
Abstract
Background:
Increasing evidence indicates that microRNA dysfunction is involved in the pathogenesis and progression of cancer. MicroRNA-222 (miR-222) is upregulated in papillary thyroid carcinoma (PTC). However, the role of miR-222 in invasion and metastasis of PTC remains unknown. This study investigated the function of miR-222 and its underlying mechanism in the progression of PTC.
Methods:
The expression of miR-222 was detected by quantitative reverse transcription polymerase chain reaction, and its correlation with various clinical characteristics was analyzed. The role of miR-222 in PTC cell migration ability was assessed with Transwell® assays and wound-healing assays in both TPC-1 and K1 cells. By using bioinformatics analyses and dual-luciferase 3′-UTR reporter assays, the study identified the direct target of miR-222 and the downstream pathways of PTC. Further, the study confirmed the role of miR-222 in promoting PTC distant metastasis in vivo by injecting TPC-1 cells into nude mice.
Results:
This study confirmed that miR-222 was upregulated in PTC tissues compared to adjacent thyroid tissues and that it correlated with aggressive cancer phenotypes. The results indicate that ectopic miR-222 enhanced cell migration and invasion of thyroid cancer cells in vitro and distant pulmonary metastases in vivo. Protein phosphatase 2 regulatory subunit B alpha (PPP2R2A), a tumor suppressor, was identified as a direct target of miR-222 through the 3′-UTR of PPP2R2A. Restoring PPP2R2A expression led to the attenuation of migration and invasion in miR-222-overexpressing thyroid cancer cells. Moreover, we found that miR-222 promoted invasion and metastasis partly through the AKT signaling pathway.
Conclusions:
Taken together, the results suggest that miR-222 promotes tumor invasion and metastasis in thyroid cancer by targeting PPP2R2A. Thus, miR-222 could serve as a potential diagnostic biomarker, as well as an attractive therapeutic tool for thyroid cancer.
Introduction
T
MicroRNAs (miRNAs) are a conserved class of small noncoding RNAs (approximately 22 nucleotides in length) that negatively regulate target gene expression (4). By binding to the 3′ untranslated region (3′-UTR) of target genes, miRNAs can induce mRNA degradation or repress mRNA translation (5). Recent studies have focused on the regulation of miRNA in tumorigenesis and progression by acting as oncogenes or tumor suppressors (6,7). MiRNAs contribute to the processes of carcinogenesis and metastasis through the regulation of cell proliferation, metabolism, apoptosis, and invasion (8,9). MiR-222-3p (indicated as miR-222 in this study) is a component of the miR-221/222 cluster, whose members are located on the X chromosome (Xp11.3) with identical seed sequences (10). MiR-222 is associated with the pathogenesis and progression of breast cancer and colorectal cancer (11 –13) and functions by influencing cell proliferation, tumor growth, and cell apoptosis (14,15). Previous reports found that miR-222 is upregulated in primary PTC tissues compared to normal thyroid tissues (16,17). Previous studies also showed overexpression of circulating miR-222 in PTC patients, and the level of miR-222 was well correlated with metastatic lymph node and advanced tumor, node, metastasis (TNM) stage, suggesting that miR-222 could be used as a biomarker for metastatic PTC (18). However, little is known regarding the specific mechanism of miR-222 in the metastasis of PTC.
The protein phosphatase 2 (PP2A) holoenzyme consists of a catalytic subunit, a scaffold subunit, and a regulatory subunit. The serine/threonine-protein phosphatase 2A 55 kDa regulatory subunit B alpha (PPP2R2A), a PP2A regulatory subunit, was suggested to be a tumor suppressor in many cancer types (19). It has been reported that PPP2R2A inactivates the downstream AKT pathway by suppressing AKT phosphorylation in several types of cancers (19,20), but little evidence exists regarding the role of PPP2R2A in PTC. The AKT pathway is well known for initiating several cancer cellular processes, including the cell cycle, cell proliferation, cell growth, and angiogenesis, and it is a key player in the tumorigenesis of various tumors, including PTC.
This study aimed to elucidate the biological functions of miR-222 and its role in regulating the PPP2R2A/AKT cascade in PTC.
Methods
Human tissue samples
Forty-two pairs of PTC tissues and corresponding adjacent thyroid tissues were obtained from patients enrolled at the First Affiliated Hospital of Sun Yat-sen University and Sun Yat-Sen University Cancer Center (Guangzhou, China), all of whom underwent surgery between December 2014 and December 2016. The tissues were immediately stored at −80°C until use. Clinicopathologic features, including age, sex, tumor size, multicentricity, extrathyroidal invasion, lymph node invasion, distant metastasis, and TNM stage, were also collected. The collection of tissue samples was obtained after receiving informed consent from all patients. All study procedures were approved by the Institutional Research Ethics Committee of Sun Yat-Sen University.
Cell lines and cell culture
The human thyroid epithelial cell line Nthy-ori 3-1 and the human PTC cell lines TPC-1 and BC-PAP were generous gifts from Professor Haixia Guan (China Medical University, Shenyang, China). The human PTC cell line K1 was obtained from the European Collection of Authenticated Cell Cultures (ECACC, Salisbury, United Kingdom). HEK293T cells was purchased from the American Type Culture Collection (ATCC, Manassas, VA). The human PTC cell lines and HEK293T cells were cultured in Dulbecco's modified Eagle's medium (DMEM; Invitrogen, Carlsbad, CA), and the Nthy-ori 3-1 cells were cultured in RPMI-1640 medium (Invitrogen). All media were supplemented with 10% fetal bovine serum (FBS; Gibco, Gaithersburg, MD). The cultures were routinely maintained at 37°C with 5% CO2 at 100% humidity.
Lentivirus construction and transfection
The recombinant lentivirus overexpressing miR-222, knockdown miR-222, and the negative control miRNA sequences (termed pLV3-miR-222, pLV3-miR-222-inhibitor, and pLV3-NC, respectively) were purchased from GenePhama (Shanghai, China). TPC-1 cells were transfected with the recombinant lentivirus plus 5 μg/mL of hexadimethrine bromide (Polybrene). To select stably transfected cells, the cells were re-suspended and cultured with puromycin (2 μg/mL) for two weeks. The transfection efficiencies were determined by quantitative reverse transcription polymerase chain reaction (qRT-PCR).
miRNA and plasmid transfections
The miR-222 mimics, inhibitor, and scrambled miRNA (negative control, N.C.) were all purchased from GenePharma (Shanghai, China). The sequence of the miR-222 mimic was: 5′-AGC UAC AUC UGG CUA CUG GGU-3′, and 3′-CCA GUA GCC AGA UGUA GCU UU-5′, and miR-222 inhibitor was 5′-ACC CAG UAG CCA GAU GUA GCU-3′. The sequence of the siRNA targeting PPP2R2A was: 5′-GAG GCC AGT CCA CGA AGA ATA-3′. PTC cells were seeded onto six-well plates and cultured for 24–48 h until 50–60% confluent, and then transiently transfected with corresponding miRNA and siRNA (all at 20 μM) using a Lipofectamine® 3000 Transfection Kit (Invitrogen) according to the manufacturer's instructions. After 48 h of miRNA transfection, the cells were harvested and used for further assays.
