
Abstract
Background:
Loss of function mutations in the thyroid hormone (TH)-specific cell membrane transporter, the monocarboxylate transporter 8 (MCT8), lead to profound psychomotor retardation and abnormal TH serum levels, with low thyroxine (T4) and high triiodothyronine (T3). Several studies point to impaired TH transport across brain barriers as a crucial pathophysiological mechanism resulting in cerebral hypothyroidism. Treatment options for MCT8-deficient patients are limited and are focused on overcoming the brain barriers. The aim of this study was to evaluate the ability of the TH analog sobetirome and its prodrug Sob-AM2 to access the brain and exert thyromimetic actions in the absence of Mct8.
Methods:
Juvenile wild-type (Wt) mice and mice lacking Mct8 and deiodinase type 2 (Mct8/Dio2KO) were treated systemically with daily injections of vehicle, 1 mg of sobetirome/kg body weight/day, or 0.3 mg of Sob-AM2/kg body weight/day for seven days. Sobetirome content was measured using liquid chromatography–tandem mass spectrometry, and T4 and T3 levels by specific radioimmunoassays. The effect of sobetirome treatment in the expression of T3-dependent genes was measured in the heart, liver, and cerebral cortex by real-time polymerase chain reaction.
Results:
Sob-AM2 treatment in Mct8/Dio2KO animals led to 1.8-fold more sobetirome content in the brain and 2.5-fold less in plasma in comparison to the treatment with the parent drug sobetirome. Both sobetirome and Sob-AM2 treatments in Mct8/Dio2KO mice greatly decreased plasma T4 and T3 levels. Dio1 and Ucp2 gene expression was altered in the liver of Mct8/Dio2KO mice and was not affected by the treatments. In the heart, Hcn2 but not Atp2a2 expression was increased after treatment with the analogs. Interestingly, both sobetirome and Sob-AM2 treatments increased the expression of several T3-dependent genes in the brain such as Hr, Abcd2, Mme, and Flywch2 in Mct8/Dio2KO mice.
Conclusions:
Sobetirome and its amide prodrug Sob-AM2 can access the brain in the absence of Mct8 and exert thyromimetic actions modulating the expression of T3-dependent genes. At the peripheral level, the administration of these TH analogs results in the depletion of circulating T4 and T3. Therefore, sobetirome and Sob-AM2 have the potential to address the cerebral hypothyroidism and the peripheral hyperthyroidism characteristic of MCT8 deficiency.
Introduction
L
Treatment options for MCT8-deficient patients are limited. Because patients present cerebral hypothyroidism in combination with peripheral hyperthyroidism, the design of an appropriate treatment is challenging. Therapeutic strategies should improve the neurological defects derived from TH deficiency in the brain and alleviate the peripheral thyrotoxicosis. The available treatments consist of a combination of levothyroxine (LT4) and propylthiouracil (PTU) (20), or the TH analog 3,5-diiodothyropropionic acid (DITPA) (21). Both experimental treatments normalize T3 levels in plasma and peripheral tissues, although without improvement to neurological impairment. The ability of the TH analog triiodothyroacetic acid (TRIAC) to ameliorate the peripheral thyrotoxicosis is currently under investigation (
Sobetirome is such a thyromimetic that may provide benefit in MCT8 deficiency. Sobetirome is a potent T3 mimic that binds to the TH receptors with subtype selectivity for thyroid hormone receptor beta (TRβ) over thyroid hormone receptor alpha (TRα) (31,32). In addition, sobetirome has the ability to cross the BBB and distribute to the CNS from a systemic dose at levels sufficient to upregulate T3 target genes, modulate brain lipid biomarkers, and affect brain developmental pathways regulated by T3 (33
–36). Sobetirome is a clinical stage molecule that has progressed into clinical trials for hyperlipidemia (37) and is currently in clinical development for X-linked adrenoleukodystrophy, an inborn error of metabolism affecting the CNS and adrenals (
The aim of this study was to evaluate the ability of sobetirome and Sob-AM2 to access the brain in the absence of Mct8 by directly measuring sobetirome levels in the brain and by analyzing the expression of T3-dependent genes following systemic administration in mice. The absence of Mct8 in mice does not lead to gross neurological abnormalities (41) because, unlike humans, mice abundantly express the organic anion transporting polypeptide 1c1 (Oatp1c1) at the BBB that mediates T4 transport into the brain (42 –44). Deiodinase type 2 (Dio2) activity is increased in the brain of single Mct8 knockout animals (Mct8KO); Dio2 converts the T4 available through the Oatp1c1 into T3, compensating for the lack of Mct8 (17,18,45). Deletion of both Mct8 and Dio2 in mice leads to a 50% reduction in the cerebral T3 content (46), similar to what has been found in the cerebral cortex of a 30th gestational week fetus with mutations in MCT8 (19) and presents a gene expression pattern in the brain very similar to that of hypothyroid animals (45). Moreover, these mice replicate the peripheral hyperthyroidism characteristic of MCT8-deficient patients (46).
