to 300 was calculated for each specimen by multiplying the distribution areas (0–100%) at each staining intensity level by the intensities, as described in previous reports (14,15).
Luciferase activity assay
Sequences (97 bp) from the 3′-UTR of the human CD147 gene containing the predicted miR-125a-5p binding site (5′-CUCAGGGA-3′) was amplified by PCR and cloned into the EcoRI and XhoI sites or KpnI and XbaI sites of the pGL3-control vector (Promega, Madison, WI). Cells were transfected with miR-125a-5p mimics and the wild type (5′-CTCAGGG-3′) or mutant pGL3/CD147-3′UTR(5′-GAGTCCC-3′) constructs using Lipofectamine 2000 (Invitrogen); all procedures were performed according to the manufacturer's instructions. The activity of the co-expressed Renilla luciferase was used to normalize transfection efficiency.
RNA preparation and quantitative real-time PCR
Total RNA was isolated from cells or specimens using TRIzol (Sangon Biotech) according to the manufacturer's instructions. For stem-loop-based microRNA real-time PCR, 5 μg of isolated RNAs were subjected to reverse transcription using SuperScript III (Invitrogen, CA,). SYBR Green-based quantitative real-time PCR was processed using a Step One Real-Time PCR System (TIANGEN) and a 2 × hot start PCR master mix (TIANGEN) with six replicates for each condition; U6 (GenePharma) was used as an internal control. Human-specific primers were synthesized as follows: miR-125a-5p, forward primer: 5′-AGCGCGTCCCTGAGACCCTTTAAC-3′ reverse primer: 5′-ATCCAGTGCAGGGTCCGAGG-3′ RT primer: 5′-GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACTCACAG-3′ miR-125b, forward primer: 5′-GCGCGTCCCTGAGACCCTAAC-3′ reverse primer: 5′-ATCCAGTGCAGGGTCCGAGG-3′ RT primer: 5′-GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACTCACAA-3′ U6, forward primer: 5′-CTCGCTTCGGCAGCACA-3′ U6 reverse primer: 5′-AACGCTTCACGAATTTGCGT-3′ GLUT1, forward primer: 5′-TTGGCTCCGGTATCGTCAAC-3′ GLUT1 reverse primer: 5′-GATGGCCACGATGCTCAGAT-3′ HK2, forward primer: 5′-AAGGAGATGGAGAAAGGGCTTG-3′ HK2 reverse primer: 5′-AGCCAGGAACTCTCCGTGTT-3′ MCT1, forward primer: 5′-CTCTGGGCGCCGCGAGATAC-3′ MCT1 reverse primer: 5′-CAACTACCACCGCCCAGCCC-3′ MCT4, forward primer: 5′-CCAGGCCCACGGCAGGTTTG-3′ MCT4 reverse primer: 5′-GCCACCGTAGTCACTGGCCG-3′
Western blot analysis
Following washes with phosphate-buffered saline, treated cells were lysed in 200 μL of radioimmunoprecipitation assay buffer (Sangon Biotech) containing protease inhibitors (Sangon Biotech), and 30 μg micrograms of protein from the cell lysates were loaded on SDS polyacrylamide gels, followed by Western blot analysis to detect the levels of the CD147 protein (CD147, Santa Cruz Biotechnology; GAPDH, Abcam).
Migration and invasion assays
For wound-healing experiments, TC cells were seeded in 6-well plates. Once the cells achieved approximately 100% confluence, a 200-μL pipette tip was used to create a scraped line. Cells were cultured in fresh medium for 24 hours and then the distances of migrating cells were imaged with a microscope and the rate of closure was assessed.
Transwell migration assays were performed using 8 μm inserts (BD Biosciences) and 1 × 104 cells for each condition. Transwell invasion assays were evaluated with the same inserts coated with 1 mg/mL Matrigel (Invitrogen) and 2 × 104 cells for each condition. Cells were fixed with 4% formaldehyde and stained with 0.1% crystal violet after 24 hours. The crystal violet stained area was analyzed using ImageJ software, and five randomly selected fields were calculated under a light microscope.
Lactate production and glucose consumption
Cells were cultured in Dulbecco's modified Eagle's medium without phenol red for 15 hours; then, the culture media were harvested to measure the lactate or glucose concentrations. Lactate levels were quantified using a Lactate Assay kit (BioVisionCA) and glucose levels were measured with a glucose assay kit (Sigma-Aldrich). All values were normalized to relative protein levels measured using a bicinchoninic acid (BCA) protein assay.