For PPP2R2A overexpression, the PPP2R2A cDNA sequence was cloned into the multiple cloning sites of the pcDNA3.1 vector (Invitrogen), and was designated as pCDNA3.1-PPP2R2A. Either pCDNA3.1-PPP2R2A or the control vector (empty pCDNA3.1) was co-transfected with miR-222 mimics or N.C. The transfection with plasmids was performed as described above.
RNA isolation and qRT-PCR analysis
Total RNA extractions from cell lines and frozen tissue samples were performed with TRIzol® reagent (Invitrogen) according to the manufacturer's instructions. To measure mRNA expression, a PrimeScript™ RT-PCR Kit (TaKaRa, Mountain View, CA) was used to synthesize cDNA from total RNA. To detect miRNA levels, a PrimeScript™ miRNA RT-PCR Kit (TaKaRa) was used for reverse transcription following the manufacturer's instructions. The qRT-PCR was then performed using SYBR Premix Ex Taq™ (TaKaRa) on a LightCycler 480 system (Roche Diagnostics, Pleasanton, CA). Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) and U6 were used as internal controls to normalize the expression levels of mRNAs and miRNAs, respectively. The primers used for amplification were designed by and purchased from Generay Biotech (Shanghai, China). The sequences of the primers were as follows: miR-222 forward, 5′-CGG GCA GCT ACAT CTG GCT-3′; reverse, Uni-miR qPCR primer (TaKaRa); U6 forward, 5′-ACG CAA ATT CGT GAA GCG TT-3′; reverse, Uni-miR qPCR primer (TaKaRa); PPP2R2A forward, 5′-AAA GGA ACT ATT CGG CTA TGT G-3′; reverse, 5′-AAA ATG ACC TGT TAC TGG GAT C-3′; GAPDH forward, 5′-GCA CCG TCA AGG CTG AGA AC-3′; and reverse, 5′-TGG TGA AGA CGC CAG TGG A-3′. Data were analyzed by the 2–ΔΔCT method.
Western blot analysis
The lysis of cell lines and frozen tissues were carried out using RIPA lysis buffer (#9806; Cell Signaling Technology, Danvers, MA) along with Halt™ Protease & Phosphatase Inhibitor Cocktail (Thermo Fisher Scientific, Santa Clara, CA). The protein concentration of each sample was measured by the BCA Protein Assay Kit (Pierce, Waltham, MA) and a TECAN Infinite M200 PRO microplate reader system (TECAN, San Jose, CA). Samples, each containing approximately 30 μg of protein, were separated by 10% sodium dodecyl sulfate polyacrylamide gel and transferred onto polyvinylidene fluoride membranes (EMB Millipore, Hayward, CA) using a wet transfer procedure. The membranes were blocked with blocking buffer (5% skim milk in TBS-T) at room temperature for one hour, then sequentially incubated with diluted primary antibodies in 5% bovine serum albumen at 4°C with gentle shaking overnight. The primary antibodies were as follows: mouse anti-human PPP2R2A (#5689; Cell Signaling Technology), rabbit anti-human AKT (#4691S; Cell Signaling Technology), rabbit anti-human phospho-AKT (Ser473; #4060S; Cell Signaling Technology), and rabbit anti-human GAPDH (Proteintech, Wuhan, China). After samples were washed with TBS-T, the membranes were incubated with appropriate secondary antibodies as follows: anti-rabbit immunoglobulin G (IgG), HRP-linked antibody (#7074; Cell Signaling Technology), or anti-mouse IgG, HRP-linked antibody (#7076; Cell Signaling Technology). The protein bands were detected using the SuperSignal West Pico Chemiluminescent Substrate (Thermo Fisher Scientific). The expression of GAPDH was used as a reference.
Dual luciferase 3′-UTR reporter assay
The vector for overexpression of human PPP2R2A 3′-UTR was purchased from GenePharma. The PPP2R2A 3′-UTR sequences containing the predicted miR-222 binding sites were inserted into the pGL3-basic vector (Promega, Madison, WI) at the XbaI restriction site immediately downstream of the luciferase gene to generate the firefly/Renilla luciferase reporter vector pGL3-basic-PPP2R2A-3′-UTR-WT. A seed-sequence transversion mutated version of the PPP2R2A 3′-UTR, named pGL3-basic-PPP2R2A-3′-UTR-Mut, was also constructed as a control (Fig. 3A).
HEK-293T cells were seeded onto 24-well plates and incubated for 24 hours. After reaching 50–60% confluency, the cells were co-transfected with the following combinations using Lipofectamine® 3000: miR-222 mimics and pGL3-basic-PPP2R2A-3′-UTR-WT; miR-222 mimics and pGL3-basic-PPP2R2A-3′-UTR-Mut; the miRNA negative control and pGL3-basic-PPP2R2A-3′-UTR-WT; and the miRNA negative control and pGL3-basic-PPP2R2A-3′-UTR-Mut. Additionally, all the groups were co-transfected with 10 ng of pRL-TK Renilla luciferase. Twenty-four hours after transfection, the cell lysates were collected and sequentially used for the detection of luciferase activity. Relative firefly luciferase activity, which was normalized to Renilla luciferase activity, was measured using a dual luciferase reporter gene assay system (Promega) according to the manufacturer's instructions.