In this work, Mct8/Dio2 KO juvenile mice were treated systemically with sobetirome or Sob-AM2, and the results indicate that these TH analogs do not depend on Mct8 for their CNS distribution and that they are able to stimulate expression of TH target genes in the brain in the absence of Mct8. Moreover, it was found that treatment with sobetirome and Sob-AM2 decreases the peripheral hyperthyroidism characteristic of Mct8 loss of function. All these data suggest that sobetirome and Sob-AM2 may provide benefit in MCT8 deficiency.
Materials and Methods
Reagents
Drugs were prepared at concentrations suitable for intraperitoneal (i.p.) injection of 5 μL/g body weight. Sobetirome (MW = 328 g/mol) and Sob-AM2 (MW = 341 g/mol) were synthesized at the Scanlan Laboratory, as previously described (31,40,47). Drug stocks were prepared by dissolving sobetirome and Sob-AM2 at 1 mg/mL in dimethyl sulfoxide (DMSO). The 1 mg/mL drug stocks were diluted in saline to obtain final solutions of 0.2 mg/mL of sobetirome and 0.06 mg/mL of Sob-AM2 in 20% DMSO (corresponding to 1.0 and 0.3 mg/kg dose, respectively). Vehicles were prepared with 20% DMSO in saline.
Ethics statement
All experimental procedures involving animals were performed following the European Union Council guidelines (directive 2010/63/UE) and Spanish regulations (R.D. 53/2013), and were approved by the ethics committees at Consejo Superior de Investigaciones Científicas (CSIC) and Community of Madrid (approval number 162/17). All efforts were made to minimize suffering, as indicated below.
Animal models and experimental design
All mice were housed at the Instituto de Investigaciones Biomédicas “Alberto Sols” under temperature- and light-controlled conditions at 22 ± 2°C on a 12:12 light–dark cycle with ad libitum access to food and water. Mct8−/y (Mct8KO) mice were originally produced by Dumitrescu et al. (17), Dio2 −/− (Dio2KO) mice were initially produced by Schneider et al. (48), and a colony was established at the animal facility in the same C57BL/6J genetic background. Mct8/Dio2KO were generated in the same C57BL/6J genetic background by mating Mct8+/y Dio2−/− males and Mct8−/+ Dio2−/− females. Experiments were carried out in juvenile male Mct8/Dio2KO mice and male wild-type (Wt) C57BL/6J mice. The Mct8 and Dio2 genotypes were confirmed by polymerase chain reaction (PCR) of tail DNA, as described (14).
On postnatal day 30 (P30), Mct8/Dio2KO mice were treated for seven days with daily i.p. injections of vehicle (n = 6), sobetirome (1.0 mg/kg body weight; n = 6), or Sob-AM2 (0.3 mg/kg body weight; n = 6). Wt mice were also treated for seven days on P30 with daily i.p. injections of vehicle (n = 8). Mice were euthanized four to six hours after the last i.p. injection. Mice were anesthetized with ketamine (75 μg/g body weight) and medetomidine hydrochloride (1 μg/g body weight), and perfused with saline to remove blood from tissues before their collection. Prior to perfusion, blood was extracted by retro-orbital collection and used for the determination of sobetirome, T4, and T3 plasma concentrations. Tissues (hemi-brain, liver, and heart) were harvested for further analyses.
Gene expression
RNA was isolated from individual hemi-brain, liver, and heart after treatment. Total RNA was extracted using TRIZOL reagent (Invitrogen; 15596026) following the manufacturer's recommendations with an additional chloroform extraction. RNA quality was assessed with the Agilent 2100 Bioanalyzer. cDNA was prepared from 250 ng of RNA using the high-capacity cDNA reverse transcription kit (Applied Biosystems). A cDNA aliquot corresponding to 5 ng of the starting RNA was used. Real-time PCR was performed with the TaqMan universal PCR master mix, No Amp Erase UNG (Applied Biosystems), on a 7900HT fast real-time PCR system (Applied Biosystems). The PCR program consisted of a hot start of 95°C for 10 minutes, followed by 40 cycles of 15 seconds at 95°C and one minute at 60°C. PCRs were performed in triplicate using the 18S gene as internal standard and the two-cycle threshold method for analysis.
The expression of the following T3-dependent genes was measured using Applied Biosystems TaqMan probes. In the brain: Abcd2 (ATP binding cassette subfamily D member 2), Cntn2 (contactin 2), Enpp2 (ectonucleotide pyrophosphatase/phosphodiesterase 2), Hr (hairless), Mc4r (melanocortin 4 receptor), Mme (membrane metallo-endopeptidase), Nefh (neurofilament heavy polypeptide), Nefm (neurofilament, medium polypeptide), Sema7a (semaphorin 7A), Flywch2 (Flywch family member 2); in the liver: Dio1 (type 1 iodothyronine deiodinase) and Ucp2 (uncoupling protein 2); and in the heart: Atp2a2 (sarcoplasmic reticulum Ca2+ ATPase pump) and Hcn2 (hyperpolarization-activated cyclic nucleotide-gated channel). Data were expressed relative to the values obtained in tissues from the Wt mice treated with vehicle (taken as 1.0) after correction for 18 S RNA.