Cellular glucose-6-phosphate and ATP levels
Cellular glucose-6-phosphate and ATP levels were measured using a Glucose-6-Phosphate Fluorometric Assay Kit (Cayman) and a CellTiter-Glo® Luminescent Cell Viability Assay (Promega), respectively. All values were normalized to the relative protein levels.
Data analysis
All quantified data were analyzed using GraphPad Prism 5 software (GraphPad Software) and are presented as the mean ± standard deviation from at least three independent experiments. Comparisons between different groups were assessed using Student's t-tests and one-way analyses of variance. The correlations between CD147 expression and clinicopathological features were assessed using the chi-squared test or Fisher's exact test. All statistical analyses were performed using SPSS 21.0 and all statistical tests were two sided. The level of significance was set to p < 0.05, p < 0.01, or p < 0.001.
Results
Up-regulation of CD147 in TC tissues and cells correlates with poor histopathology
The clinical features of the 200 TC patients are reported in Table 1. We first evaluated CD147 expression in 200 TC and 20 nodular goiter specimens using immunohistochemistry to investigate the potential significance of CD147 in the development and progression of TC. CD147 was located in a diffuse pattern in the cytoplasm of tumor cells and overexpressed in TC tissues with various degrees of differentiation (Fig. 1A and C); however, cytoplasmic CD147 levels were significantly higher in TC tissues than in nodular goiter tissues (Fig. 1B), and significantly higher across different pathological stages (Fig. 1D). CD147 H-scores for each type of TC is shown in Table 2.

Upregulation of CD147 expression in thyroid cancer (TC) tissues and cells correlated with a poor histopathology. (
The data were subjected to Fisher's exact test.
pT, pathologic.
CD147 expression presented as a semiquantitative H-score: 0 to 300 was calculated for eachspecimen by multiplying the distribution areas (0–100%) at each staining intensity level by the intensities (0, negative; 1, weak staining; 2, moderate staining; and 3,strong staining).
ATC, anaplastic thyroid cancer; FTC, follicular thyroid carcinoma; MTC, medullary thyroid cancer; PTC, papillary thyroid cancer.
Using the CD147 expression level as a biomarker of TC, the area under the receiver operating characteristic curve analysis revealed a value of 0.8246 (Fig. 1E). Immunoblot analysis of lysates obtained from surgical samples of five patients with PTC confirmed increases in CD147 expression in tumor tissues compared to peritumoral tissues (Fig. 1F). CD147/GAPDH expression was significantly different between tumor and peritumoral tissues (Fig. 1G). We also detected CD147 expression in TC cells, including BCPAP, K1, TPC-1, FRO, and WRO cells (Fig. 1H); CD147 was expressed at higher levels in K1 cells than in other cells. As shown in Table 1, cytoplasmic CD147 expression levels closely correlated with lymph node metastasis (p < 0.01; chi-squared test) and depth of tumor invasion (p < 0.01; chi-squaredtest).
CD147 is a direct target of miR-125a-5p in TC
miRNAs have emerged as a class of powerful gene expression regulators, and changes in miRNA expression appear to be a common characteristic of cancers, including TC (14,16). The loss or suppression of miRNAs targeting CD147 may cause aberrant overexpression of CD147 in TC. Therefore, we used a comprehensive bioinformatics analysis as a filter to generate a selective miRNA library for subsequent screening to identify potential upstream miRNA that targets CD147.