Transwell® assay
Cell migration and invasion were evaluated by Transwell assays with or without Matrigel®. The procedure was described in a previous study (21). In summary, for the cell migration assays, a 24-well Boyden chamber (8.0 μm pore size; Corning, Corning, NY) with fibronectin (Roche CustomBiotech, Indianapolis, IN) was used. Forty-eight hours after transfection, TPC-1 or K1 cells (6 × 104) in 100 μL of serum-free DMEM was seeded into the upper chamber, while the lower chamber was filled with 600 μL of DMEM with 10% FBS. After incubation at 37°C with 5% CO2 for 24 hours, the upper chamber was cleaned with a cotton swab to remove the cells on the upper surface of the membrane. Afterwards, the migrated cells remaining on the bottom of the membrane were fixed with 4% paraformaldehyde and stained with 1% crystal violet. For cell invasion assays, a Matrigel-precoated Transwell chamber (Corning) was used instead, and the procedures were performed as previously described above. Images were obtained using a DMI4000B microscope (Leica, Wetzlar, Germany), and the number of cells was counted in five random fields (200 × ).
Wound-healing assays
Wound-healing assays were performed to assess cell migration using the Ibidi 2-Well Culture Insert system (Ibidi, Martinsried, Germany) according to the manufacturer's instructions. Briefly, after the insert was placed onto a 12-well plate, TPC-1 or K1 cells (6 × 104/mL) were applied into each well. At 50–60% confluency, the cells were transfected with exogenous miRNAs or plasmids and cultured at 37°C and 5% CO2 until 90–95% confluent. The insert was gently removed by using sterile tweezers, and the well was filled with serum-free DMEM. The cultures were incubated for 24 hours and photographed using an Axio Observer Z1 phase contrast microscope (Zeiss, Oberkochen, Germany).
Tumor metastasis assay in vivo
BALB/c nude mice (male, 5–6 weeks old) were purchased from Vital River (Beijing, China) and housed in the SPF barrier facilities of Sun Yat-Sen University. The randomized mice were divided into three groups—pLV3-miR-222, pLV3-miR-222-inhibitor, and pLV3-NC—and each group contained eight mice. One hundred microliters of transfected TPC-1 cell suspension (1 × 106 cells/mouse) was injected into the mice through the lateral tail vein. Six weeks after injection, the mice were sacrificed, and their lungs were excised, fixed, paraffin embedded, and sectioned consecutively (4 μm sections) for hematoxylin and eosin staining. All experiments involving animals were performed following the approved protocols of the Ethical Committee of Sun Yat-Sen University.
Statistical analysis
All experiments were performed independently at least three times. Data are presented as the mean ± standard deviation (SD) and were analyzed using GraphPad Prism v5.00 (GraphPad Software, Inc., La Jolla, CA). Student's t-test or one-way analysis of variance was used to determine statistical significance of differences between two groups or among variant groups, respectively. A p-value of <0.05 was considered statistically significant.
Results
Upregulation of miR-222 in human PTC tissues is associated with advanced stages in PTC patients
A previous study showed that serum miR-222 expression was significantly increased in PTC patients relative to benign tissues and healthy controls. In addition, higher serum miR-222 levels were found to be well correlated with lymph node metastasis and advance TNM stages (18).
To explore the role of miR-222 in PTC, the expression of miR-222 was further examined in 42 pairs of freshly frozen PTC tissues and corresponding adjacent thyroid tissues by qRT-PCR. The results showed that the miR-222 levels were significantly higher in PTC tissues than in their adjacent normal tissues (Fig. 1A). As shown in Figure 1B, increased miR-222 was found in 88% (37/42) of the PTC tissues with respect to the normal counterpart tissues. Then, a clinical analysis was performed to determine the clinicopathologic significance of miR-222 in PTC patients, and the results are summarized in Table 1. Consistent with previous findings, miR-222 expression was markedly elevated in the PTC tissues of patients with advanced TNM stages (Fig. 1C; p = 0.012). However, no significant correlation was observed between miR-222 expression and lymph node metastasis (p = 0.311). Interestingly, higher miR-222 levels were found to be significantly correlated with age and multicentricity (p = 0.012 and 0.002, respectively).
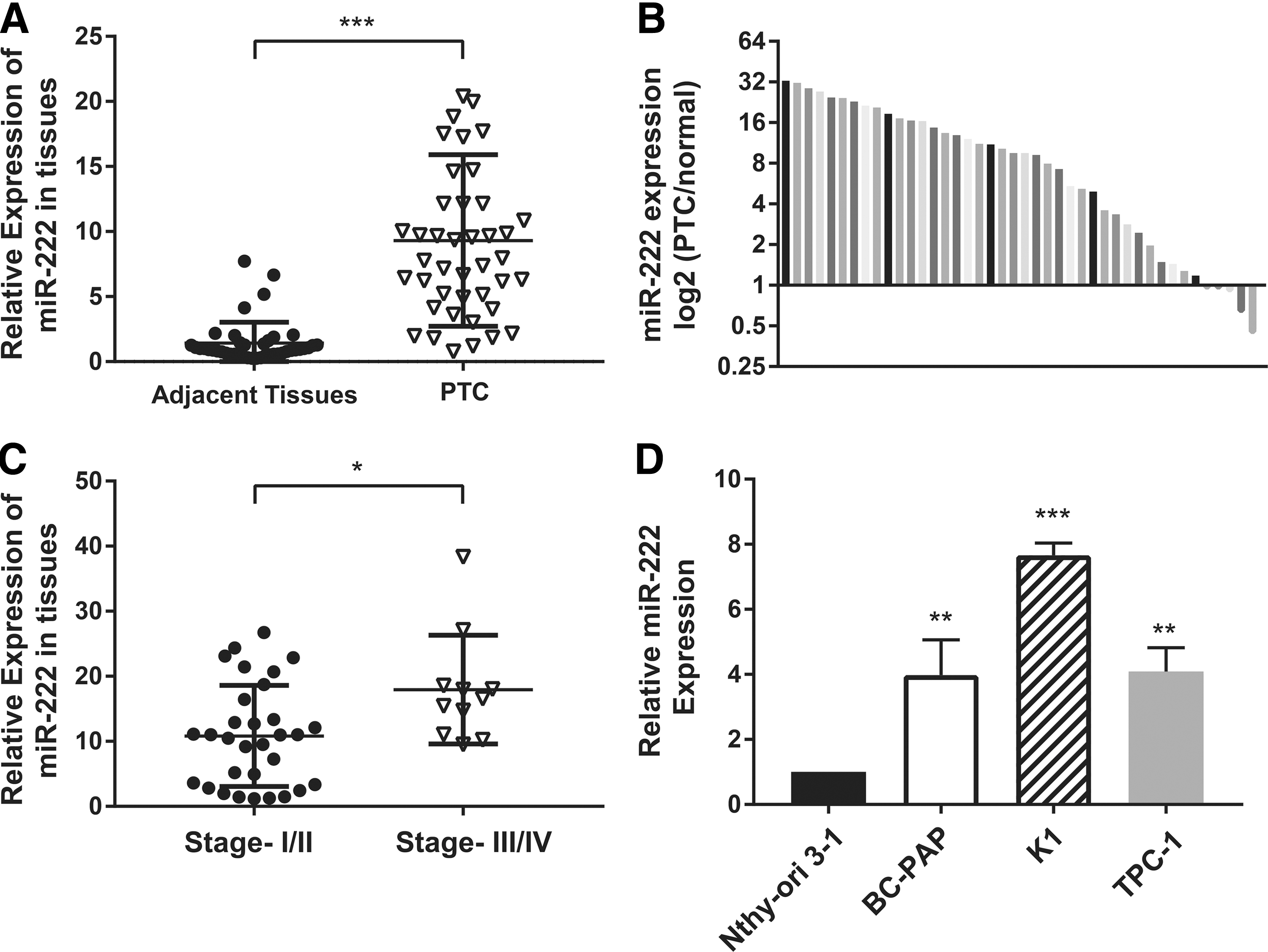
The expression of miR-222 is upregulated in papillary thyroid carcinoma (PTC) tissues and PTC cell lines. (
p < 0.05; ** p < 0.01.