Hormonal determinations in plasma
High specific activity 125I-T3 and 125I-T4 (3000 μCi/μg) were labeled with 125I (Perkin Elmer, NEZ033A) using as substrates (3 –5)-T2 (Sigma–Aldrich; D0629) and T3 (Sigma–Aldrich; T2877), respectively, according to Weeke and Orskov (49) and Obregon et al. (50). The separation of the labeled products was modified using ascending paper chromatography for 16 hours in the presence of butanol:ethanol:ammonia 0.5N (5:1:2) as solvent. The 125I-T3 and 125I-T4 were eluted and kept in ethanol at 4°C.
T3 and T4 were extracted from individual 80 μL aliquots of plasma with methanol (1:6), evaporated to dryness, and taken up in the radioimmunoassay (RIA) buffer for determinations. Determinations of T3 and T4 were performed, as previously described (51,52), with the dynamic range being 0.4–100 pg T3/tube and 2.5–320 pg T4/tube. For both RIAs, the coefficients for intra- and inter-assay variation were 5% and 10–15%, respectively. The cross-reactivity of both sobetirome and Sob-AM2 on the T3 and T4 antiserum was calculated to be <0.12% in every case.
Sobetirome determinations in plasma and brain
Sobetirome was measured in plasma and brain homogenate using liquid chromatography–tandem mass spectrometry (LC-MS/MS), as previously described (38), with the following modifications. The plasma sample (25 μL) was extracted with 125 μL of acetonitrile, and an extracted standard curve ranging from 1 to 2000 ng/mL was prepared in control plasma. The brain homogenate (125 μL, 200 mg of tissue/mL) was extracted with 500 μL of acetonitrile, and an extracted standard curve ranging from 1 to 100 ng/g of tissue was prepared in control brain homogenate. Control samples from Wt and Mct8/Dio2KO vehicle-treated mice had no detectable levels of sobetirome.
Statistical analysis
Differences between means were obtained by one-way analysis of variance and the Bonferroni's post hoc test to correct for multiple comparisons using the GraphPad software (GraphPad Software, Inc., La Jolla, CA). Levels of significance were set at p < 0.05, p < 0.01, and p < 0.001.
For sobetirome measurements, statistical significance was determined by Student's t-test comparing the amount of sobetirome in plasma and brain after treatment with sobetirome or the sobetirome prodrug (Sob-AM2). The level of significance was set at p ≤ 0.05.
Results
Sobetirome and Sob-AM2 distribution in plasma and brain