Thirty-one miRNAs were successfully identified as candidate miRNAs (Fig. 2A). Quantitative PCR was used to screen the candidate miRNA library and examine the downregulation of miRNAs in PTC samples compared with peritumoral tissues (Fig. 2B). We identified four miRNAs (miR-939, miR-328, miR-342, and miR-125a-5p) that were downregulated in PTC samples (n = 3) compared to peritumoral tissues (n = 3) (Fig. 2C). PTC and peritumoral tissues were obtained from three patients. We transfected these selected miRNA mimics or anti-miRNAs in TC cells to determine whether CD147 expression was selectively regulated by the four aforementioned miRNAs. The miR-125a-5p mimics suppressed CD147 expression in both TPC-1 and K1 cells, especially in K1; however, mimics of miR-939, miR-328, and miR-342 increased CD147 expression in TPC-1 cells. Anti-miR-125a-5p increased CD147 expression in TPC-1 cells, while anti-miR-939, anti-miR-328, and anti-miR-342 decreased CD147 expression in both TPC-1 and K1 cells. These data suggest that miR-125a-5p is a specific regulator of CD147 in TC cells (Fig. 2D and Supplementary Fig. S1; Supplementary Data are available online at
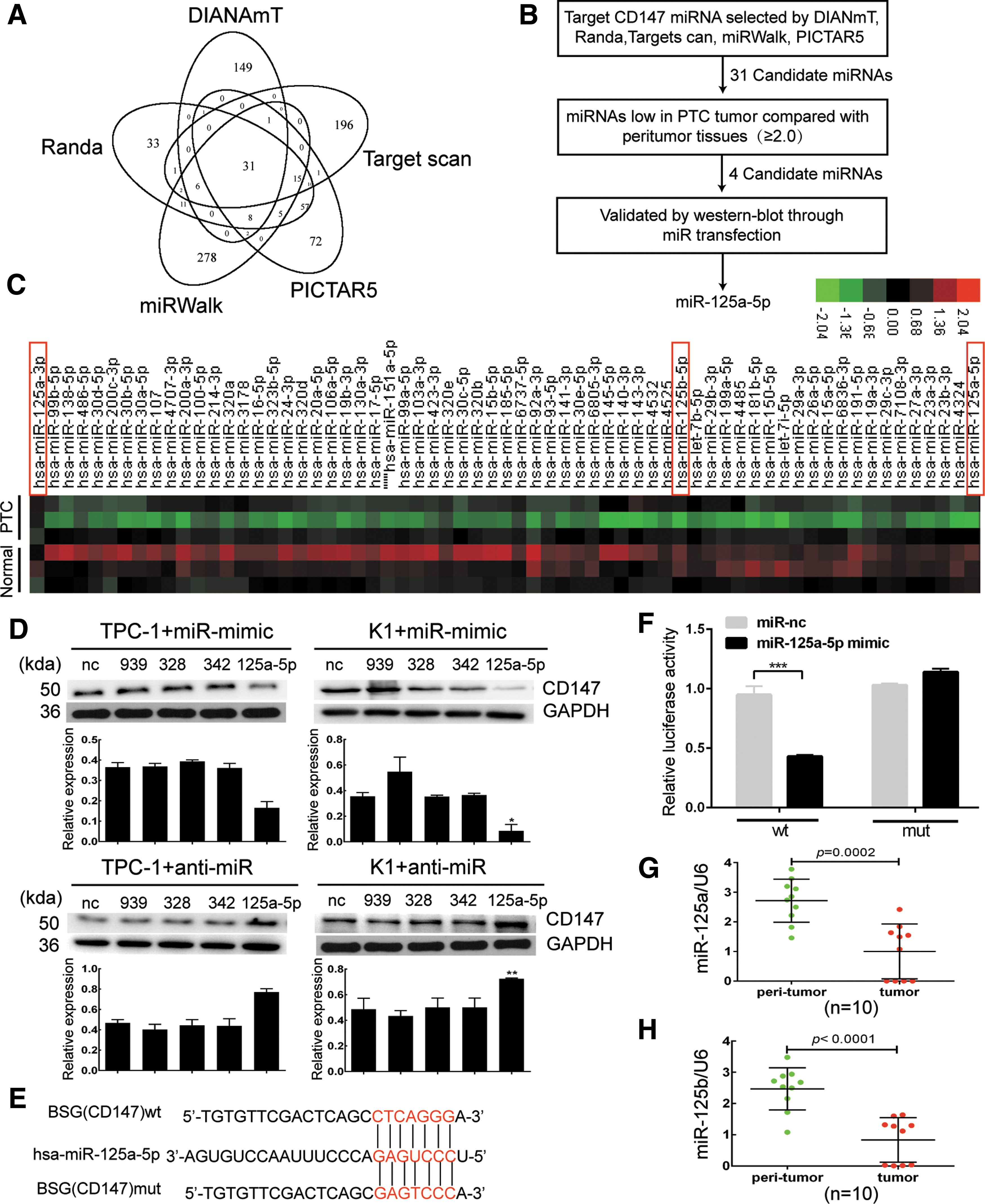
CD147 is a direct target of miR-125a-5p. (
According to the results from the TargetScan algorithm, bases from 461 to 468 in the CD147 (BSG) 3′-UTR have perfect complementarity to the seed sequence of miR-125a-5p. We constructed a mutated CD147 3′-UTR luciferase reporter to substantiate the site-specific repression of CD147 by miR-125a-5p (Fig. 2E), and found that it exhibited a complete restoration of the luciferase activity inhibited by the miR-125a-5p mimics (Fig. 2F). In addition, we estimated the expression levels of miR-125a-5p and miR-125b in PTC tissues using quantitative PCR analysis and normalized their values to U6; miR-125a-5p and miR-125b were significantly down-regulated in the PTC tissues from the 10-patient cohort compared with the peritumoral tissues (Fig. 2G and H). H-scores of these PTC tissues are shown in Table 3. Based on these data, CD147 is a novel direct target of miR-125a-5p in TC cells.
