
Abstract

A recent publication by Tanaka et al. described the spontaneous development of Sjögren's syndrome in C57BL/6 mice with a conditional knockout of the Satb1 gene in hematopoietic cells (1). Satb1 (special AT-rich sequence binding protein) is a genome organizer involved in establishing immune tolerance by impairing negative selection of T cells in the thymus and reducing the efficacy of regulatory T cells. Not mentioned by Tanaka et al. (1) is that Sjögren's syndrome also develops in NOD.H2h4 mice that spontaneously develop autoimmune thyroiditis [reviewed by Braley-Mullen and Yu (2)]. We wish to point out that our recent genetic analysis of NOD.H2h4 mice revealed that a locus associated with thyroid autoimmunity contains the Satb1 gene (3).
NOD.H2h4 mice were derived by substituting the major histocompatibility class (MHC) molecule I-Ak on chromosome 17 from B10.A4R mice for I-Ag7 in diabetes-prone nonobese diabetic (NOD) mice [reviewed in Braley-Mullen and Yu (2)). Instead of developing type 1 diabetes, NOD.H2h4 mice develop thyroiditis in association with autoantibodies to thyroglobulin and thyroid peroxidase. In searching for the genetic basis for these traits, we observed highly significant linkage for thyroglobulin antibodies (LOD scores >4) to a locus on chromosome 17 (3). This locus includes portions from the B10.A4R (27.3–44.8 Mb) and NOD.H2h4 (44.8–75.96 Mb) strains, the former containing MHC region genes (Fig. 1).
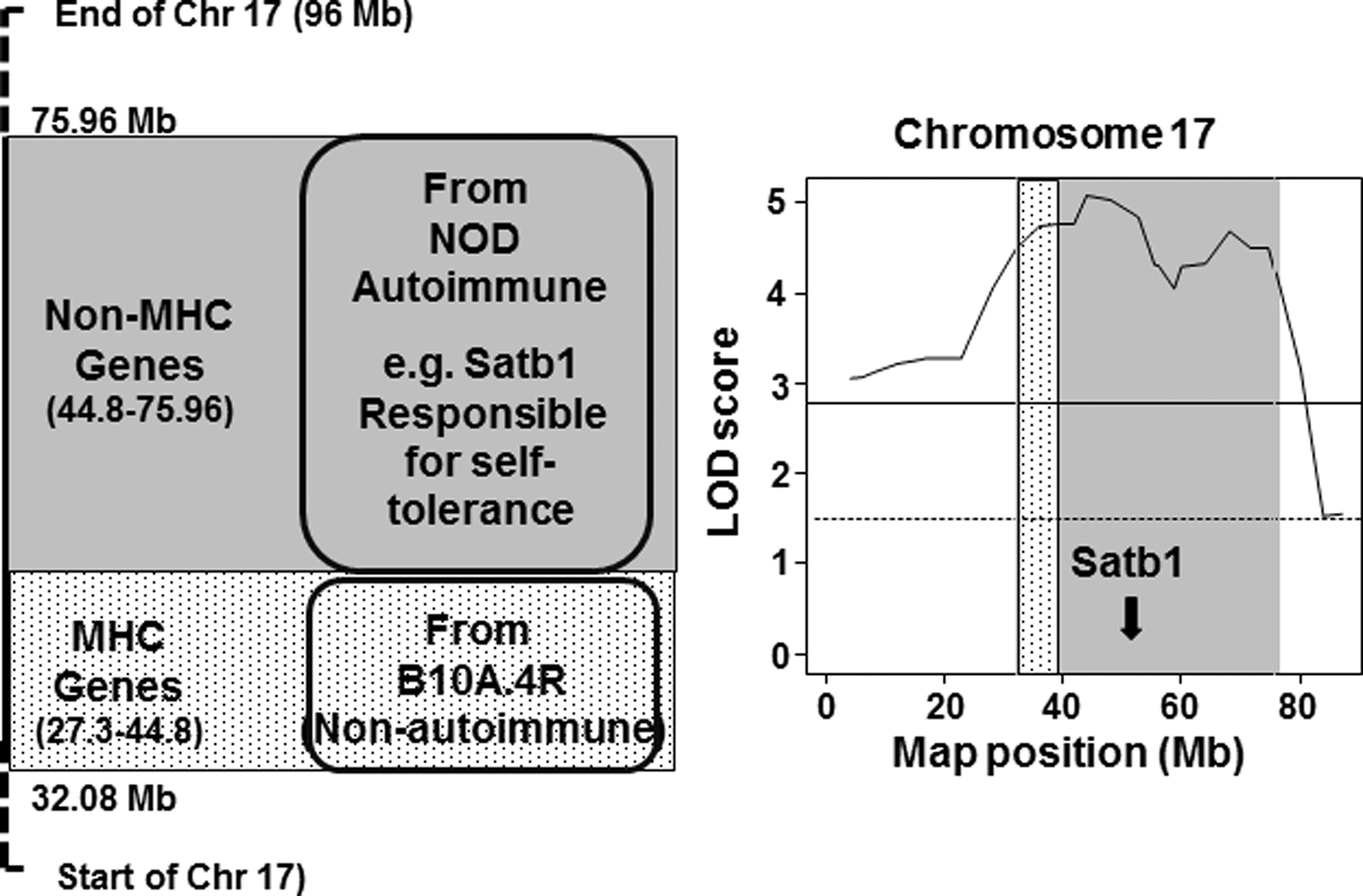
Left panel: Schematic illustration of the chromosome 17 locus linked to TgAb in NOD.H2h4 mice and its origin from the autoimmune NOD mouse strain (gray shading) and the nonautoimmune B10A.4R strain (speckled shading). Right panel: Linkage analysis (high resolution map) for this Chromosome 17 locus including the grey and speckled shading to emphasize the origin of genes from the NOD and B10.A4R strains [adapted with permission from McLachlan SM et al. 2017 (3)].
Because there was also strong linkage to adjacent non-MHC region genes from the NOD.H2h4 mice, we reviewed the genes within this region (3) and noted the presence of Satb1 at 51.9 Mb (Fig. 1), a gene with a role (as mentioned above) in immune tolerance. We therefore speculated that polymorphisms in genes like Satb1 could contribute to the loss of