Sobetirome was measured in the plasma and brain samples using LC-MS/MS. Samples were analyzed four to six hours after drug treatment, which consisted of once-daily 1 mg/kg of sobetirome (i.p.) or 0.3 mg/kg of Sob-AM2 (i.p.) for seven days. The amount of sobetirome measured in plasma was 522 ± 65 ng/mL after administration of sobetirome (1 mg/kg), and 66 ± 13 ng/mL after the administration of Sob-AM2 (0.3 mg/kg). After dose correction of the sobetirome concentration data from the different doses of sobetirome and Sob-AM2 (Fig. 1), sobetirome treatment resulted in around 2.5-fold greater exposure of sobetirome in plasma compared to Sob-AM2 treatment. The amount of sobetirome in the brain was 20 ± 4 ng/g after sobetirome treatment and 10 ± 3 ng/g after Sob-AM2 treatment. After dose correction (Fig. 1), Sob-AM2 treatment resulted in 1.8-fold more sobetirome in the brain than treatment with the parent drug sobetirome.
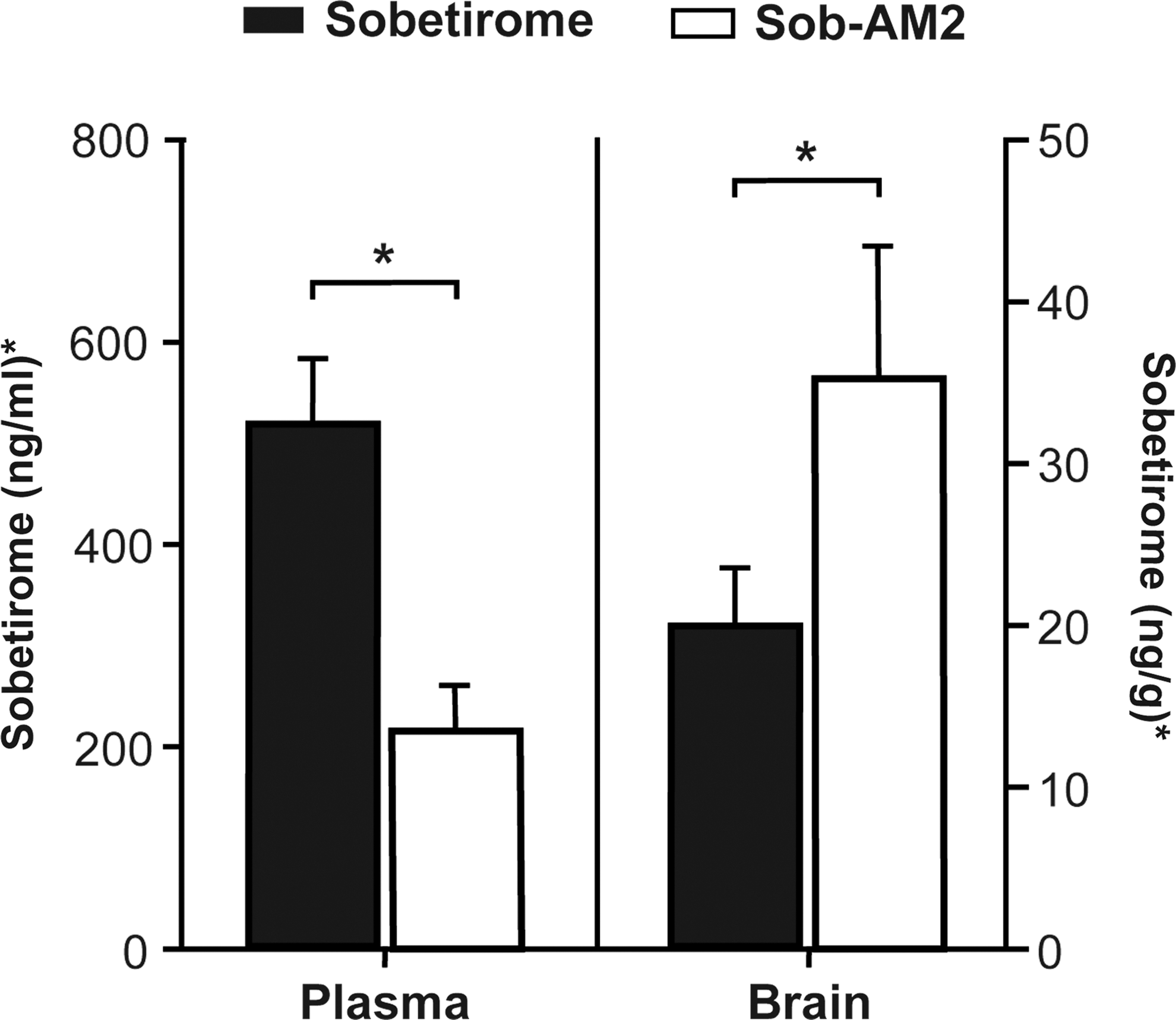
Sobetirome concentrations in plasma and brain. Mice lacking Mct8 and deiodinase type 2 (Mct8/Dio2KO) were treated for seven days with once-daily intraperitoneal (i.p.) injections of sobetirome (1 mg/kg) or Sob-AM2 (0.3 mg/kg). The plasma and brain were harvested four to six hours after the last dose and were analyzed for sobetirome concentration by liquid chromatography–tandem mass spectrometry. *The Sob-AM2 concentrations were dose-corrected in order to compare sobetirome (1 mg/kg) and Sob-AM2 (0.3 mg/kg).
Effects of sobetirome and Sob-AM2 in plasma TH levels
The effects of sobetirome and Sob-AM2 treatments on plasma TH levels were evaluated in Mct8/Dio2KO mice. Mct8/Dio2KO mice treated with vehicle presented the distinctive thyroid function profile of single Mct8KO mice (17,18), as previously reported (46). This involved decreased T4 and increased T3 plasma levels compared to Wt mice (Fig. 2) consistent with the peripheral hyperthyroidism characteristic of MCT8 deficiency. Sobetirome and Sob-AM2 treatment (once-daily, 7 days) of Mct8/Dio2KO animals had similar effects on plasma T4 and T3 levels. Both treatments decreased T4 levels in Mct8/Dio2KO mice to approximately 10% of the Wt and 30% of the Mct8/Dio2KO values of animals treated with vehicle (Fig. 2). Moreover, T3 levels decreased to approximately 15% of the Wt values and to 10% of the values of Mct8/Dio2KO mice treated with vehicle (Fig. 2). Thus, daily treatment with sobetirome or Sob-AM2 for seven days resulted in depletion of circulating THs.