CD147 expression presented as H-score, microRNAs expression presented as quantitative PCR results and normalized to U6.
The expression of miR-125a-5p negatively correlates with cell viability and migration
We next analyzed miR-125a-5p expression in TC cell lines and also examined the effect of miR-125a-5p on TC cell viability and migration in vitro. MiR-125a-5p was expressed at lower levels in TPC-1, K1, BCPAP, and FRO cells than in WRO cells, whereas miR-125b was expressed at lower levels in K1, BCPAP, and FRO cells than in TPC-1 and WRO cells (Fig. 3A and B).
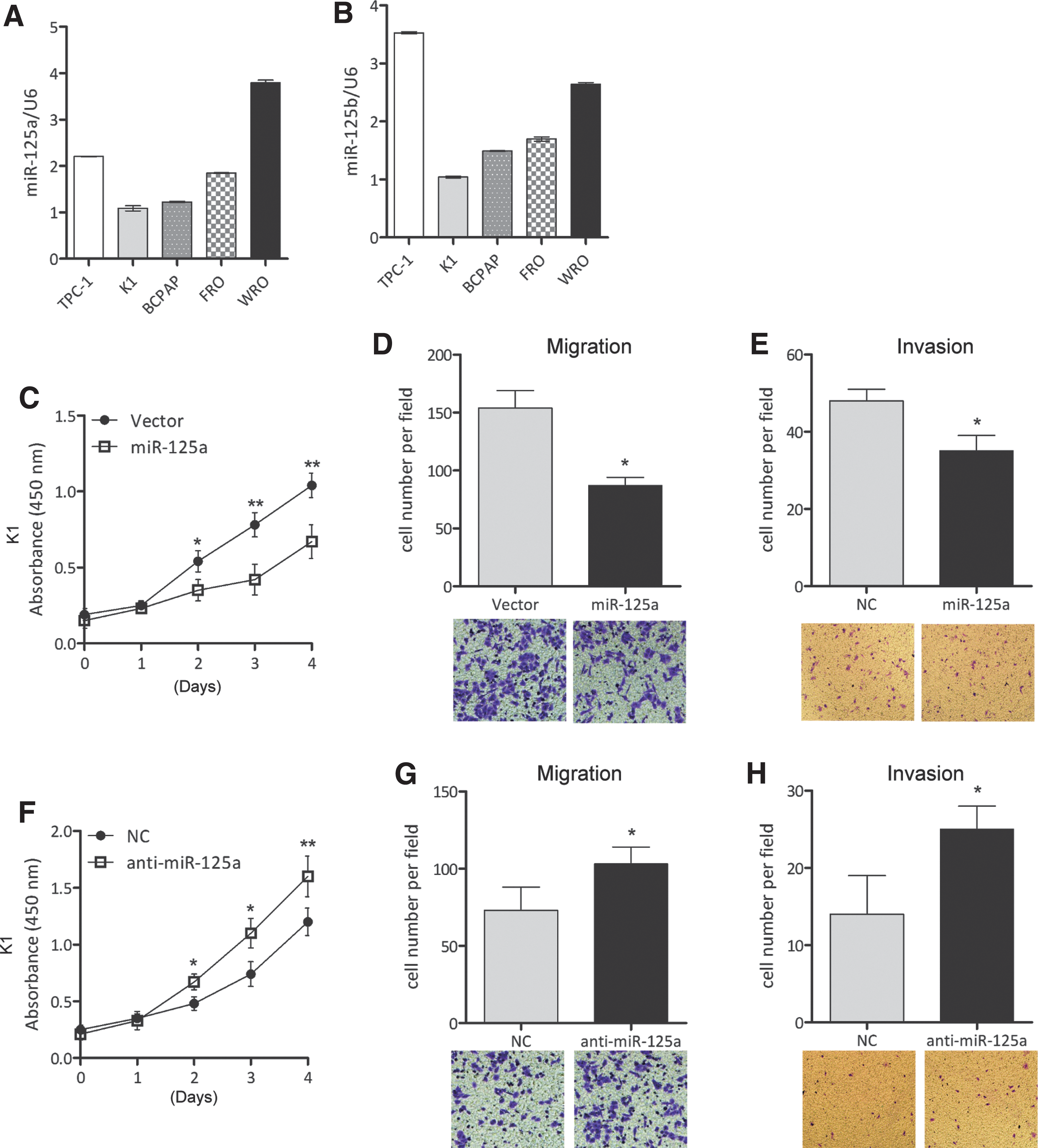
The expression of miR-125a-5p is negatively correlated with cell growth and migration. (
Since CD147 was expressed at higher levels in K1 cells than in the other cells, K1 cells are therefore the best model for assessing the negative relationship between CD147 and miR-125a-5p. The effects of miR-125a-5p on cell viability were evaluated in K1 cells using the MTT assay. Cells transfected with miR-125a-5p mimics were less viable than nontreated cells (vector); however, anti-miR-125a-5p increased cell viability (Fig. 3C and F, and Supplementary Fig. S1). Cell migration was analyzed