immune tolerance to thyroid autoantigens and the development of thyroid autoimmunity in NOD.H2h4 mice (3). In addition to Sjögren's syndrome in conditional Satb1 knockout mice (1), lymphocytic infiltrates were previously observed in lung, liver, and stomach. However, thyroid tissue does not appear to have been examined, and it would be of interest to know if autoantibodies to thyroid autoantigens arise in these animals.
With the new information that knockout of Satb1 in C57BL/6 mice leads to multi-organ autoimmunity, if Satb1 contributes to thyroid autoimmunity in NOD.H2h4 mice, then Satb1 is either nonfunctional or has a variable phenotype that may also lead to Sjögren's syndrome. In mice with the conditional Satb1 knockout, Sjögren's syndrome develops rapidly being apparent in 4 week old mice, whereas in the NOD.H2h4 strain, Sjögren's syndrome is rare in 8 week old mice and the symptoms are most evident in 1 year old females (2). This difference in time course is consistent with the total absence of Satb1 in the knockout strain versus the possibility of a dysfunctional Satb1 protein in NOD.H2h4 mice.
Turning to autoimmune thyroid disease in humans, despite evidence for a contribution by regulatory T cells, the major susceptibility genes identified do not include SATB1, which is located on chromosome 3. Only two thyroid autoimmunity susceptibility genes in humans are located on chromosome 3. First, LPP (LIM domain containing preferred translocation partner in lipoma) was included in seven newly identified loci for thyroid autoimmunity in a United Kingdom population (4). Its location on chromosome 3 (188,153,050..188,890,671 Mb) is upstream of SATB1 (18,345,388..18,445,588 Mb). The second thyroid autoimmunity susceptibility gene on chromosome 3, KALRN (kalirin RhoGEF kinase), linked to development of thyroid peroxidase autoantibodies (5), is also located in a different chromosome 3 region (124,033,341..124,726,325 Mb). Nevertheless, because of the similarity of the NOD.H2h4 thyroiditis model to human Hashimoto's disease, re-examining SATB1 as a candidate gene for thyroid autoimmunity may be of value.
Footnotes
Acknowledgments
This work was supported by the National Institutes of Health Grants DK54684 (S.M.M.)
Author Disclosure Statement
No competing financial interests exist.