
Abstract
Background:
The intrafollicular space of thyroid follicles is the storage compartment for thyroid hormones. Its pH has been established at around 7.6 at least after thyrotropin (TSH) stimulation. This alkaline intrafollicular pH is thought to be critical for iodide coupling to thyroglobulin and internalization of iodinated thyroglobulin. At least in mice, this alkalinization requires the expression of pendrin (Slc26a4) within the apical membrane, and a lack of pendrin results in acidic follicular lumen pH. Yet, the mechanism importing HCO3 − into the cytoplasm is unknown. This study investigated whether the rather ubiquitous sodium bicarbonate cotransporter NBCe1 (SLC4A4) might play this role. It also examined which variant was expressed and where it was localized in both rat and human thyroid tissue. Lastly, the dependence of its expression on TSH was studied.
Methods:
Reverse transcription polymerase chain reaction, immunofluorescence, and Western blotting were used to test whether TSH stimulated NBCe1 protein expression in vivo. Subcellular localization of NBCe1 was performed using immunofluorescence in both rat and human thyroid. Cultured thyroid cells were also used to attempt to define how TSH affects NBCe1 expression.
Results:
Only transcripts of the NBCe1-B variant were detected in both rat and human thyroid. Of interest, NBCe1-C was not detected in human tissues, not even in the brain. On immunofluorescence microscopy, the immunostaining of NBCe1 mainly appeared in the basolateral membrane upon stimulation with TSH. This TSH induction of basolateral membrane expression of NBCe1 protein was confirmed in vivo in rat thyroid and in vitro on human thyroid slices.
Conclusions:
This study demonstrates the expression of the sodium bicarbonate cotransporter NBCe1-B in rat and human thyroid. Additionally, the data suggest that TSH blocks the degradation of NBCe1 protein by trafficking it to the basolateral membrane. Hence, TSH increases NBCe1 half-life without increasing its synthesis.
Introduction
Sodium bicarbonate cotransporter (NBC) mediates the obligatory coupled movement of Na+ and HCO3 − ions across the plasma membrane into many cells (1) and thus plays an important role in the regulation of acid-base homeostasis in intracellular fluid. In epithelia, it may also act to secrete or reabsorb bicarbonate ions and is located within the basolateral membrane. NBC belongs to the large family of solute carriers (SLC4), which is divided into four Cl−-HCO3 − exchangers (AEs) and five NBC. There are two electrogenic NBC (NBCe1 and NBCe2) and three electroneutral ones (1). Within the NBC subfamily, NBCe1 is ubiquitously expressed and has been the best characterized up to now. This study was devised to detect the expression of NBCe1 in the rat and human thyroid. Initially, three main NBCe1 splice variants arising from the same gene in humans and rodents (SLC4A4) (2,3) were described. The “kidney” NBCe1 variant (NBCe1-A; kNBC1/SLC4a4-A) encodes 1035 amino acids (4 –6). Two isoforms were described from rat brain. NBCe1-B (rb1NBC1/SLC4a4-B) encodes 1079 amino acids, with 41 N-terminal amino acids of NBCe1-A being replaced by 85 distinct amino acids in NBCe1-B (7) and NBCe1-C (rb2NBC/SLC4a4-C). NBCe1-C is similar to NBCe1-B except for a 97 bp deletion that replaces the 46 C-terminal residues of NBCe1-B, giving rise to 61 unique C-terminal amino acids (7).
The sequence of the “pancreatic” NBCe1 open reading frame (pNBC1) was published in 1999 (8) with 91% similarity in total throughout the entire sequence. However, in “NCBI staff-curated” database sequences, this sequence actually corresponded to one of the brain isoforms, NBCe1-B (7).
Two novel NBCe1 variants, NBCe1-D and NBCe1-E, have been identified in mouse reproductive tract tissues. These two variants contain a deletion of 27 nucleotides at the 3′ end or exon 6, resulting in a deletion of nine amino acids (9). Their expression was not examined in this study.
NBCe1 transcripts and their corresponding proteins have been identified in several tissues other than the kidney, pancreas, and brain (10). NBCe1-A, the first member cloned (2), was found to be located in the basolateral membranes of the proximal tubules of salamander and mammalian kidney, and NBCe1-B was found in the basolateral membrane of rat and human pancreatic duct cells (11 –15).
In 1998, Abuladze et al. (8) demonstrated that the human thyroid expresses NBCe1-B mRNA on Northern blotting. However, thyroid expression of NBCe1 has not yet been studied further. This study investigated whether NBCe1 is expressed in rat and human thyroid epithelial cells. Then, the study explored which variants are present, where the corresponding proteins are located in the cell, and whether their expression varies according to thyroid status.
Methods
Tissue samples and ethics statement
Female Wistar rats about two months old and weighing 200 g were purchased from Janvier Labs (Le-Genest-Saint-Isle, France). All experimental procedures on rats were conducted in accordance with the protocol approved by the Ethical and Animal Welfare Committee of the Université Libre de Bruxelles, Brussels, Belgium (protocol number 375N). Human thyroid tissue was obtained from surgical resection of normal tissue adjoining solitary nodules or glandular carcinoma according to a protocol approved by the Ethics Committee of Erasme Hospital, Brussels, Belgium.
Rat treatments
Following one week of acclimation, rats were randomly assigned to four experimental groups of four rats and kept for up to four weeks on various treatments: (i) control, (ii) 2 mg/L thyroxine (T4), (iii) 250 mg/L methylmercaptoimidazole (MMI), and (iv) 10 g/L sodium perchlorate (ClO4 −). Unless specified, all chemicals were purchased from Sigma–Aldrich (St. Louis, MO) and given in their drinking water. The rats were housed and handled according to the rules of the Belgian regulations for animal care. The animals were anesthetized with ketamine hydrochloride and xylazine, and their thyroids were quickly dissected and processed, as detailed below.
Cell culture
Follicles were isolated by collagenase digestion and differential centrifugation from fresh human thyroid, as previously described (16). The follicles were cultured in the following medium: Dulbecco's modified Eagle's medium:Ham's F12:MCDB 104 medium (2:1:1; Life Technologies, Merelbeke, Belgium) supplemented with 2 mM sodium pyruvate, 100 IU/mL penicillin, 100 μg/mL streptomycin, 2.5 μg/mL Fungizone (Life Technologies), 5 μg/mL insulin (Sigma–Aldrich), and 40 μg/mL ascorbic acid. The thyrocytes were cultured for four days. To ensure optimal spreading of the follicles, 1% fetal bovine serum (FBS; Life Technologies) was added for the first 24 h. The cells were then treated with the tested agents.