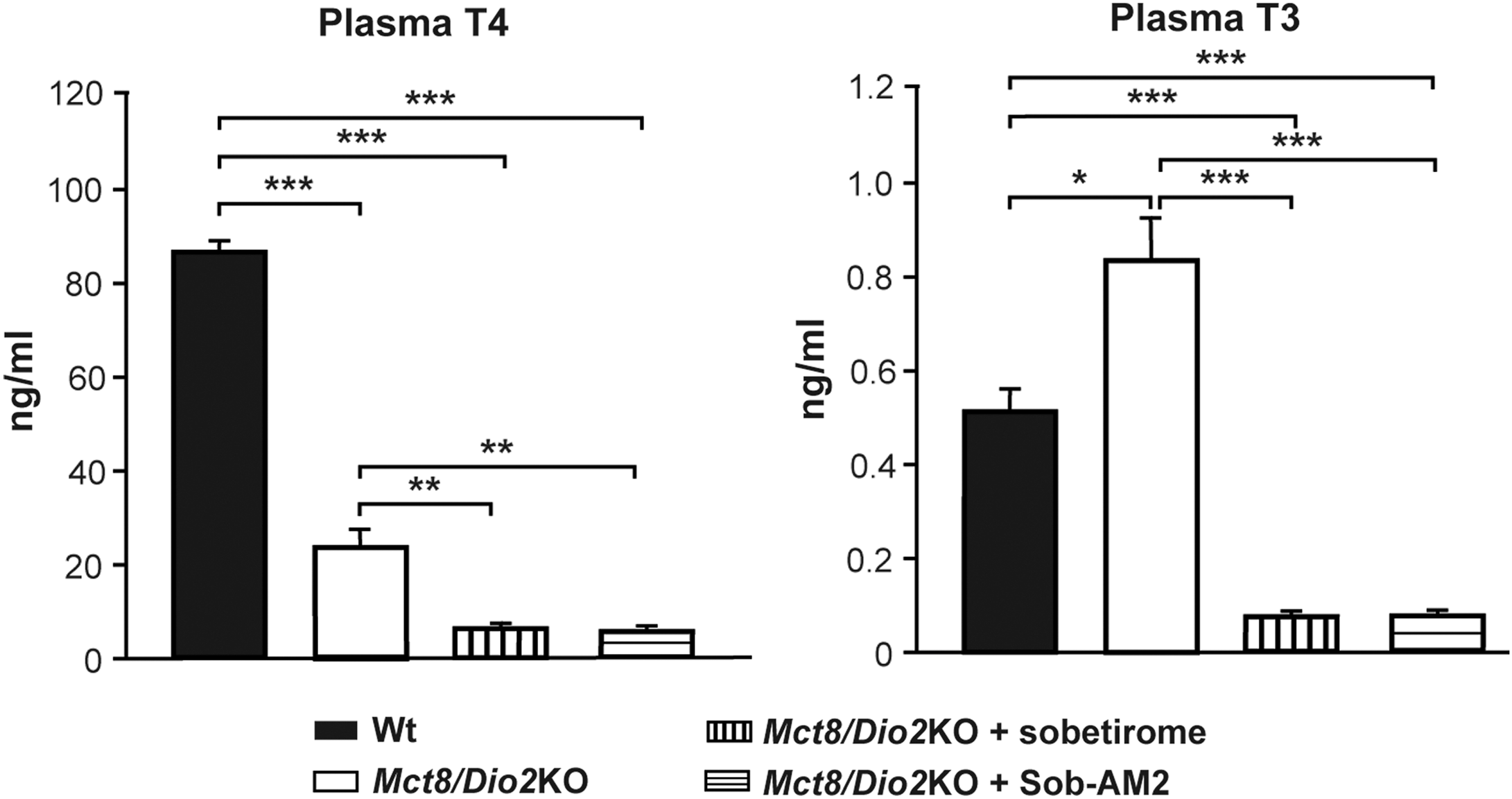
Thyroxine (T4) and triiodothyronine (T3) plasma levels in wild-type (Wt) and Mct8/Dio2KO mice in basal conditions and after sobetirome and Sob-AM2 treatments. Measures were obtained by specific radioimmunoassays. *p < 0.05, **p < 0.01, and ***p < 0.001 were determined by one-way analysis of variance (ANOVA) and Bonferroni's post hoc test.
Effects of sobetirome and Sob-AM2 in the liver and heart
In the liver, the expression of the T3-dependent gene encoding the enzyme type 1 iodothyronine deiodinase (Dio1) responsible for metabolizing T4 into T3 (53) was increased fivefold in the Mct8/Dio2KO mice compared to Wt animals (Fig. 3). This increase in the expression of Dio1 in Mct8/Dio2KO is similar to what has been observed in Mct8KO animals (17,18). The expression of another T3-dependent gene, Ucp2, which has been found to be increased in the liver of Mct8/Dio2KO mice (46), was also studied. Its expression was found to be altered in Mct8/Dio2KO mice (a fivefold increase for Ucp2). The expression levels of these genes did not return to the Wt values after treatment with sobetirome or Sob-AM2 (Fig. 3).

Dio1, Ucp2 expression in the liver, and Hcn2 and Atp2a2 expression in the heart in Wt and Mct8/Dio2KO mice in basal conditions and after sobetirome and Sob-AM2 treatments. Gene expression measurements were obtained by quantitative polymerase chain reaction, and the data are expressed relative to 18S RNA. *p < 0.05, **p < 0.01, and ***p < 0.001 were determined by one-way ANOVA and Bonferroni's post hoc test. Dio1, type 1 iodothyronine deiodinase mRNA; Ucp2, uncoupling protein 2 mRNA; Hcn2, hyperpolarization-activated cyclic nucleotide-gated channel mRNA; Atp2a2, sarcoplasmic reticulum Ca2+ ATPase pump mRNA.
In the heart, the expression of the T3-dependent genes Hcn2 and Atp2a2 was not affected in Mct8/Dio2KO mice compared to Wt animals. However, the treatment with sobetirome and Sob-AM2 increased the expression of Hcn2 but not that of Atp2a2 (Fig. 3).