PTC, papillary thyroid carcinoma; LN, lymph node; TNM, tumor, node, metastasis.
Moreover, miR-222 expression was determined in different PTC cell lines by qRT-PCR. The PTC cell lines, BC-PAP, K1, and TPC, exhibited higher levels of miR-222 than the normal thyroid epithelial cell line, Nthy-ori 3-1.
Thus, it was found that upregulation of miR-222 in human PTC tissues was closely associated with advanced TNM stages and might play an important role in PTC distant metastasis.
MiR-222 promotes PTC cell migration and invasion in vitro
To explore the biological functions of miR-222 in PTC progression, K1 cells with relatively higher levels of miR-222 and a BRAFV600E mutation, and TPC-1 cells, which carry the RET/PTC1 rearrangement and express relative lower levels of miR-222, were transiently transfected with miR-222 mimics, inhibitor, or the negative control oligonucleotides. Transwell assays without Matrigel demonstrated that overexpression of miR-222 promoted K1 and TPC-1 cell motility compared to the negative control groups. By contrast, the suppression of miR-222 exerted the reverse effect (Fig. 2A and B). Transwell assays with Matrigel showed that with ectopic miR-222, these two PTC cell lines presented dramatically enhanced invasiveness, while with the miR-222 inhibitor, they showed marked attenuation of invasion capacities (Fig. 2C and D).
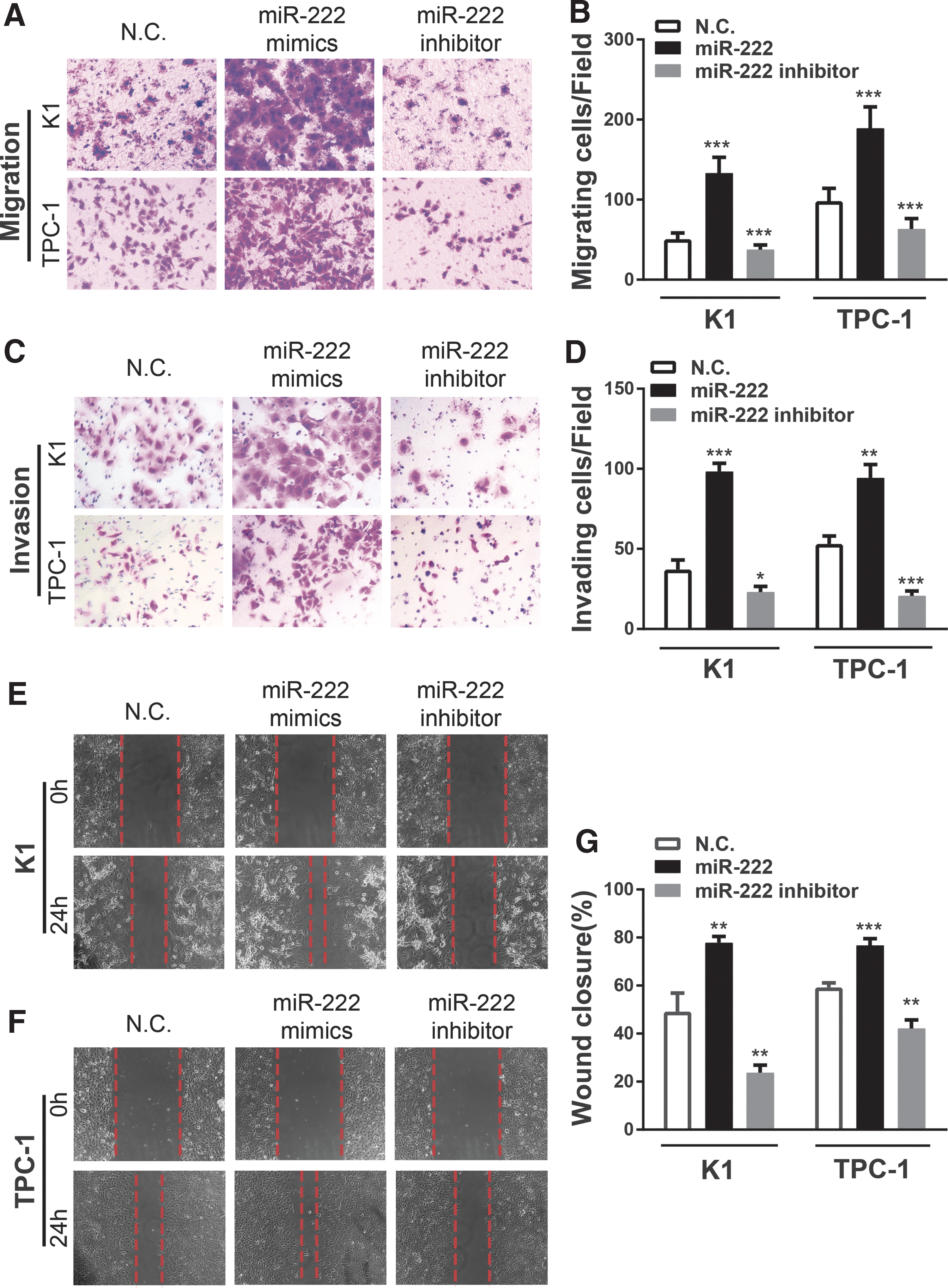
Overexpression of miR-222 promotes human PTC cell migration and invasion in vitro, while miR-222 knockdown exerts the reverse effects. (
Consistent with these observations, miR-222 increased the migration of K1 cells as observed with wound-healing assays (Fig. 2E). The wound closure of K1 cells after transfection with miR-222 mimics for 24 hours was dramatically elevated compared to the negative control cells. Similar findings were also observed in TPC-1 cells (Fig. 2F and G).