PCCl3 cells were grown in Coon's modified Ham's F12 medium (Life Technologies) supplemented with 5% decomplemented FBS, 5 μg/mL transferrin, 1 μg/mL insulin (Sigma–Aldrich), 1 mIU/mL thyrotropin (TSH), and antibiotics (see above). The cells were deprived of TSH for three days before stimulation for 24, 48, and 72 h with 1 mIU/mL TSH or 10 μM forskolin (Calbiochem, Merck Biosciences, Darmstadt, Germany).
Hormone measurement
Hormones were measured in rat serum samples from abdominal artery blood. Total T4 measurement was performed using solid-phase 125I radioimmunoassay (Coat-A-Count Canine T4; Siemens Medical Solutions Diagnostics, Diagnostic Products Corp., Los Angeles, CA). Serum TSH concentration was measured using a rat TSH radioimmunoassay (MP Biomedicals, Brussels, Belgium).
Immunodetection
Rat thyroid
Tissues were quickly dissected, fixed by 4 h of immersion in 4% (wt./vol.) paraformaldehyde in 0.1 M phosphate buffer at pH 7.4, rinsed in running tap water (for 2 h), before being embedded in paraffin and cut into 10 μm sections with a standard microtome. Alternatively, after fixation, the tissues were transferred to successive grades of sucrose solutions (10%, 20%, and 30% each overnight), embedded in Tissue-Tek OCT™ compound (Sakura Finetek Europe, Leiden, The Netherlands), snap-frozen in cold 2-methylbutane, and stored at −80°C. Using a Leitz cryostat, 15 μm sections were cut, mounted on poly-L-lysine-coated slides, and stored at −20°C. They were used for the detection of the thyroid-specific iodinated thyroglobulin (Tg-I) and the sodium–iodide transporter (NIS), as previously described (17). In brief, after incubation with primary antibodies, mouse anti-Tg-I antibody B1 (1:2000; a gift from Dr. C. Ris-Stalpers, University of Amsterdam, Amsterdam, The Netherlands) and rabbit anti-NIS antibody (1:20,000; a gift from Dr. N. Carrasco, Yale School of Medicine, New Haven, CT), tissue sections were incubated with secondary antibodies conjugated to a peroxidase-labeled polymer (EnVision detection; Dako, Glostrup, Denmark) for Tg-I, and with a goat biotinylated secondary antibody (Jackson ImmunoResearch, West Grove, PA) and an avidin-conjugated horseradish peroxidase ABC system (Dako) for NIS. Labeling was detected with aminoethylcarbazole (Dako) for Tg-I and diaminobenzidine (Dako) for NIS, and sections were slightly counterstained with Mayer's hematoxylin. The anti-NBC used in Figure 3 was obtained from Chemicon International (Temecula, CA) and was replaced with ab187511 from Abcam (Cambridge, United Kingdom; Fig. 5) when the Chemicon antibody was no longer available. Subcellular localization of NBCe1 was performed on both rat and human thyroid (Fig. 5) using immunofluorescence labeling. Again, briefly, tissue sections were incubated with a blocking solution of 10% (v/v) normal goat serum in Tris-buffered saline solution (TBS; 50 mM Tris–HCl, 110 mM NaCl, pH 7.5). The slides were then submitted to an epitope unmasking procedure in which the slides were immersed in a 0.01 M citrate buffer (pH 6) and boiled in a microwave oven for 10 min (2450 MHz, 850 W). Incubation with 1:100 NBCe1 antibody (Abcam) was performed overnight in 1:100 blocking serum diluted in TBS. After the tissues were rinsed three times in TBS, they were incubated for 30 min with a goat anti-rabbit antibody conjugated to Alexa 488 at 1:100 dilution (Molecular Probes, Ghent, Belgium). Immunolabeling was followed by nuclear propidium iodide staining. Lastly, after two rinses in TBS, the cryostat slides were washed in water and mounted using Glycergel (Dako) supplemented with 2.0% of the anti-fading compound 1,4-diazobicyclo[2.2.2]octane (DABCO; Sigma–Aldrich). The sections were examined under a Zeiss Axioplan microscope, and images were acquired using an AxioCam HRc camera.
Human thyroid
Human thyroid tissue was cut into thin (300 μm) slices using a Stadie-Riggs microtome (Arthur Thomas, Philadelphia, PA). The slices were incubated for 24 h in culture medium with or without TSH (3 mIU
Western blot analysis
Protein samples (30 μg) were extracted from the thyroids of rats treated for four weeks, as previously described (18). Human thyroid cell cultures and PCCl3 cells were scraped in ice-cold RIPA lysis buffer (50 mM Tris-HCl, pH 7.4, 150 mM NaCl, 1% NP-40, 0.25% sodium desoxycholate, supplemented with Roche protease inhibitor tablets) and centrifuged at 21,250 g for 15 min at 4°C. All samples were then solubilized using Laemmli buffer (60 mM Tris-HCl, pH 6.8, 2% sodium dodecyl sulfate [SDS], 100 mM dithiothreitol supplemented with 100 mM sodium fluoride, 66.7 μg/mL leupeptine, 1 mM vanadate, and 150 μg/mL benzamidine). This was done for separation by means of SDS-polyacrylamide gel electrophoresis on 8% polyacrylamide gels, and blotting onto a nitrocellulose membrane. After incubation with the Odyssey Blocking Buffer (LI-COR Biosciences GmbH, Bad Homburg, Germany), the membranes were subsequently incubated with the polyclonal rabbit anti-NBCe1 antibodies AB3204 (1:200; Chemicon; Fig. 3B) and ab187511 (1:100; Abcam; Figs. 4 and 7), as well as with rabbit anti-NIS (1:2000), monoclonal mouse anti-Tg-I (1:5000), and a rabbit anti-β actin antibody (1:750). The membranes were then washed three times in phosphate-buffered saline and incubated with either goat anti-mouse secondary antibody conjugated to IRDye 680 (for Tg-I) or an anti-rabbit secondary antibody conjugated to IRDye 800 (for the other proteins). The blotted proteins were detected using the Odyssey infrared imaging system.
To estimate whether TSH impedes the degradation of NBCe1 protein, monolayers of PCCl3 were deprived or not of TSH for three days, treated with 100 μM protein synthesis inhibitor cycloheximide for up to 8 h, and processed for Western blotting to quantify NBCe1 protein expression.