using transwell assays, and cell invasion was analyzed using a Matrigel-coated transwell assay. MiR-125a-5p mimics reduced cell migration and invasion but anti-miR-125a-5p increased the migration and invasion of K1 cells (Fig. 3D, E, G, and H).
CD147 mediates the effects of miR-125a-5p on TC cell growth and migration
K1 cells were transfected with CD147 or control siRNA prior to transfection with miR-125a-5p mimics or anti-miR-125a-5p, followed by assessments of cell growth and migration, respectively. Cell viability was evaluated using the MTT assay and cell migration was analyzed using wound-healing and transwell assays. Inhibition of miR-125a-5p significantly promoted the growth and migration of K1 cells; however, CD147 knockdown prevented increased growth and migration induced by anti-miR-125a-5p expression (Fig. 4A–E, Supplementary Fig. S1A, and Supplementary Fig. S2A). K1 cells transfected with miR-125a-5p mimics showed attenuated growth and migration but CD147 overexpression restored cell growth and migration (Fig. 4F–J, Supplementary Fig. S1B, and Supplementary Fig. S2B). Thus, the inhibitory effects of miR-125a-5p are partially mediated by targeting CD147.
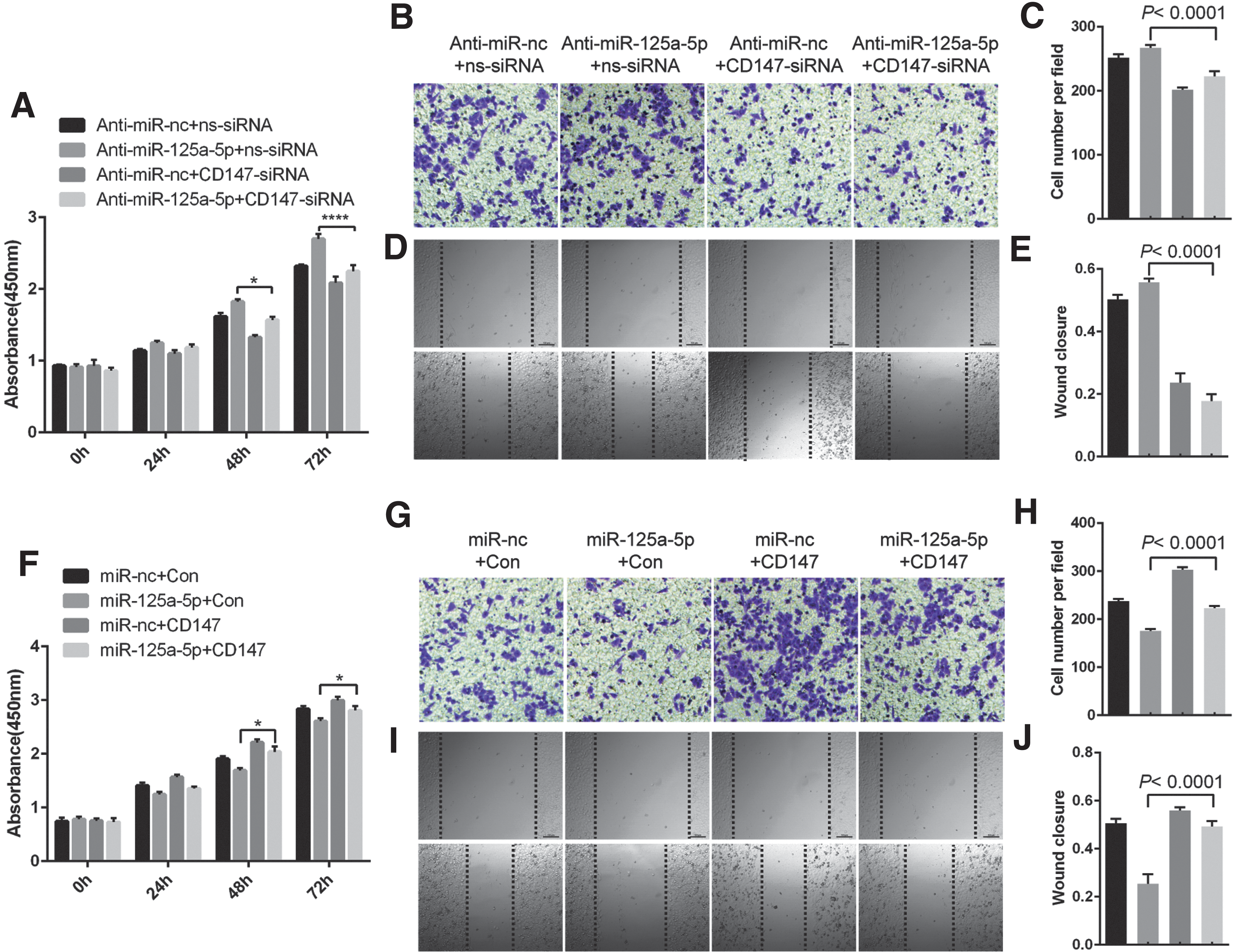
CD147 mediates the effects of miR-125a-5p on TC cell growth and migration. (
The miR-125a-5p/CD147 axis regulates glucose metabolism in TC cells
CD147 promotes the development of cancer by regulating the glycolytic process in cancer cells (10,11,17). It regulates the lactic acid transport process along with co-expressed MCT1 and/or MCT4 (8) while simultaneously up-regulating the expression of glucose metabolism-related proteins, including GLUT1, HK2, MCT1, and MCT4 (18). We first investigated the biological function of the miR-125a-5p/CD147 axis in cancer glycolysis to determine its underlying mechanism. Ectopic expression of miR-125a-5p in human TC K1cells efficiently reduced glucose consumption, glucose-6-phosphate, and
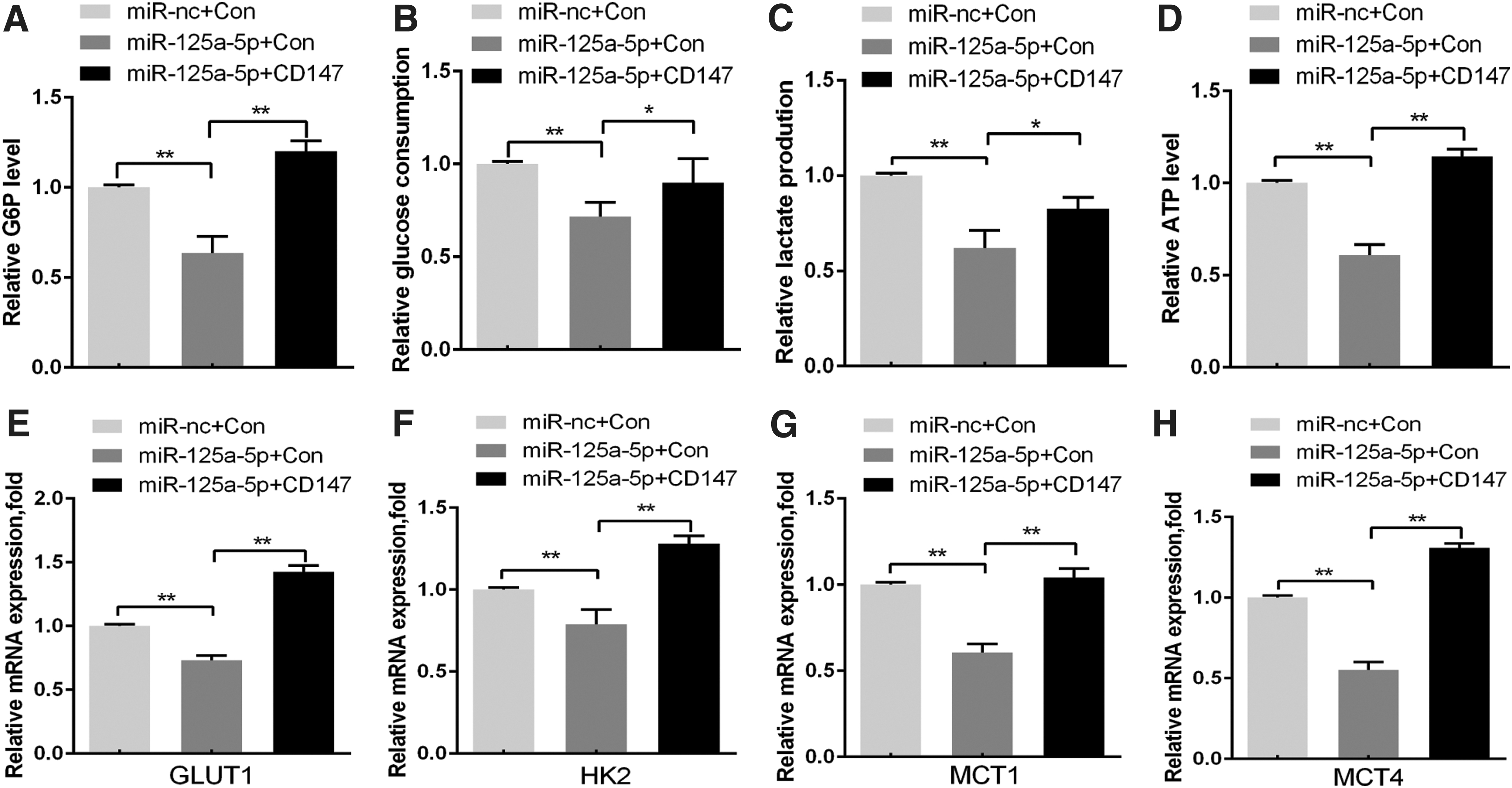
The miR-125a-5p/CD147 axis regulates glucose metabolism in TC cells. (

Metabolic differences between normal and TC cells. Normal cells primarily metabolize glucose to pyruvate for growth and survival, followed by complete oxidation of pyruvate to CO2 through the tricarboxylic acid cycle. TC cells convert most glucose to lactate, regardless of the availability of O2, through the overexpression of some special glucose metabolism-related proteins, including GLUT1, HK2, MCT1, and MCT4, which divert glucose metabolites from energy production to anabolic process to accelerate cell proliferation, migration and invasion. miR-125a-5p inhibits the synergistic effect of CD147 on lactate transporters (MCT1/MCT4) and reduces the viability, migration, and invasion of thyroid cancer cells, which were increased by aerobic glycolysis.
Discussion
Here, we provide evidence that miR-125a-5p is down-regulated in TC tissues and that miR-125a-5p is abnormally expressed in numerous cancers. Down-regulation of miR-125a-5p has been observed in other types of cancers, including liver, colon, lung, breast, and ovarian cancers (19). The causative factors underlying down-regulation of miR-125a-5p in these kinds of cancers remain unclear; however, promoter methylation and the efficiency of transcription factor binding to promoter profoundly influence gene expression levels (20).Their functional disorder is associated with aberrant expression of many carcinogenic and antioncogenes. However, further studies are needed to elucidate mechanisms underlying the down-regulation of miR-125a-5p in TC.
