
Abstract
Background:
There is growing evidence that sphingosine-1-phosphate (S1P), a pleiotropic bioactive sphingolipid metabolite synthesized intracellularly by two closely related sphingosine kinases (SphKs), SphK1 and SphK2, is involved in inflammation. However, the role of SphKs/S1P/S1P receptors (S1PRs) in autoimmune thyroiditis (AIT) has not been studied to date.
Methods:
This study examined whether SphK1/S1P/S1PR1 signaling is aberrantly altered in thyroid tissues and serum of both AIT patients and a spontaneously autoimmune thyroiditis (SAT) mouse model. Murine CD4+T cells were employed to further investigate the downstream signaling of SphK1/S1P/S1PR1. Furthermore, a total of 102 NOD.H-2h4 mice, randomly divided into different groups, were used to investigate the therapeutic effect of S1PR1 blockade and its potential mechanism.
Results:
We found that components of the SphK1/S1P/S1PR1 pathway were abnormally expressed in patients with Hashimoto thyroiditis and in a SAT mouse model. In addition, S1P could activate signal transducer and activator of transcription 3 (STAT3) through S1PR1 and its downstream signaling pathways in CD4+T cells of NOD.H-2h4 mice. Furthermore, an in vivo study demonstrated that blocking S1PR1 by FTY720 administration could reduce the incidence and severity of thyroiditis and goiter in SAT mice in a time-dependent manner. The proportions of STAT3-related and inflammation-related cell subtypes, such as T helper 1, T helper 17, and follicular T helper cells, were elevated in the SAT group when compared to the control group, and these cell subtypes decreased after FTY720 administration. Furthermore, the downstream inflammatory cytokines of STAT3 were also downregulated after FTY720 administration.
Conclusion:
The present study shows that blocking Sphk1/S1P/S1PR1 signaling can ameliorate the severity of AIT, providing evidence of a promising therapeutic target for AIT.
Introduction
Hashimoto's thyroiditis (HT), the most common form of autoimmune thyroiditis (AIT), affects 2–12% of the population and is more prevalent in women than in men (1,2). Environmental factors, such as increased levels of dietary iodine, are associated with the incidence and severity of HT; thus iodine is considered a modifying factor (3). NOD.H-2h4 mice express the K haplotype on the NOD genetic background and spontaneously develop autoimmune thyroiditis upon the addition of iodide in drinking water, provide a useful mouse model of iodine-induced HT (4,5). Although HT was first described over a century ago, currently, the therapy for primary and permanent hypothyroidism seen in many forms of HT consists of daily, lifelong, oral administration of synthetic levothyroxine. It is therefore a replacement therapy rather than treatment of the underlying pathogenesis of HT (6). Dietary supplementation with selenium is being evaluated as a tool to protect the thyroid gland from autoimmune damage, but results from randomized clinical trials are ambiguous (7). Therefore, HT remains a complex disease of unknown pathogenesis that awaits preventative interventions or novel forms of treatment.
There is growing evidence that sphingosine-1-phosphate (S1P), which is synthesized intracellularly by two closely related sphingosine kinases (SphKs), SphK1 and SphK2, is involved in inflammation and cancer. S1P acts as an intracellular messenger and as an extracellular ligand of five G-coupled cell surface receptors designated sphingosine-1-phosphate receptors 1–5 (S1PR1–S1PR5) and regulates numerous cellular processes important for cell growth survival, invasion, lymphocyte trafficking, and cytokine and chemokine production (8). Previous studies have indicated that abnormal levels of S1P are associated with the severity of many autoimmune diseases, such as multiple sclerosis, rheumatoid arthritis, and lupus erythematosus (9,10). Furthermore, modulation of S1PRs by immunomodulatory drug Fingolimod (FTY720) may ameliorate the severity of these diseases (10,11). However, the underlying molecular mechanism of how FTY720 works on lymphocytes remains unclear and it is still a major research focus.
As of now, little is known about SphKs/S1P/S1PR1 signaling and its role in AIT. Given the complex background, we investigated whether there is an abnormality in the expression of components of SphKs/S1P/S1PR1 signaling in HT patients and a spontaneously autoimmune thyroiditis (SAT) mouse model. We also investigated the relationship of S1P signaling CD4+T cells from SAT mice and whether blocking SphKs/S1P/S1PR1 signaling by FTY720 may have beneficial effects on the severity of autoimmune thyroiditis.
Methods and Materials
Patients and samples
Human thyroid tissue was obtained as described previously (12). In brief, subjects underwent surgery for thyroid adenomas or nodules or for a partial thyroidectomy to relieve tracheal compression caused by goiter in Hashimoto thyroiditis. Thyroid tissue for the normal control group (NOR group) as indicated in Figure 1, was obtained from subjects who were euthyroid and not exposed to radioactive iodine, antithyroid drugs, or thyroid hormone replacement and who did not have systemic autoimmune disease or any severe illnesses. Subjects with malignant nodules were excluded. Serum samples were also obtained before surgery to test thyroid function and detect autoantibodies, which were assessed by an electrochemiluminescence method (Architect i2000SR, Abbott Laboratories). All of the patients who showed positive thyroid autoantibodies (TPOAb and TgAb >50 IU/mL) received a pathologic diagnosis of Hashimoto's disease. Age- and sex-matched controls negative for the presence of autoantibodies were included. It has been shown that thyroid disorders are more prevalent in women (1,13), and in order to be aligned with the clinical scenario, human thyroid tissue was obtained from female patients. The clinical characteristics are summarized in Supplementary Table S1.