Effects of sobetirome and Sob-AM2 on the CNS
The ability of systemically dosed sobetirome and Sob-AM2 to exert an action at the genomic level in the brain in Mct8 deficiency was next evaluated. Genes were selected that have been previously described to be decreased in the cerebral cortex of both hypothyroid and Mct8/Dio2KO mice (45): Cntn2, enriched in oligodendrocytes and neurons; Nefh and Nefm, enriched in neurons; and Sema7a, enriched in endothelial cells. Other T3-dependent genes were also evaluated that have recently been shown to be decreased in the cerebral cortex and the striatum of both hypothyroid and Mct8/Dio2KO mice by RNA-sequencing (Beatriz Morte, pers. commun., manuscript under preparation): Hr, which is enriched in oligodendrocytes, astrocytes, and neurons; Mme and Flywch2, which are enriched in neurons; Enpp2 and Mc4r, which are enriched in oligodendrocytes; and Abcd2, which is enriched in astrocytes. Furthermore, with the exception of Cntn2 and Nefh, all the above genes are directly regulated by T3 in primary mouse cerebrocortical cells (54).
Mct8/Dio2KO animals presented a threefold decrease in the expression of Hr and a twofold decrease in the expression of Abcd2, Mme, and Flywch2 compared to Wt (Fig. 4). Treatment with sobetirome and Sob-AM2 had similar effects in each of these genes, increasing the expression of Hr 3.5-fold to equal the expression level in Wt mice, and increasing the expression of Abcd2 and Mme 1.7- and 1.5-fold, respectively. Flywch2 expression increased threefold after sobetirome treatment and 2.2-fold after treatment with Sob-AM2 (Fig. 4). The expression of Cntn2, Nefh, Nefm, and Sema7a was decreased in the Mct8/Dio2KO mice, as previously reported (45): Cntn2 (1.7-fold), Nefh (1.5-fold), Nefm (1.8-fold), and Sema7a (1.8-fold). Neither sobetirome nor Sob-AM2 treatments had any effect in the expression of these four genes in Mct8/Dio2KO mice (Fig. 4). The deregulated expression of Enpp2 and Mc4r was not statistically significant in Mct8/Dio2KO animals, and neither sobetirome nor Sob-AM2 treatments had any effect on the expression of these genes (Fig. 4).
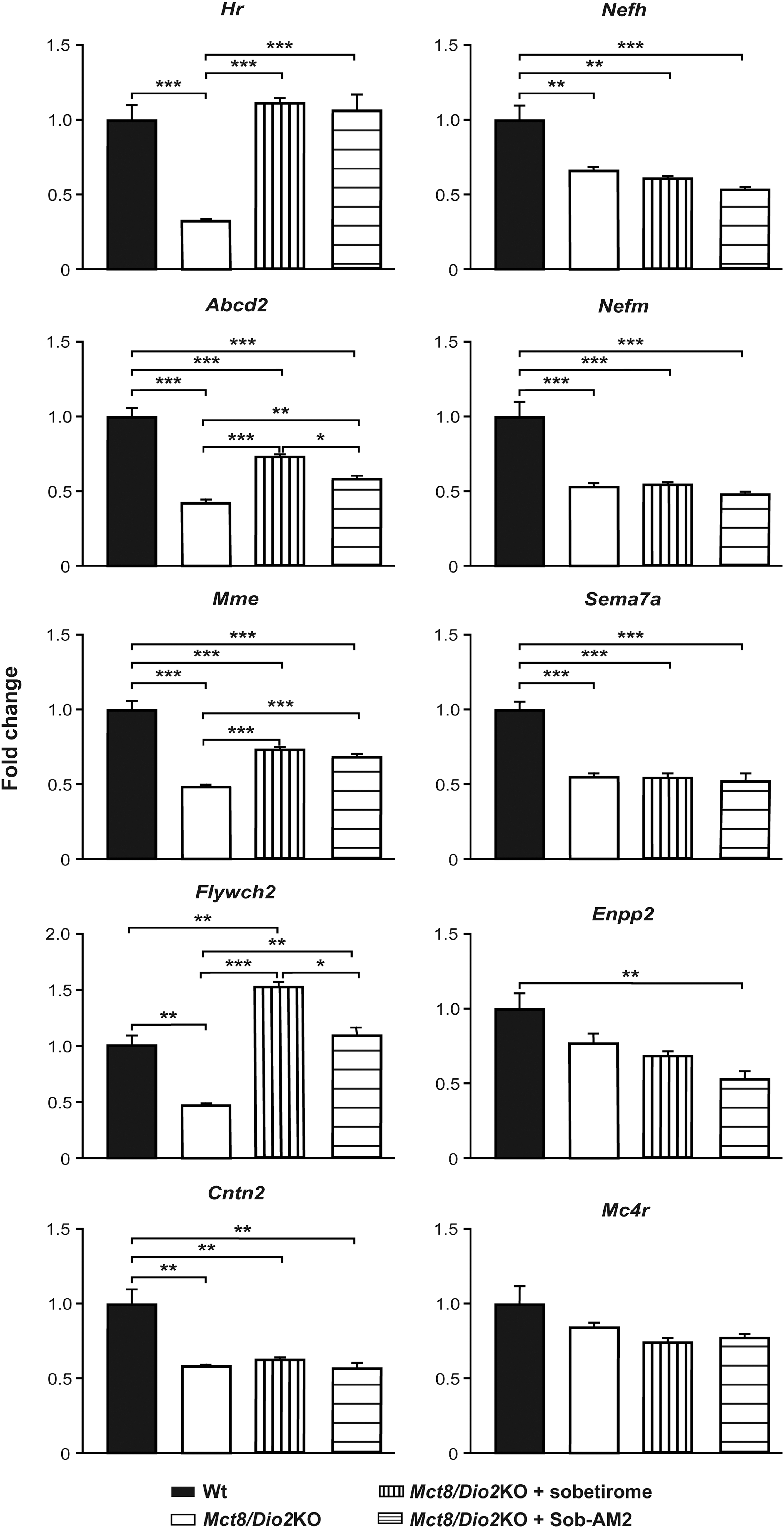
Gene expression analysis of T3-dependent genes in the brain in Wt and Mct8/Dio2KO mice in basal conditions and after sobetirome and Sob-AM2 treatments. Measurements were obtained by quantitative polymerase chain reaction, and the data are expressed relative to 18S RNA. *p < 0.05, **p < 0.01, and ***p < 0.001 were determined by one-way ANOVA and Bonferroni's post hoc test. Abcd2, ATP binding cassette subfamily D member 2 mRNA; Cntn2, contactin 2 mRNA; Enpp2, ectonucleotide pyrophosphatase/phosphodiesterase 2 mRNA; Flywch2, Flywch family member 2 mRNA; Hr, hairless mRNA; Mc4r, melanocortin 4 receptor mRNA; Mme, membrane metallo-endopeptidase mRNA; Nefh, neurofilament heavy polypeptide mRNA; Nefm, neurofilament, medium polypeptide mRNA; Sema7a, semaphorin 7A mRNA.