Reverse transcription polymerase chain reaction
Total RNA from PCCl3 cells, islets, pancreas, thyroid, kidney, cerebellum, cortex, hippocampus, and whole brain from rat and human tissues were extracted using the AURUM™ total RNA fatty acid and tissue kit (Bio-Rad Laboratories, Hercules, CA). The quality of the total RNA was analyzed on an Experion automated electrophoresis system (Bio-Rad Laboratories), and the quantity was measured using a NanoDrop spectrophotometer (NanoDrop Technologies, Inc., Wilmington, DE). Reverse transcription was performed on 1 μg total RNA using the iScript™ cDNA synthesis kit (Bio-Rad Laboratories).
The reverse transcription polymerase chain reactions (RT-PCR) were carried out in a final volume of 20 μL containing 1 μL cDNA added to 4 μL GoTaq™ Green 5 × buffer, 0.2 mM dNTP, 0.5 μM each primer, and 0.5 IU GoTaq™ polymerase (Promega, Madison, WI) on the MyCycler thermal cycler (Bio-Rad Laboratories). Specific primers for the different NBCe1 variants of rats and humans were designed on sequences curated by the staff of GenBank, as described in Table 1.
Selection of Primers Used for the Amplification of NBCe1 Variants
Rat NBCe1 primers: The specific forward primer for the 5′ splice kidney variant was based on the 5′ sequence domain (5′-untranslated; nt52-34) of AF027362. The reverse primer was based on the nt292–272 coding sequence (CDS) of AF027362 (yet common to the sequences of all variants) giving an amplicon length of 344 nt. The forward primer for the amplification of both the shorter (X1) and larger (X2) 3′-splice variants of NBC comprised the sequence nucleotide (nt) 2933–3360 of both XM_006250788 (X1) and XM_006250789 (X2). The reverse primer comprised the sequence nt3332–3313 for XM_006250788 (X1), giving an amplicon length of 400 nt, and nt3429–3410 for XM_006250789 (X2), giving an amplicon length of 493 nt. The specific primer for amplification of the splice variant X1 (nt3210–3230) was positioned on the nt3219 deletion site of the CDS of XM 006250788. With the corresponding reverse primer (nt3694–3675), the amplicon length was 485 nt. Localization of these primers is depicted in Figure 1A.
Human NBCe1 primers: The forward primer for amplification of kidney NBCe1-A was based on the specific 5′-sequence of NM_003759 (splice variant 2) corresponding to nt1–19 of the CDS. The reverse primer was based on the sequence common to all of the variants. For NM_003759 this corresponded to nt688–662 of the CDS, producing an amplicon length of 688 nt. Primers amplifying all NBC splice variants were based on sequences common to all of the splice variants. However, this produced amplicons of different sizes: NBCe1-A, based on splice variant 2, kidney, (NM_003759), amplicon primer nt2690–3236 of the CDS, amplicon length 547 nt. NBCe1-B, based on splice variant 1 (NM_001098484 curated from pancreatic sequences), amplicon nt2822–3368, amplicon length 547 nt. NBCe1-C, based on splice variant (NM_001134742), amplicon nt2822–3271 of the CDS, length 450 nt. Splice variant X1 (XM_011532390), amplicon nt2264–2810 of the CDS, length 547 nt. The forward primer for specific amplification of brain NBCe1-C was based on the splicing (deletion) site in the 3′-domain of NM_001134742 (splice variant 3) nt3086–3109/splicing site nt3099 of the CDS. The reverse primer was based on nt3578–3559 of the CDS of the same entry, amplicon length 493 nt. Localization of these primers is depicted in Figure 1B.
A schematic representation of the three NBCe1 variants, as well as the localization of the primers on NBCe1 sequences, is depicted in Figure 1 for rat and human transcripts. The PCR was set to the following conditions for the rat and human transcripts: 33 cycles of 30 s at 95°C, 1 min at 60°C, and 1 min at 72°C, with an initial 90 s denaturation step and a final 5 min elongation step. PCR amplification products were subjected to electrophoresis on a 1.2% agarose gel in TAE buffer (40 mM Tris, 40 mM acetate, and 1 mM EDTA in 0.5 μg/mL ethidium bromide). The DNA was visualized by means of UV transillumination using a gel-documentation apparatus (Fusion FX5; Vilber Lourmat, Marne-la-Vallée, France).
Quantitative RT-PCR
Quantification of NBCe1 mRNA expression in PCCl3 cells and in control and MMI-treated rats was performed on a CFX96 real-time system and C1000 thermal cycler (Bio-Rad Laboratories) using iQ
Statistical analysis
The normality of distributions was assessed by means of the Shapiro–Wilk and Kolmogorov–Smirnov tests, with the Lilliefors significance correction test. Data sets with normal distributions were compared using parametric tests. Data sets containing at least one non-normal distribution were compared using the nonparametric Mann–Whitney test. Statistical tests were performed using IBM SPSS Statistics for Windows v22 (IBM Corp., Armonk, NY) and OriginPro 8 (OriginLab, Northampton, MA). Data are presented as the mean ± standard error of the mean. Differences between experimental and control values were considered significant at p < 0.05.
Results
RT-PCR analysis for NBCe1 expression in PCCl3 cells and in rat and human thyroids
RT-PCR was used to analyze the expression of NBCe1 mRNA in human and rat thyroids. Three pairs of primers were selected for the different splice variants of humans and rats (Table 1). The position of these primers on the sequences of the different variants is depicted in Figure 1A for rats and in Figure 1B for humans. The first pair in each group was specific for the kidney variant, NBCe1-A. The second pair was designed to amplify all NBCe1 variants but gave different amplicon sizes for “NBCe1-A plus NBCe1-B” and “NBCe1-C.” This was to estimate the relative amount of the variants. As NBCe1-A is only expressed in the kidney, the relative amount of NBCe1-B could be estimated. The third pair was specific for NBCe1-C and enabled this variant to be detected with higher sensitivity.
RT-PCR amplification of the NBCe1-A specific region yielded a fragment of the expected 344 bp size in rat kidney, which had higher expression compared to the islets and cerebellum. No amplification products were obtained in rat PCCl3 cells, thyroid, cortex, or whole brain (Fig. 2A). Amplification of the human NBCe1-A-specific region (Fig. 2B) was only observed in the kidney (688 bp amplicon), whereas no amplification was observed in human thyroid, pancreas, hippocampus, cerebellum, or cortex.