Collectively, the findings indicate that miR-222 significantly enhances the migration and invasion activity of PTC cells in vitro.
MiR-222 regulates PPP2R2A expression by directly targeting its 3′-UTR
To investigate the molecular mechanism for the promotion of migration and invasion by miR-222, potential targets were predicted using bioinformatics analyses. Three widely used prediction algorithms (miRecords, mirPath, and TargetScan) were adopted to screen for the candidate targets of miR-222 systematically. Among all the targets, PPP2R2A was identified as a putative miR-222 target based on the sequence at position 452–458 of its mRNA 3′-UTR fully complementary to the “seed region” of miR-222 (Fig. 3A).

PPP2R2A is a direct target of miR-222. (
To verify further whether the PPP2R2A transcript was the direct target of miR-222, luciferase reporter vectors with the 3′-UTR of PPP2R2A containing the predicted miR-222 binding site (pGL3-basic-PPP2R2A-3′-UTR-WT) or its corresponding mutation (pGL3-basic-PPP2R2A-3′-UTR-Mut) were constructed. For the dual luciferase assays, the wild-type or mutant vector was co-transfected into HEK293T cells with scrambled miRNA (N.C.) or miR-222 mimics, and pRL-TK luciferase reporters. MiR-222 overexpression only reduced relative luciferase activity with the wild-type vector. The mutant vector was unaffected by the change in miR-222 (Fig. 3B), indicating that miR-222 reduced PPP2R2A expression by directly binding to the potential specific sequence in the PPP2R2A 3′-UTR.
PPP2R2A expression was further analyzed in PTC cell lines and PTC tissues. As shown in Figure 3C, mRNA levels of PPP2R2A in PTC cell lines were remarkably reduced compared to the normal thyroid epithelial cell line, Nthy-ori 3-1. In addition, by using qRT-PCR, it was confirmed that endogenous mRNA levels of PPP2R2A were significantly downregulated in 42 PTC frozen tissues compared to their matched adjacent normal tissues (Fig. 3D). In three patients with elevated miR-222 levels in PTC tissues compared to their adjacent tissues, PPP2R2A protein levels were also measured. In the presence of higher miR-222 levels, the PPP2R2A protein expression was dramatically reduced in PTC tissues (Fig. 3E). Furthermore, as shown in Figure 3F, a significant inverse correlation was confirmed between miR-222 and PPP2R2A mRNA levels in the same set of PTC clinical tissues as Figure 1B.
Overall, these findings demonstrate that both the mRNA and protein expression levels of PPP2R2A are significantly decreased in PTC tissues when compared to the adjacent tissues.
MiR-222 activates the downstream tumor-associated genes of PPP2R2A and AKT