Discussion
MCT8 deficiency results in a hypothyroid state in the brain with contrasting peripheral hyperthyroidism. Thyromimetic agents that can access the brain and restore TH action while at the same time restoring euthyroid status to the periphery would potentially provide clinical benefit in MCT8 deficiency. This study explored the peripheral and central effects of the TH agonist sobetirome and its CNS-selective amide prodrug, Sob-AM2, in mice deficient in Mct8, with special focus on their ability to reach the brain and exert a thyromimetic action at the genomic level in the absence of Mct8. Because sobetirome and Sob-AM2 are known to distribute to the CNS from a systemic dose, it is important to know whether CNS distribution of these agents is affected by MCT8 deficiency. The ultimate goal was to obtain evidence that sobetirome and Sob-AM2 are valuable compounds for the treatment of patients with MCT8 mutations.
It was found that both sobetirome (1.0 mg/kg) and Sob-AM2 (0.3 mg/kg) reduce plasma T4 and T3 in Mct8/Dio2KO juvenile mice when dosed once-daily for seven days. This same dose (1.0 mg/kg) and dosing schedule (7-day/q.d.) of sobetirome in Wt mice was shown previously to suppress TSH (>50%); T4 and T3 levels were not measured in this study (33). A subsequent seven-day/q.d. study in rats using a lower dose (0.16 mg/kg) of sobetirome resulted in a similar level of TSH suppression and a significant 35% reduction in plasma T4; plasma T3 was also reduced but not significantly (55). This suggests that the observed T4 and T3 reductions in Mct8/Dio2KO mice upon sobetirome treatment occur via a mechanism involving sobetirome induced central suppression of TSH. Treatment with the CNS-selective Sob-AM2 results in a similar depletion of T4 and T3, suggesting that elevated brain and reduced blood levels of sobetirome are sufficient to produce this effect on the hypothalamic–pituitary–thyroid axis.
In the liver, the expression of Dio1 was increased in double Mct8/Dio2KO mice, as is the case of single Mct8KO (17,18). Ucp2 mRNA was also increased in Mct8/Dio2KO mice. In all cases, these alterations are probably due to high T3 plasma levels that impact liver gene expression. After treatment with either sobetirome or Sob-AM2, the expression of these genes did not return to the Wt values, despite the decreased T3 levels in the plasma of these animals. Therefore, the high expression of Dio1 and Ucp2 observed in the liver of both sobetirome- and Sob-AM2-treated Mct8/Dio2KO, in comparison to Wt values, could be due to direct sobetirome agonist effect in the liver, as previously observed with other thyromimetic agents (28). These results, together with evidence from a number of studies, indicate that sobetirome could be exerting a thyromimetic effect in the liver by virtue of the TRβ preference of liver and the sobetirome distribution preference of liver compared to other tissues (37).
Mct8 is expressed in the heart (4,56). However, in MCT8-deficient patients, the heart function is normal, despite exposure to high serum T3 levels. This indicates a partially impaired T3 transport to the heart in MCT8 deficiency and suggests the involvement of additional TH transporters (57). The present findings support this statement and point to a state of cardiac euthyroidism, as Atp2a2 and Hcn2 expression is normal in the heart of the vehicle-treated Mct8/Dio2KO mice. Interestingly, the expression of Hcn2 but not Atp2a2 in the heart was increased in Mct8/Dio2KO after treatment with sobetirome or Sob-AM2. The lack of response to sobetirome or Sob-AM2 treatments in Atp2a2 expression is consistent with the established cardiac-sparing properties of sobetirome (37); TH heart responses are mediated in large part by TRα1 (58), and sobetirome does not distribute efficiently to the heart compared to other tissues (33). However, the effect of these analogs in the heart will probably depend on the dosage, the amount reaching the tissue, and, as observed in this case, by the sensitivity of the different T3-dependent genes.