Schematic localization of primers used for the amplification of rat (

Reverse transcription polymerase chain reaction (RT-PCR) analysis with NBCe1 variant-specific primers in rat thyroid tissues and rat PCCl3 cells (
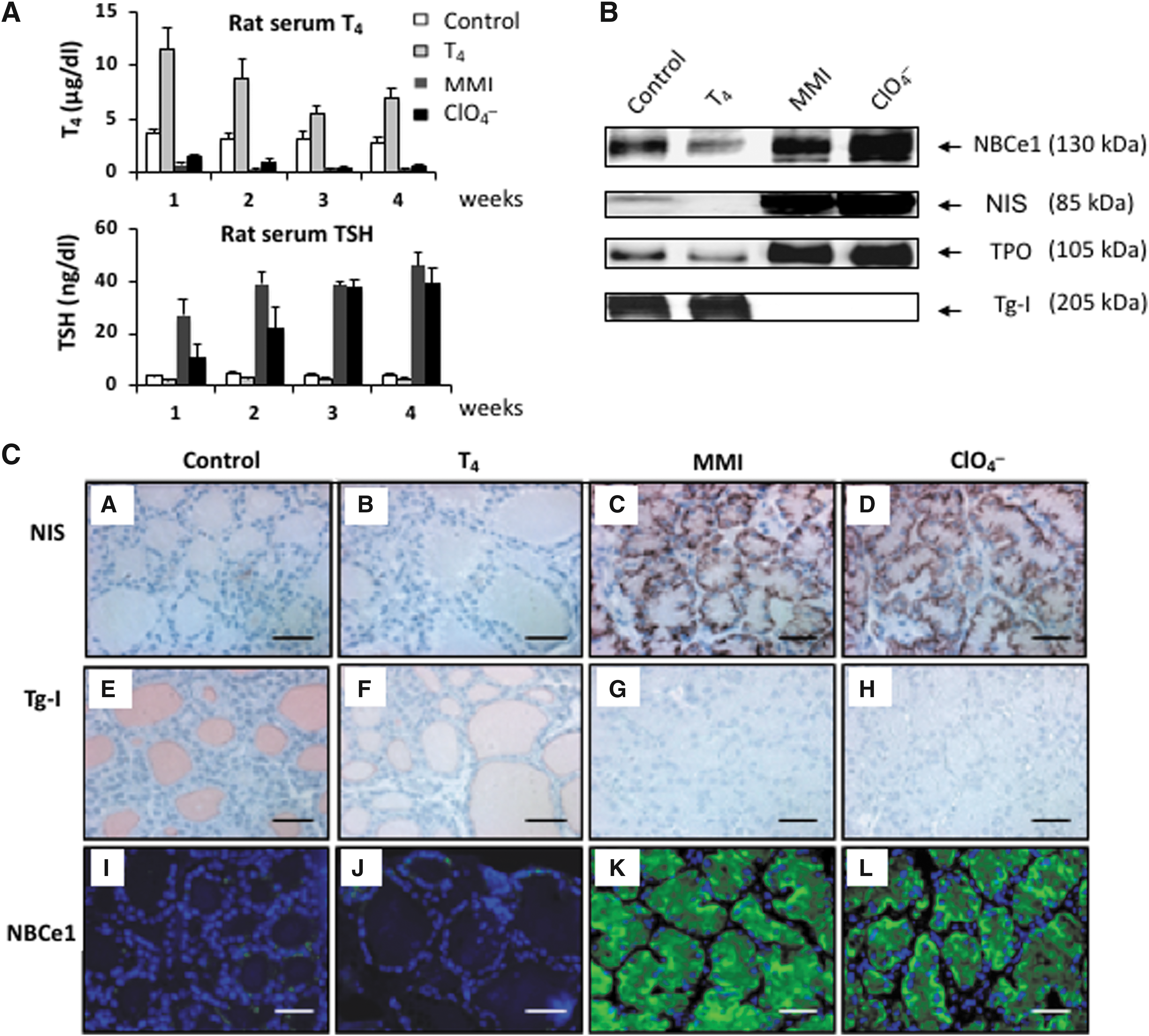
Effect of treatment of rats with H2O (control), thyroxine (T4), methylmercaptoimidazole (MMI), or sodium perchlorate (ClO4
−)for four weeks. (
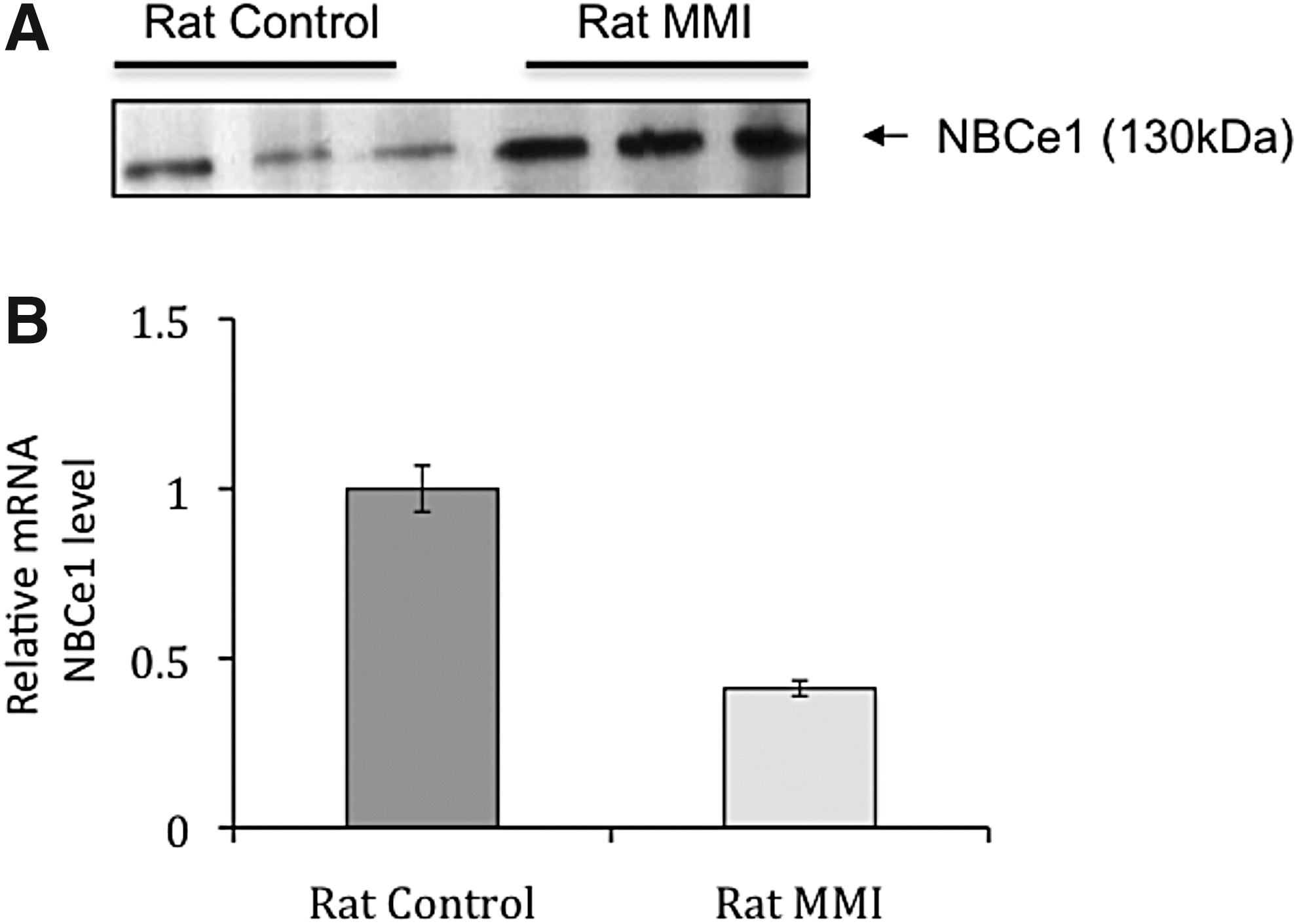
Expression of NBCe1 protein and transcripts in the thyroid of controls versus MMI-treated rats. (
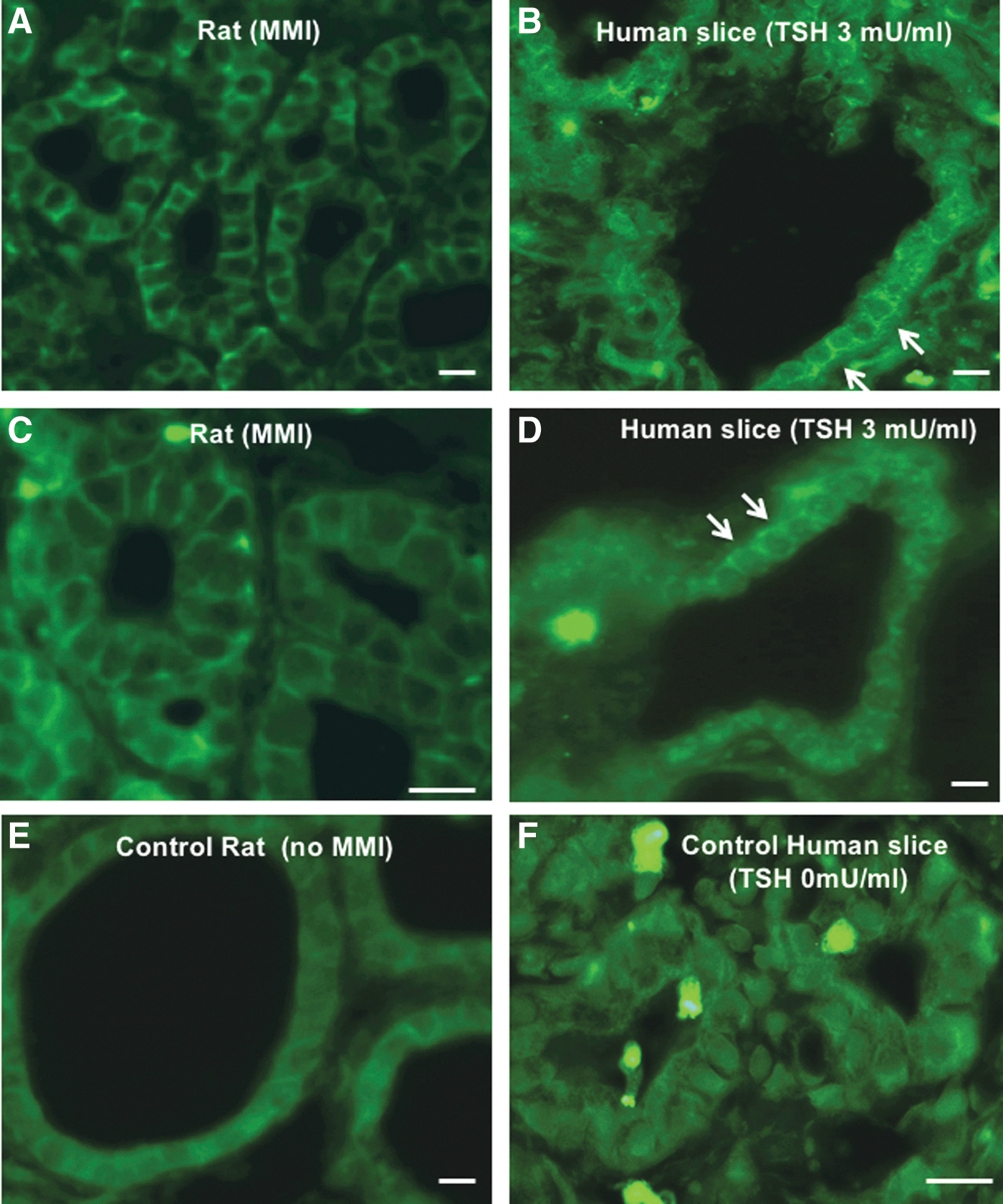
Subcellular immunolocalization of NBCe1 in rat (
Using primers that amplify two rat isoforms at the same time (NBCe1-B and NBCe1-C), transcripts of the expected 497 bp size corresponding to NBCe1-B (without deletion) were observed in PCCl3 cells, pancreatic islets, thyroid, kidney, cerebellum, cortex, and whole brain (Fig. 2A). Amplicons of both 497 and 400 bp (400 bp is expected from the NBCe1-C variant with deletion) were simultaneously amplified in all rat brain tissues. In human tissues, using primers designed to amplify both NBCe1-B and NBCe1-C, only a 547 bp amplicon corresponding to the NBCe1-B transcript was detected in pancreas, thyroid, kidney, and all human brain tissues (Fig. 2B).
NBCe1-C transcripts were also specifically amplified. The results confirmed massive amounts of NBCe1-C transcripts in all rat brain tissues, as well in rat PCCl3 cells and kidney (Fig. 3C). Amplification of this variant was at the limit of detection in rat thyroid (Fig. 2A). Using specific primers for human NBCe1-C, discrete amplification was found in kidney tissue only (Fig. 2B) and, notably, no NBCe1-C transcripts in human brain tissues.