In addition, qRT-PCR and Western blotting assays were performed to evaluate PPP2R2A expression further in both miR-222-overexpressing and -reduced PTC cell lines. As shown in Figure 4A and B, the PPP2R2A mRNA levels were substantially decreased after ectopic overexpression of miR-222 in K1 and TPC-1 cells. Conversely, knockdown of miR-222 remarkably promoted PPP2R2A expression. As indicated in Figure 4C–I, p-AKT expression was significantly increased with miR-222 mimics and decreased with miR-222 inhibitor in both K1 and TPC-1 cells. These findings all suggest that miR-222 promotes the tumor-associated PPP2R2A/AKT signaling pathway in PTC.
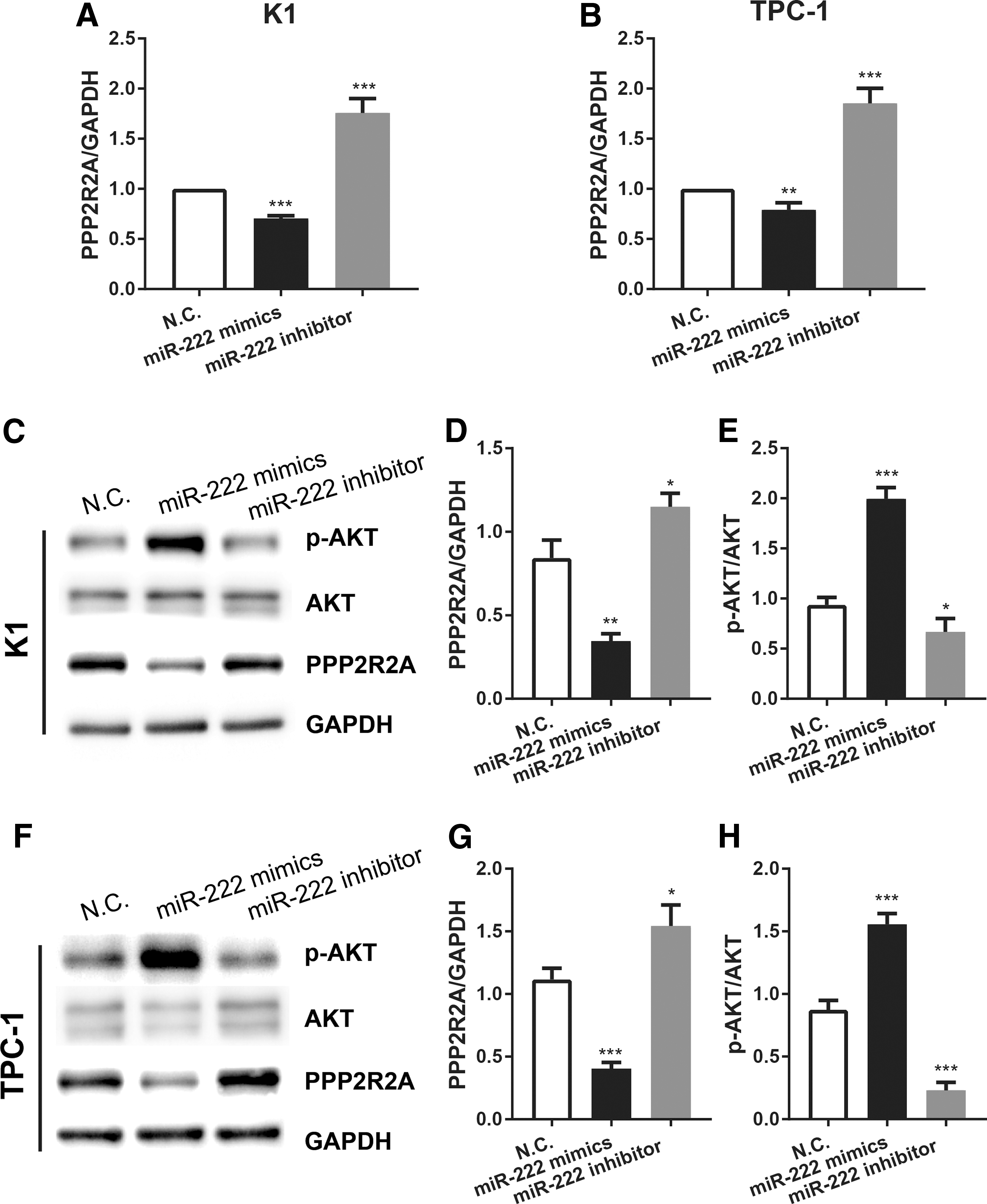
miR-222 activates the downstream the protein phosphatase 2 regulatory subunit B alpha (PPP2R2A)/AKT pathway in human PTC cell lines. (
PPP2R2A functions as a tumor suppressor in PTC
To explore the functional role of PPP2R2A further, a PPP2R2A siRNA (siPPP2R2A) was constructed to knock down its expression in PTC cells. As shown in Figure 5A and B, the mRNA and protein levels of PPP2R2A were significantly decreased after siPPP2R2A transfection. Meanwhile, the results show that silencing PPP2R2A expression promotes the phosphorylation of AKT, indicating that AKT is the downstream target of PPP2R2A in PTC cells. The biological impacts of PPP2R2A in PTC on cell migration and invasion were also assessed. A Transwell assay showed that after knocking down PPP2R2A, increased cell migration and invasion were observed both in K1 and TPC-1 cells (Fig. 5C–E), indicating that PPP2R2A serves as a potential tumor suppressor in PTC. This phenotype was similar to the miR-222-mimics-induced phenotype.

Knocking down PPP2R2A enhances PTC cell migration and invasion. (
Restoration of PPP2R2A expression reverses miR-222-mediated increases of migration and invasion of PTC cells
Since miR-222 significantly promoted PTC cell migration and invasion, and inhibited PPP2R2A expression by targeting its 3′-UTR, it is hypothesized that overexpression of PPP2R2A would reverse the phenotype of PTC cells associated with miR-222 expression. To verify this hypothesis, a plasmid containing the cDNA sequence of PPP2R2A mRNA (pCDNA3.1-PPP2R2A) was constructed. TPC-1 and K1 cells were co-transfected with scrambled RNA or miR-222 mimics and PPP2R2A vector or empty pCDNA3.1 vector. The protein levels of PPP2R2A, total AKT, and p-AKT were analyzed by Western blots. As shown in Figure 6A and B, PPP2R2A expression in PTC cells was strongly reduced after transfection with miR-222 mimics, whereas it was significantly elevated after co-transfection with PPP2R2A vector. Meanwhile, p-AKT expression was significantly increased after miR-222 mimic transfection, but could be reversed after PPP2R2A vector co-transfection.
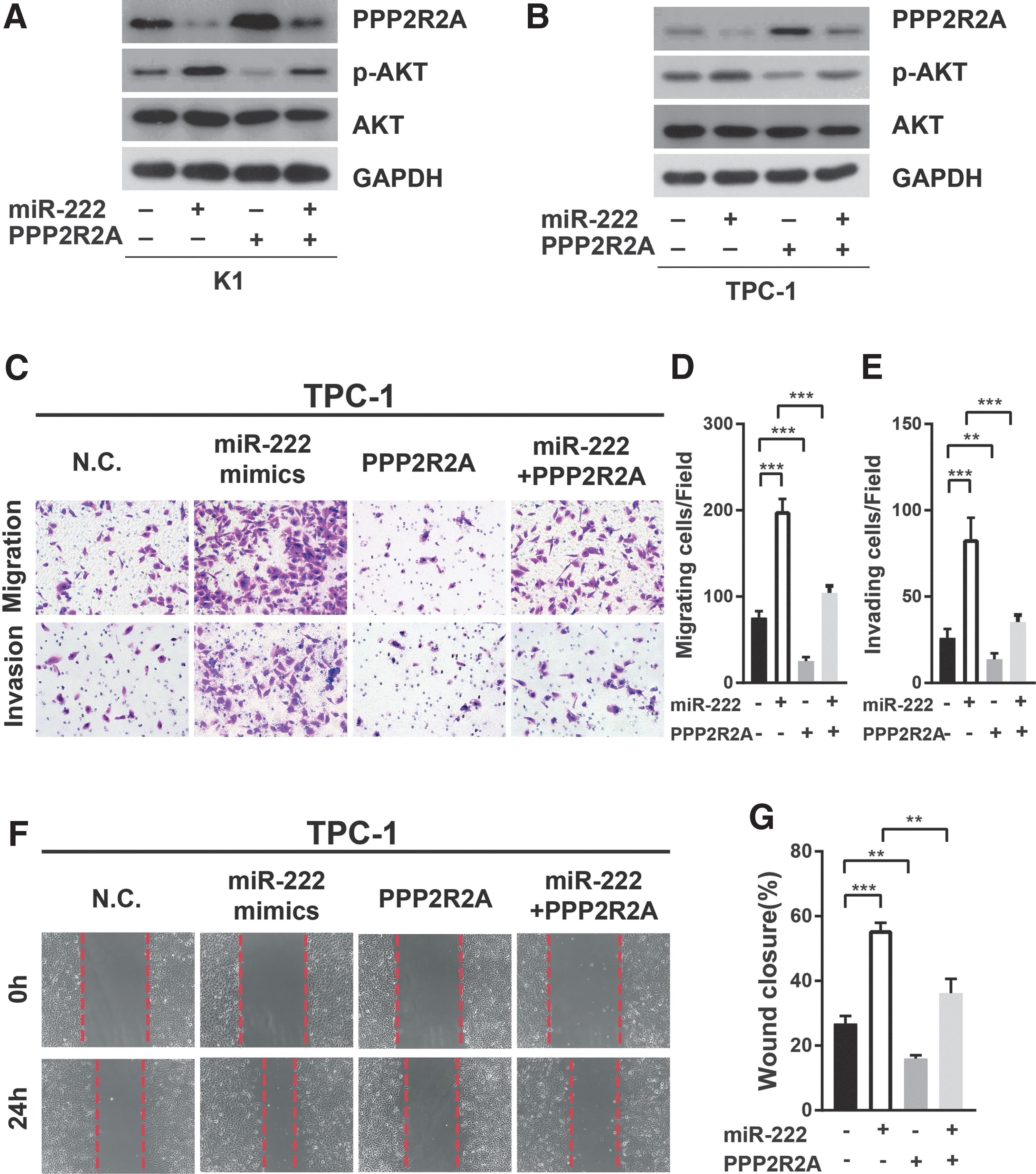
Protein phosphatase 2 regulatory subunit B alpha (PPP2R2A) is involved in miR-222-mediated cell migration and invasion. (
Functional studies, including Transwell assays and wound-healing assays, demonstrated that the effects of miR-222 on promoting migration and invasion of TPC-1 cells could be abrogated by the restoration of PPP2R2A expression (Fig. 6C–G).