We also found that miR-125a-5p expression was negatively correlated with levels of the CD147 protein, a pivotal effector in the process of tumor-associated glycolysis, and we ascribe significant diagnostic and prognostic value to the down-regulation of miR-125a-5p during TC progression (21,22). Based on our mechanistic studies, miR-125a-5p exerts its tumor-suppressor function by directly targeting and repressing CD147, thereby reducing the expression of glucose metabolism-related proteins, leading to the inhibition of glycolysis and subsequent reductions in TC cell viability, survival, migration, and invasion. CD147 overexpression abolished the inhibitory effects of miR-125a-5p on CD147, indicating that miR-125a-5p specifically inhibits CD147, yet, the therapeutic role of miR-125a-5p in TC remains to be elucidated and we cannot exclude the potential participation of other miRNAs in regulating CD147 expression.
CD147, also known as extracellular matrix metalloproteinase inducer, or Basigin (BSG), is a protein encoded by the BSG gene, and several reports have confirmed its role in tumorigenesis (23). CD147 can be used as a diagnostic and prognostic indicator in patients with TC. According to Tan et al. (2008), CD147 is expressed at significantly higher levels in TC tissues than in healthy or benign thyroid tissues, suggesting that CD147 is a prognostic factor for differentiated thyroid carcinoma (24). Aratake et al. (2010) reported the feasibility of analyzing CD147 expression using fine needle aspiration samples, and this research indicated that an immunocytochemical analysis of CD147 may represent a novel avenue to evaluate the differentiation of TC (25). Results from Omi and colleagues provide in vivo evidence for CD147 up-regulation in follicular thyroid carcinoma and in vitro evidence for epidermal growth factor-stimulated CD147 expression via the PI3K, ERK, and JNK pathways (9).In the present study, we detected CD147 expression in a group of 200 TC specimens and analyzed the relationship between CD147 expression and clinicopathological factors. We found that CD147 expression was significantly correlated with lymph node metastasis and tumor invasion. Nonetheless, the mechanism underlying CD147 overexpression in TC is currently unclear.
We also provide definitive evidence supporting the hypothesis that miR-125a-5p negatively regulates CD147 expression. We verified that miR-125a-5p is reduced in TC tissues, highlighting its potential use as a diagnostic marker for patients with TC. By taking advantage of the publicly available biology databases, we accurately predicted miRNAs that target CD147 and confirmed the findings using miRNA chip assays and bioinformatics. Therefore, the methods used in our primary screen may also be used for other types of cancer.
Studies have reported that miR-125a-5p acts as a tumor suppressor in several kinds of cancers, including TC (26 —29). In this study, miR-125a-5p effectively inhibited the proliferation and invasion of TC cells; however, the underlying mechanism how miR-125a-5p affects TC cells remains unclear. To our knowledge, this report is the first to describe the screening and characterization of the mechanism by which miR-125a-5p regulates glucose metabolism in TC cells. Since lactate is the final product of glycolysis in cancer cells and can easily be quantified using a simple enzymatic reaction, we were able to detect extracellular lactate levels as an indicator of the glucose metabolism rate (30). We also found that miR-125a-5p suppresses lactate production, cellular ATP levels, and glucose consumption of TC cells, which was phenocopied via CD147knockdown. As GLUT1, HK2, MCT1, and MCT4 are critical glycolysis-related proteins and their expression levels are significantly regulated by the miR-125a-5p/CD147 axis, we hypothesized that the miR-125a-5p/CD147 axis likely plays a key role in the aerobic glycolysis of TC cells. However, aerobic glycolysis is difficult to measure in vivo, and current research has only provided in vitro data.
In conclusion, miR-125a-5p and its target gene, CD147, are deregulated in human TC and correlated with clinical outcomes of TC patients. Moreover, miR-125a-5p functions as a tumor suppressor in TC by directly targeting and repressing CD147 expression, thereby inhibiting aerobic glycolysis and subsequently suppressing cancer cell viability, migration, and invasion. Our observations also suggest an important role for miR-125a-5p in glucose metabolism in the TC microenvironment and provide novel prognostic biomarkers for patients with TC.
Footnotes
Acknowledgments
We wish to thank all the patients who participated in this study. This work was supported by the Hunan Science and Technology Project (No. 2016JC2056), Hunan Provincial Innovation Foundation for Postgraduate (No. CX2017B073), and Clinical Research Fund of Xiangya Hospital Central South University (No. 2016L05).
Author Disclosure Statement
No competing financial interests exist.