In vivo modulation of TSH level in rats
The expression of thyroid-specific proteins varies with thyroid activity and is mainly influenced by TSH. NBCe1 protein expression was therefore studied in four groups of four rats subjected to various treatments to increase or decrease serum TSH levels. Figure 3A summarizes the TSH and T4 levels in the four groups. As expected, administering T4 downregulated serum TSH values, while administering MMI or ClO4 − upregulated serum TSH concentration. T4 levels increased in the T4-treated animals, while they slightly decreased over time in the MMI and ClO4 − groups compared to control animals. TSH levels decreased in T4-treated rats, whereas they significantly increased and rapidly plateaued as early as the second week of MMI treatment. In the ClO4 − group, TSH levels increased more slowly, rising to 10.3 ng/mL after the first week (compared to 27.1 ng/mL with MMI). However, they then steadily increased and, after three weeks, achieved almost the same plateau as the MMI-treated animals (37.4 ng/mL vs. 38.5 ng/mL; Fig. 3A). After the four weeks of treatment, serum TSH concentrations in the MMI and ClO4 − groups reached their highest level (46.1 and 39.6 ng/mL, respectively), that is, about 10 times higher than in control rats (Fig. 3A).
Western blot analysis of key rat thyroid proteins as a function of TSH level
Western blot analysis of thyroid homogenates of rats subjected to different treatments showed that NIS and thyroperoxidase (TPO) expression decreased in the thyroids of T4-treated rats, while it significantly increased in thyroid homogenates of MMI- and ClO4 −-treated rats compared to control rats (Fig. 3B). Tg-I was almost completely absent in the thyroids of MMI- and ClO4 −-treated rats (Fig. 3B). NBCe1 protein expression fluctuated along the same lines as that of NIS and TPO (Fig. 3B) and paralleled TSH hormone levels, that is, it was downregulated by T4 treatment and upregulated by MMI and ClO4 − treatment (Fig. 3A).
Histology and immunohistochemistry of rat thyroids as a function of TSH level
After four weeks of treatment, the thyroid glands of T4-treated rats were slightly smaller than control thyroids, and the follicles were relatively large and lined with a thin epithelium instead of the cuboidal epithelial cells in controls. Conversely, the thyroids of MMI- and ClO4 −-treated rats were larger than control thyroids (data not shown). Both MMI- and ClO4 −-treated rats showed thyroids with drastically reduced colloid and luminal spaces. On immunohistochemical studies, NIS was barely visible in the basolateral position of a small number of control follicular cells (Fig. 3C, picture A), and was not detected in the flat cells of T4-treated thyroids (Fig. 3C, picture B). Its expression was strongly increased in MMI- and ClO4 −-treated rats (Fig. 3C, pictures C and D). Tg-I was demonstrated in colloid control thyroids (Fig. 3C, picture E) and slightly decreased in the thyroids of T4-treated rats (Fig. 3C, picture F). Labeling was completely abolished in the thyroid glands of MMI- and ClO4 −-treated rats (Fig. 3C, pictures G and H). Thyroidal expression of NBCe1 was slightly decreased by T4 treatment (Fig. 3C, picture J) but drastically increased by MMI and ClO4 − treatment (Fig. 3C, pictures K and L), appearing as basolateral membrane labeling quite similar to NIS. Thus, at least in rat tissues, Western blot analysis and immunohistochemical labeling were in excellent agreement regarding increased NBCe1 protein expression, which appeared in response to the TSH elevation. However, in these experiments, NBCe1 transcript levels were unfortunately not measured. Further experiments were therefore devised to find out whether TSH increases NBCe1 mRNA. Three rats were treated with MMI for three weeks and compared to three controls. While the thyroids of control rats showed variable NBCe1 protein expression levels on Western blots, the MMI-treated rats all exhibited an increase in NBCe1 protein expression (Fig. 4A). Yet, this increase in NBCe1 protein expression did not correlate with an increase in NBCe1 mRNA levels, which decreased (Fig. 4B). This might suggest that TSH-induced increases in NBCe1 expression are not due to increased synthesis but rather to increased translation, stabilization, or inhibition of the degradation pathway.
Subcellular immunolocalization of NBCe1 in rat and human thyroids
Further immunofluorescence experiments were performed to examine the cellular and subcellular localization of NBCe1 in rat and human thyroids. These experiments were carried out with the commercial Abcam antibodies, which do not discriminate between the different splice variants. Rat kidney, where NBCe1 is expressed in the basolateral membrane of the proximal tubules, was used as a positive control (not shown). This approach led to the detection of NBCe1 in both rat and human thyroids. In rat tissues, the labeling was of very low intensity and diffusely scattered intracellularly in the control group (Fig. 5E), whereas it was intensely expressed at the basolateral membrane after three weeks of MMI treatment (Fig. 5A and C). Slices of human thyroid were treated (or not) for one day with 3 mIU/mL TSH. It must be mentioned here that some damage occurred in human slices in vitro. Whereas in the control slices the labeling was diffusely intracellular (Fig. 5F), the slices treated with TSH showed intracellular as well as basolateral membrane labeling (Fig. 5B and D). This suggests that TSH mainly induces NBCe1 exocytosis toward the basolateral membrane, possibly thereby impeding its degradation and increasing its half-life.
Hormonal modulation of NBCe1 expression in cultured thyroid cells
Western blotting was performed using the Chemicon or Abcam antibodies. A 130 kDa band was detected in both human and rat cultured thyroid cells, corresponding to the expected size of the NBCe1 protein. Human thyroid primary cultures (Fig. 6A) or rat PCCl3 cells (Fig. 6B) were stimulated with 1 mIU/mL TSH for up to three days. Compared to unstimulated control samples, NBCe1 protein expression was increased by TSH treatment, although it was more gradual in rat PCCl3 cells than in human primary culture cells (Fig. 6A and B). In response to stimulation with 10 μM forskolin, NBCe1 protein expression increased as well in PCCl3 cells, but no difference was observed between 24 and 72 h of incubation (Fig. 6C and data not shown). Yet, the increase in NBCe1 protein levels with all of these treatments in PCCl3 cells did not change NBCe1 mRNA levels (Fig. 6D), suggesting again that the half-life of the protein is increased rather than its synthesis.