Taken together, these results support the hypothesis that miR-222 enhances migration and invasion in thyroid cancer, at least partially by targeting PPP2R2A.
MiR-222 enhances in vivo lung metastasis
To evaluate whether miR-222 enhances tumor metastasis in vivo, TPC-1 cells were stably transfected with three different lentivirus vectors (pLV3-miR-222, pLV3-miR-222-inhibitor, and pLV3-NC), and separately inoculated into the tail veins of BALB/c nude mice. Mice were sacrificed eight weeks post injection, and their lungs were fixed for further investigations.
Strikingly, compared to the negative control group, mice injected with TPC-1 cells overexpressing miR-222 displayed a higher incidence of lung metastasis (Fig. 7A). Macro- and microscopic histological analyses confirmed a significant increase of tumor nodules in the lungs (Fig. 7B–E). Meanwhile, incidence of lung metastasis and the number of tumor nodules were dramatically reduced in TPC-1 cells with miR-222 inhibitor versus the control cells, suggesting that the metastatic potential of PTC cell lines is promoted by forced miR-222 expression and diminished by the impaired miR-222 expression. Furthermore, immunohistochemistry of PPP2R2A and p-AKT in the lung tumor nodules was conducted. Mice injected with TPC-1 cells with miR-222 mimics showed a significant lower expression of PPP2R2A and a lower level of p-AKT in the lung tumor nodules versus the control group, while tumor nodules formed by TPC-1 cells with inhibition of miR-222 displayed a higher expression of PPP2R2A and lower levels of p-AKT (Fig. 7F–H). Overall, miR-222 appears to play an essential role in the metastatic ability of PTC cell lines through altered activation of the PPP2R2A/AKT pathway in nude mice in vivo.
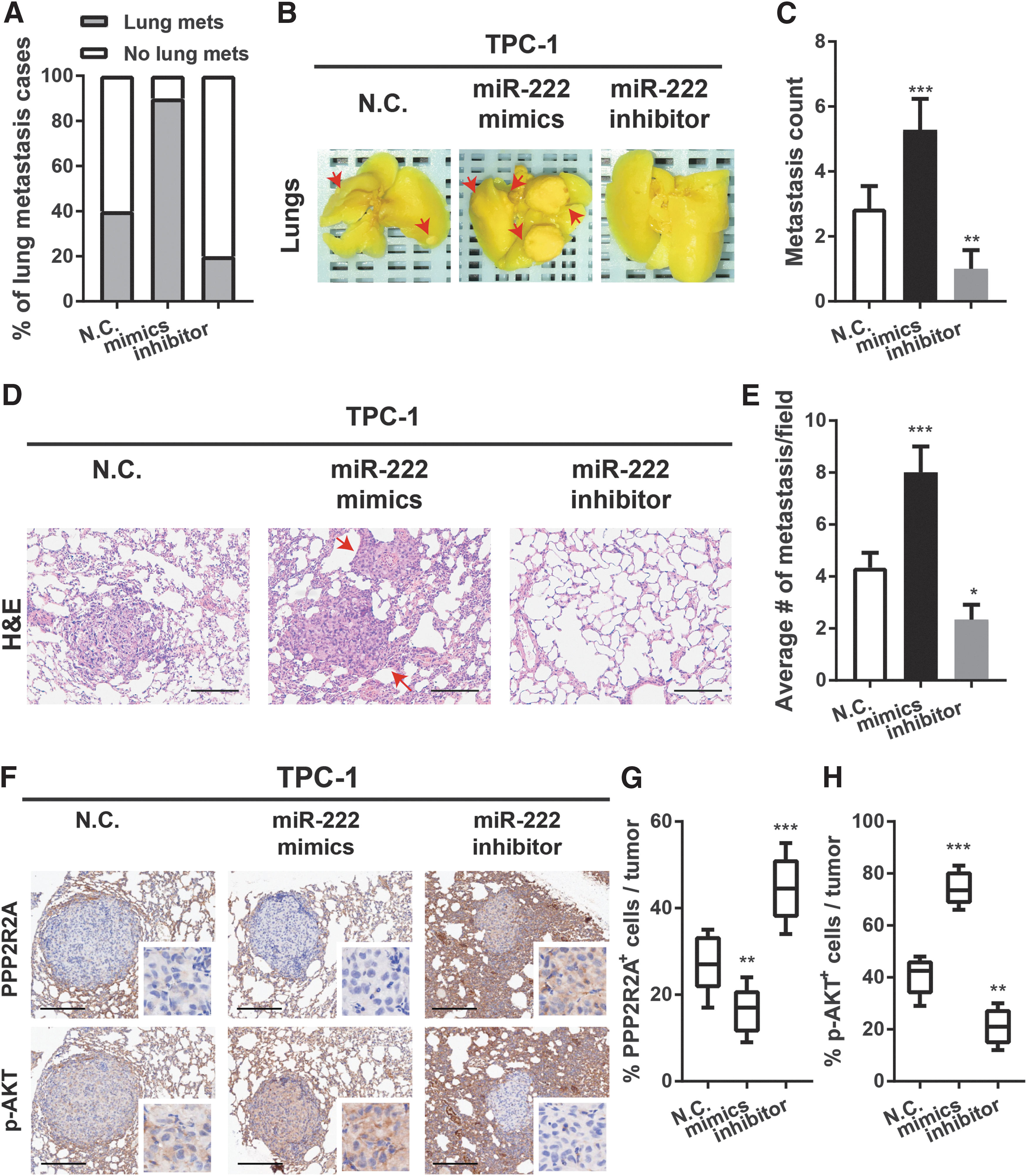
MiR-222 enhances PTC metastases into the lungs in vivo. (
Discussion
It is well established that miRNAs play essential roles in cancer pathogenesis and progression. MiRNAs contribute to the processes of carcinogenesis and metastasis by regulating cell proliferation, metabolism, apoptosis, and invasion (8,9). MiR-222 is widely known for its upregulation in many cancer types, including breast cancer (14), colorectal cancer (15), and pancreatic cancer (22). However, the role of miR-222 in invasion and metastasis of thyroid cancer remains unknown.