MCT8-deficient patients present with severe neurological deficits resulting from cerebral hypothyroidism, most likely due to impaired TH transport across the BBB (14 –16). A thyromimetic agent that could access the brain despite the MCT8 deficiency and restore at least a subset of cerebral T3 actions could provide clinical benefit to these patients. It was found that both sobetirome and Sob-AM2 reach the brain from a systemic dose in the absence of Mct8, indicating that these agents cross the BBB by an MCT8 independent mechanism. Whether that involves passive diffusion or active transport via a different transporter is unknown at present. The possibility that these compounds are transported across the BBB by Oatp1c1 cannot be ruled out. As human BBB presents a low content of OATP1C1, this could present a potential limitation for the clinical use of these analogs in MCT8 deficiency. However, since in humans there is a considerable expression of OATP1C1 in other barriers such as the choroid plexus (43), the treatment with these analogs could still be beneficial for patients. Sob-AM2 has greatly enhanced brain penetrance compared to unmodified sobetirome, as previously described, amounting to a nearly 10-fold increase in sobetirome total exposure and a 60-fold increase in brain:serum ratio, a metric for distribution to the CNS from systemic circulation (40). The full magnitude of this effect is not readily apparent from the data presented here (Fig. 1) because this study involved brain sampling at four to six hours post-thyromimetic administration, a time period considerably later than the time (T max) at which maximal brain concentrations are reached (C max) following a systemic Sob-AM2 dose, which is about 30 minutes. Nevertheless, a significant increase in sobetirome brain levels delivered by Sob-AM2 versus sobetirome was observed at this later brain sampling time point.
In addition to the bioanalytical measurements of sobetirome and Sob-AM2 brain penetrance, the functional thyromimetic effects on brain TH target genes were examined in Mct8/Dio2KO mice. Altogether, the results indicate that both sobetirome and Sob-AM2 are able to reach the brain and mediate the expression of some T3-dependent genes in the absence of Mct8. Brain expression of Hr, Abcd2, Mme, Flywch2, Enpp2, and Mc4r are decreased in hypothyroid and Mct8/Dio2KO mice (Beatriz Morte, pers. commun., manuscript under preparation) and are also directly regulated by T3 (54). The expressions of Hr, Abcd2, Mme, and Flywch2 were decreased in Mct8/Dio2KO mice, and treatment with both sobetirome and Sob-AM2 increased the expression of these genes, reaching, or even increasing, normalized Wt control values in the case of Hr and Flywch2. This indicates that sobetirome and Sob-AM2 are able to exert an action in the expression of some T3-dependent genes mainly expressed in neurons and astrocytes. The expression levels of the genes Enpp2 and Mc4r, which are enriched in oligodendrocytes, were not altered in Mct8/Dio2KO mice and were not increased after treatment with sobetirome or Sob-AM2. The expression of brain Cntn2, Nefh, Nefm, and Sema7a was also studied. These genes are altered in Mct8/Dio2KO mice at similar developmental stages as those used in the present study (45). The expression of all of these genes was decreased in Mct8/Dio2KO mice compared to Wt, and no effect of treatment with sobetirome or Sob-AM2 was observed. The reason underlying the lack of action of the treatment in inducing the expression of certain T3 target genes is not known and could depend on the duration and the timing of thyromimetic treatment, the age of the animals, the different target brain cells types, and/or the TH receptor subtype and TH receptor associated factors mediating the expression of each gene (34,59,60).
To conclude, the present studies indicate that in the absence of MCT8, sobetirome and Sob-AM2 are able to distribute to the brain from a systemic dose and exert thyromimetic actions. At the dose and duration in which brain distribution occurs, substantial circulating T4 and T3 depletion is also observed with both agents. Cerebral hypothyroidism, particularly during brain development, and peripheral hyperthyroidism are the two main TH abnormalities in MCT8 deficiency that produce the array of afflictions in patients with this inborn error. Sobetirome and Sob-AM2 at least partially address both the central and peripheral issues, suggesting that these thyromimetics have a profile that may be clinically beneficial to patients with MCT8 deficiency.
Footnotes
Acknowledgments
This work was supported by the Spanish Ministry of Economy Industry and Competitiveness (SAF2014-54919-R and SAF2017-86342-R to A.G.F.), the Sherman Foundation (OTR02211 to A.G.F.), and the National Institutes of Health (DK52798 to T.S.S. and 2T32DK007680-21 to M.D.H.), and the National Multiple Sclerosis Society (FG 2023A 1/2 to M.D.H.). The cost of this publication has been paid in part by FEDER funds. We thank Beatriz Morte for her advice in the selection of T3-dependent genes and Eva Ausó for helpful reading of the manuscript. We also thank María Camino de Lucas for animal care and the Bioanalytical Shared Resource/Pharmacokinetics Core Facility, which is part of the University Shared Resource Program at Oregon Health and Science University, for analytical support.
Author Disclosure Statement
T.S.S. is a founder of NeuroVia, Inc., and an inventor of a licensed pending patent application on prodrugs of sobetirome. T.S.S. and M.D.H. are inventors on a licensed pending patent application for the use of sobetirome in demyelinating diseases. The remaining authors have nothing to disclose.