Expression of NBCe1 protein in cultured thyroid cells. (
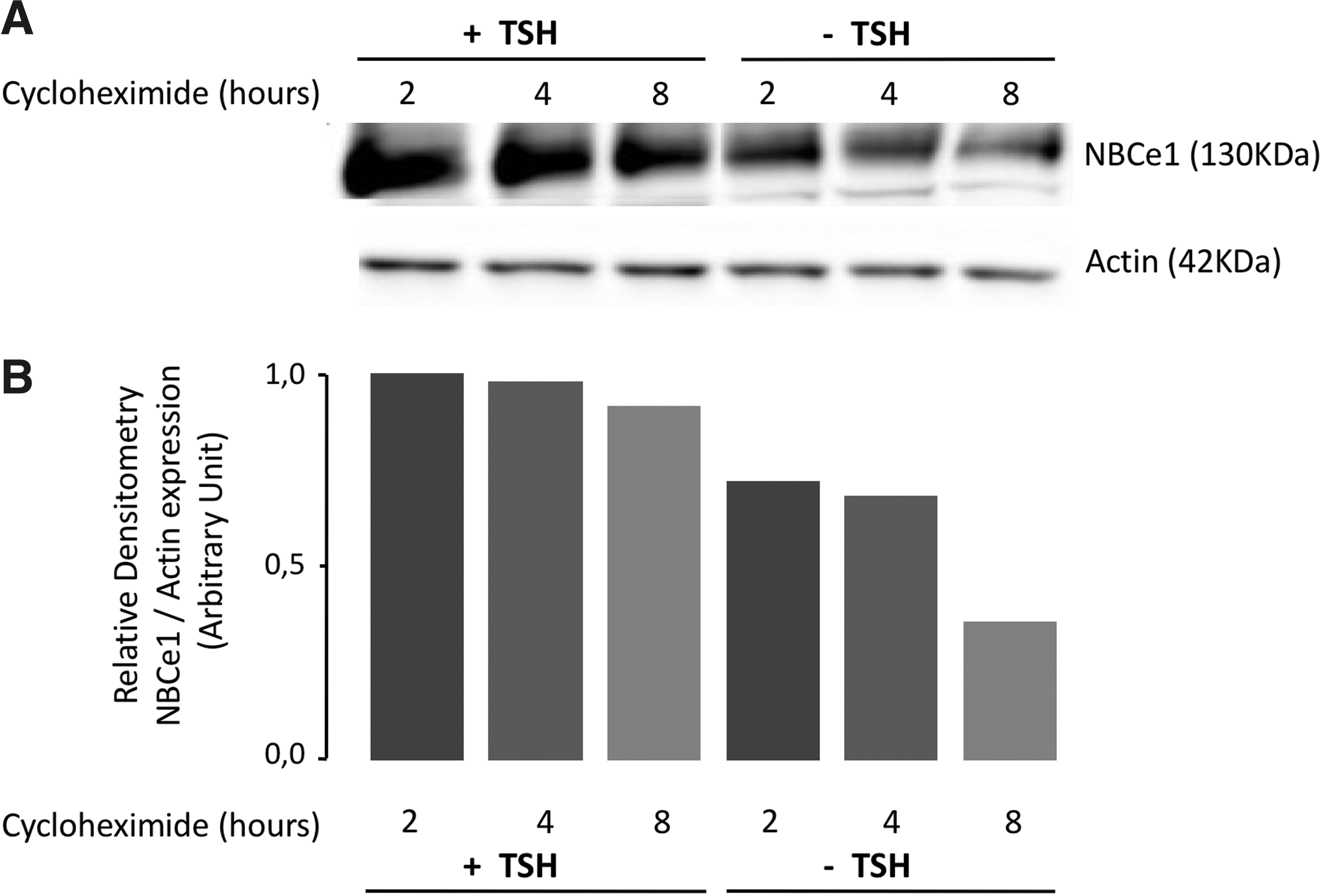
Effect of inhibition of protein synthesis on NBCe1 protein expression in PCCl3 monolayers cultured with or without TSH. Confluent PCCl3 monolayers were cultured for three days with or without TSH (1 mIU/mL) and then exposed to cycloheximide (100 μM) for up to 8 h to block protein synthesis. NBCe1 protein expression was quantified by Western blot analysis using the Abcam antibody
Hormonal modulation of NBCe1 protein degradation
Confluent PCCl3 monolayers cultured for three days with or without TSH (1 mIU/mL) were exposed to cycloheximide (100 μM) for up to 8 h to prevent protein synthesis. NBCe1 protein was quantified at 2, 4, and 8 h both with and without TSH. While there was almost no decrease in NBCe1 in the cells cultured with TSH (Fig. 7A and B), there was a clear decrease in NBCe1 protein when cells were cultured without TSH but exposed to cycloheximide. Based on two similar experiments, a rough estimate of the half-life was about 6 h. This indicates that TSH prolongs NBCe1 protein expression by impeding its degradation.
Discussion
This study demonstrates the expression of NBCe1 (SLC4A4) in human and rat thyroids. RT-PCR amplification of NBCe1 variants in rat and human thyroid cDNA revealed the presence of significant amounts of NBCe1-B transcripts only (Fig. 2). In rat thyroid PCCl3 cells, NBCe1-C was also amplified by primers specific for that variant (Fig. 2A, picture c). Additionally, using primers that simultaneously amplify NBCe1-B and NBCe1-C, the relative amount of each variant was estimated and was found to be lower for NBCe1-C (Fig. 2A, pictures b and c). In any case, no NBCe1-A was detected in the thyroid, while huge amounts of its mRNA were detected in the kidney. These results confirm the presence of NBCe1-B (and very small amounts of NBCe1-C) in rat but not human thyroid. Thus, NBCe1-B may be considered as the main, if not the only, isoform expressed in the thyroid. This may have functional implications. While the kidney isoform NBCe1-A mediates the exit of 1 Na+ with 3 HCO3 − at the basolateral membrane of the rat kidney proximal tubule, NBCe1-B, like NBCe1-C, mediates the entry of 1 Na+ with 2 HCO3 − in the rat exocrine pancreas and rat brain, respectively (1). The expression of NBCe1 protein within the thyroid was confirmed by immunofluorescence and Western blotting. For that analysis, commercial antibodies were used, which vary in their availability. The Chemicon antibody reacts with the A and B (but not C) isoforms, while the Abcam antibody does not discriminate between the different isoforms. As the presence of NBCe1-A transcripts was ruled out, the isoform detected with the Chemicon antibody corresponds to NBCe1-B. The cellular distribution of NBCe1 in rat and human thyroids demonstrated low, diffuse intracellular labeling in control conditions, whereas after TSH stimulation, intense basolateral membrane labeling was observed in both rat and human thyroids, as well as some intracellular vesicular labeling in human thyroids. However, it should be emphasized that the rats were treated for three weeks (as opposed to the human slices, which were treated for one day). Basolateral membrane localization of NBCe1 is quite usual in epithelia (19,20). At least in rat tissue, high TSH levels did not increase NBCe1 mRNA levels, thus suggesting that TSH does not increase NBCe1 synthesis but rather prevents its degradation by redirecting it to the basolateral membrane. The intracellular NBCe1 localization observed in human thyroid is probably the result of NBCe1 cotransporter trafficking to the basolateral membrane on TSH stimulation. Increased expression was also evident by Western blot analysis. Furthermore, experiments with cultured cells (primary human thyrocyte cultures and rat PCCl3 cells) showed that TSH and forskolin increased NBCe1-B expression two- to fourfold, probably due to exocytosis to the basolateral membrane. Similarly, cyclic adenosine monophosphate (cAMP) has previously been reported to stimulate the expression of NBCe1 and its basolateral membrane localization in pancreatic ducts with increased trans-epithelial bicarbonate secretion (21,22). The exact effect of protein kinase A (PKA) has not been examined in this context. In fact, a single consensus PKA phosphorylation site at Thr49 (KRKT49) has been described in the N-terminus of brain and pancreatic NBCe1-B (8,23). In murine colonic crypts, cholinergic and cAMP-dependent stimuli induce NBCe1 expression and translocation into the basolateral membrane (24), although both stimuli appear to require the protein kinase C pathway (24).