This study confirms the upregulation of miR-222 in PTC tissues compared to adjacent thyroid tissues, as well as its correlation with aggressive cancer phenotypes. Increased miR-222 expression enhances cell migration and invasion of thyroid cancer cells in vitro and distant pulmonary metastases in vivo. Decreasing tumor suppressor PPP2R2A levels may be a mechanism whereby miR-222 promotes invasion and metastasis in thyroid cancer. MiR-222 has been identified as an oncogene, and studies have shown that miR-222 is closely associated with cell proliferation and in promoting invasion and metastasis. In pancreatic cancer (22), overexpression of miR-222 dramatically enhanced cell proliferation and invasion by downregulating its target gene TIMP-2. It has also been reported that increased miR-222 expression caused by Helicobacter pylori infection promotes gastric cancer cell growth and invasion by suppressing RECK expression (23). Previous studies demonstrated that miR-222 was frequently overexpressed both in serum and tumor tissues in PTC patients and was associated with tumor aggressiveness and recurrence (24,25). Previous studies also found a positive correlation between circulating miR-222 levels and metastatic PTC, suggesting that miR-222 plays a role in tumor invasion and metastasis in PTC (18). However, the direct targets of miR-222 and its biological functions in PTC metastasis remain poorly understood. The present study shows that miR-222 is commonly upregulated in PTC tissues and that it is correlated with advanced stages of cancer. It also demonstrates that miR-222 promotes tumor cell migration and invasion, but not the proliferation in vitro, and the development of distant pulmonary metastasis in vivo. These findings indicate that miR-222 plays an essential role in thyroid cancer invasion and metastasis.
Regarding its mechanism, previous studies have shown that miR-222 exerts its function as a tumor metastasis promoter by targeting HAI-1, GNAI3, and ERα (23,26,27). In thyroid cancer, miR-222 induces cell-cycle arrest through negative regulation of its target p27 (28). This study confirm that miR-222 directly regulates PPP2R2A via binding to its mRNA 3′-UTR. PP2A is a serine-threonine protein phosphatase consisting of three subunits: a catalytic subunit (C), a scaffolding subunit (A), and a regulatory B subunit. Different B subunits lead to distinct substrate specificity and subcellular localization (29). PPP2R2A, also referred to as PP2A regulatory B subunit B55α, has been well-established as a master regulator of the cell cycle (19,30). However, its role in tumor metastasis remains less investigated. PPP2R2A was reported to play a role in tumor invasion and metastasis in pancreatic cancer (31), while others have reported (32) that PPP2R2A enhances the binding of c-Jun to genes involved in cell migration and invasion, resulting in elevated cell migration. This study demonstrates the downregulation of PPP2R2A expression in PTC tissues and cell lines and a reverse correlation between PPP2R2A and miR-222 levels. Further, it confirms the role of PPP2R2A as a tumor suppressor in thyroid cancer by suppressing or overexpressing it. While inhibiting PPP2R2A promotes cell migration and invasion, forced expression of PPP2R2A suppressed the migration and invasion capacities. Because miR-222 directly targets and inhibits PPP2R2A expression, further gain-of function assays demonstrate that higher levels of PPP2R2A attenuate the miR-222-induced cell motility and invasion, suggesting that the miR-222/PPP2R2A pathway plays a critical role in the regulation of thyroid cancer progression. To the authors' knowledge, this is the first study reporting PPP2R2A as a regulator in cancer invasion and metastasis in thyroid cancer.
To date, little is known about the underlying molecular mechanism of the miR-222/PPP2R2A axis in metastasis in thyroid cancer. Several studies have demonstrated a negative correlation between PPP2R2A and AKT phosphorylation in cancer. The AKT pathway is well known for initiating several cellular cancer processes, including alterations of the cell cycle, cell proliferation, cell growth, and angiogenesis (33 –35). Shouse et al. (19) reported that loss-of-function mutations of PPP2R2A induce activation of the AKT pathway in acute myeloid leukemia patients. A recent study also indicated that in pancreatic cancer, the PITX2 protein promoted PPP2R2A expression by binding to its promoter, further inhibiting the phosphorylation of AKT (20). Additionally, different AKT isoforms perform distinct functions in tumor progression (36), and generally AKT is phosphorylated and activated in tumor metastasis. Consistent with these findings, the present results show that forced expression of miR-222 in PTC cell lines downregulate PPP2R2A expression and enhance phosphorylation of AKT, while inhibiting miR-222 expression induces the opposite effect. This finding indicates that miR-222 promotes invasion and metastasis by regulating the PPP2R2A/AKT pathway.
Preclinical and clinical studies have demonstrated that miRNA targeted therapies using miRNA mimics and inhibitors (anti-miRs) may become novel therapeutic approaches for treating diseases (37,38). The present results demonstrate that miR-222 inhibition suppresses the metastasis ability of PTC cell lines in mouse models. Therefore, exogenous anti-miR-222 might serve as a novel therapeutic strategy in treating thyroid cancer, especially aggressive thyroid cancer. However, miRNA-based therapy requires further studies to validate this strategy.
Taken together, the current studies suggest that upregulation of miR-222 promotes tumor invasion and metastasis of thyroid cancer in vivo and in vitro. MiR-222 regulates the PPP2R2A/AKT pathway, suggesting that miR-222 could serve as a biomarker and potential therapeutic target for PTC patients.
Footnotes
Acknowledgments
We thank Professor Haixia Guan (The First Affiliated Hospital of China Medical University) for his generous gifts TPC-1, BC-PAP and Nthy-ori 3-1 cells. This research was supported by grants from the National Natural Science Foundation of China (no. 81272932), the Finance Department Foundation of Guangdong Province (no. 20160902), and the Medical Scientific Research Foundation of Guangdong Province (no. A2016065).
Author Disclosure Statement
None declared. No competing financial interest exists.