To investigate in vivo whether TSH also controls NBCe1 expression in the rat thyroid, a model of thyroid quiescence was generated by decreasing TSH levels in rats with T4 treatment, as well as a model of thyroid hyperactivation by increasing TSH levels with MMI and ClO4 − treatments added to their drinking water. The different effects of the T4, MMI, and ClO4 − treatments were reflected in the size and morphology of the thyroid glands of these rats, which were smaller than controls in the T4 group but two to three times larger in the MMI and ClO4 − groups. The lower level of TSH in the T4 group and higher level of TSH in the MMI and ClO4 − groups were confirmed compared to their matched controls. Using immunohistochemistry and Western blotting, the expression of NBCe1 as well as that of NIS and TPO was observed to follow TSH levels (25,26). Thus, TSH appears as a positive regulator of NBCe1 (basolateral membrane) expression as of other proteins important for thyroid function (27).
It is interesting that ClO4 −, which impairs iodide uptake by NIS and progressively depletes the thyroid of iodide, has a slower effect on TSH levels than MMI, which immediately blocks TPO and thyroid hormone synthesis. The effect of TSH on increased NBCe1 protein expression is not correlated with increased NBCe1 mRNA expression, at least not in rats exposed to MMI (Fig. 4). Similarly, TSH and forskolin stimulation in PCCl3 cells failed to increase NBCe1 mRNA levels (Fig. 6D) but did increase NBCe1 protein expression (Fig. 6B and C). Indeed, exposing PCCl3 cells to cycloheximide with or without TSH treatment to inhibit protein synthesis resulted in decreased NBCe1 expression only when TSH was removed (Fig. 7A and B), thus suggesting that TSH acts through a posttranscriptional mechanism, increased mRNA stability, increased translation, and inhibition of NBCe1 protein turnover, that is, inhibition of degradation or stabilization of the protein.
Similar data have been reported for several other membrane proteins, including ionic pumps such as rat lung Na+-K+-ATPase (28) and T-type Ca2+ channels in prostate cancer cells (29). The exact function of NBCe1 in thyroid physiology is currently unknown. It appears quite possible that basolateral NBCe1, present in both rat and human thyroid, participates in the control of cytosolic pH and possibly mediates the first step of trans-epithelial HCO3 − secretion toward the lumen, as in pancreatic ducts and the intestine. The pH of the intrafollicular space has been established at around 7.6 or higher after TSH treatment (30,31). In mice, this alkaline intrafollicular pH is critically dependent on the expression of pendrin (Slc26a4) in the apical membrane. Lack of pendrin expression results in acidic follicular lumen pH (31). Thus, the secretion of HCO3 − into the follicular lumen may rely on NBCe1 in the basolateral membrane and pendrin in the apical membrane. NBCe1-B is similarly expressed in the basolateral membrane of pancreatic and salivary ducts, corroborating its primary role of sustaining HCO3 − secretion induced by cAMP-mediated stimuli. For instance, basolateral NBCe1-B appears to be responsible for secretin-stimulated uptake of HCO3 − into pancreatic duct cells (21). In the human pancreas, the NBCe1-B variant is also exclusively observed in the basolateral membrane and mediates the entry of HCO3 − from the basolateral medium into the cell. Physiologically, the exit of HCO3 − from the cell has only been observed with the NBCe1-A variant and not with the B variant. As only the expression of the NBCe1-B variant was detected in the thyroid in this study, it therefore appears to ensure the entry of 1 Na+ with 2 HCO3 −. This should slightly hyperpolarize the basolateral membrane, which may be of importance for maintaining a favorable basolateral membrane potential for sustained activity of the NIS cotransporter, which itself mediates the entry of 2 Na+ with 1 I−, and so depolarizes the basolateral membrane potential. Thus, as TSH stimulates both cotransporters (NIS and NBCe1-B), their respective effects on the basolateral membrane potential may possibly cancel each other to such an extent that their abundance is similar. Yet, according to Purtell et al. (32), the KCNQ1-KCNE2 K+ channel provides the conductance for charge compensation whenever needed, as for Na+-K+-ATPase. These two membrane proteins appear to be essential for establishing the membrane potential. The exact relevance of inducing cytoplasmic alkalinization in thyroid physiology is still unknown. As previously mentioned, it may represent the first step in alkalinization of the follicular space, which facilitates iodide coupling to thyroglobulin, as well as binding and internalization of thyroglobulin (33,34). Alternatively, alkalinization of the thyrocyte cytoplasm may be required for the proliferative effect of TSH.
In summary, this study demonstrates that the thyroid expresses only the NBCe1-B isoform of NBC and that TSH strongly increases its expression and basolateral membrane insertion. The lack of increased transcription probably reflects an increase in NBCe1-B stability.
Footnotes
Acknowledgments
This work was supported by grants from the Dr. Jean-Pierre Naets and the Alphonse and Jean Forton funds, Brussels, Belgium, and the Belgian FRS-FNRS research fund.
Author Disclosure Statement
The authors have nothing to disclose.