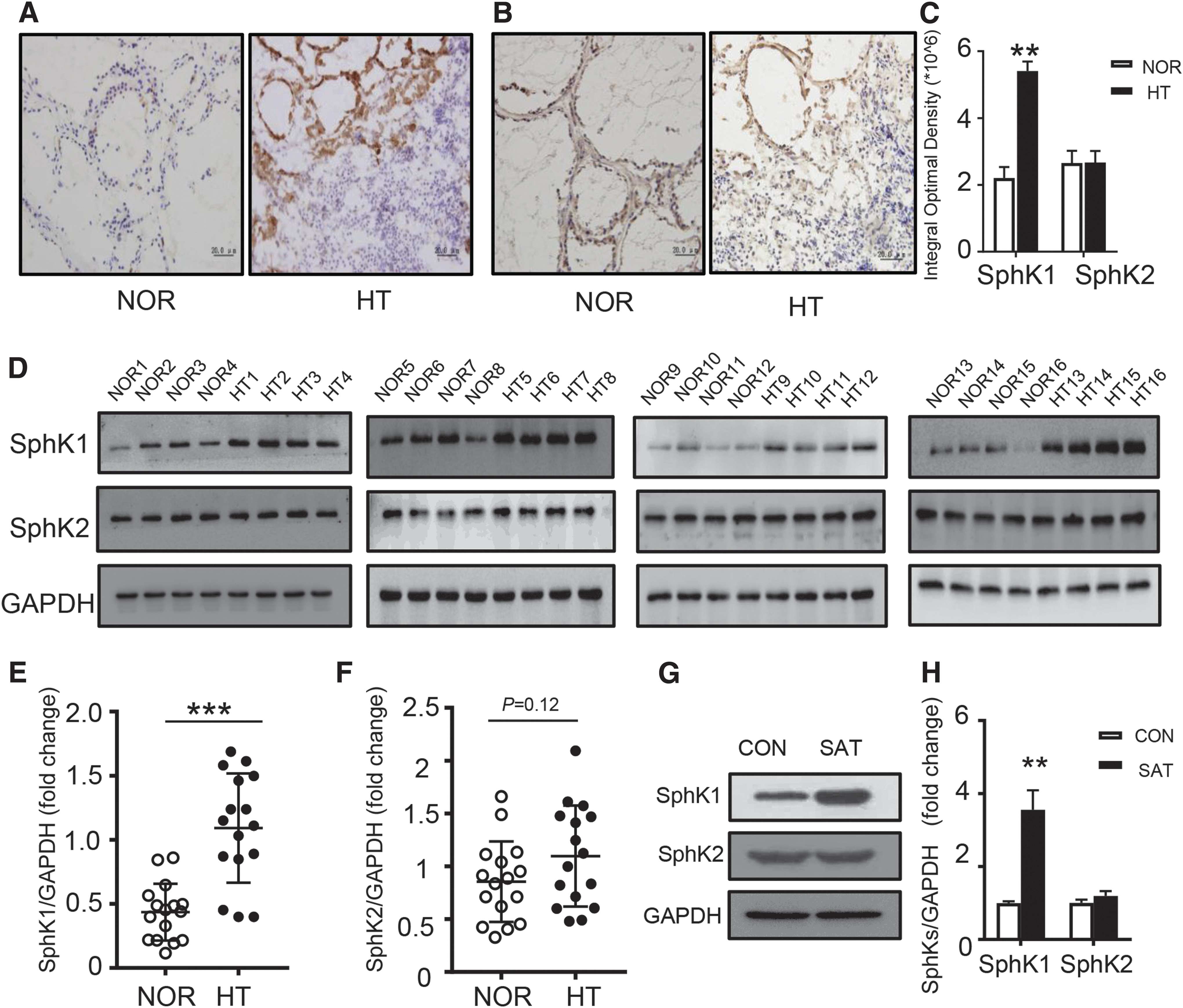
Expression of SphK1 and SphK2 in thyroid tissue.
As indicated in Figure 2, to investigate circulating S1P concentrations in HT patients, serum samples were derived from patients newly diagnosed with HT who were matched for age, sex, and body mass index. Thirty-nine patients with overt hypothyroidism (OH) and 34 patients with subclinical hypothyroidism (SCH) were enrolled. Likewise, 39 similarly matched euthyroid subjects with isolated autoantibody positivity (TPOAb and TgAb >50 IU/mL) and 41 NOR subjects were included (characteristics and diagnostic criteria are listed in Supplementary Table S2). Subjects with any identified acute or chronic illness, such as diabetes or systemic lupus erythematosus were excluded.

Concentrations of S1P and expressions of S1PRs in AIT.
All of the research protocols were approved by the Medical Ethics Committee of The First Hospital Affiliated to China Medical University and were congruent with the Declaration of Helsinki. All of the participants provided written informed consent after the research protocols were carefully explained to them.
Animals
NOD.H-2h4 mice were purchased from Jackson Laboratory). Animals were bred and maintained under specific pathogen-free conditions in a controlled temperature and humidity environment with 12-h day–night cycles in the animal facility of China Medical University as previously described (4). This study was carried out in strict accordance with the recommendations in the Guide for the Care and Use of Laboratory Animals of the National Institutes of Health. The protocol was approved by the Committee on the Ethics of Animal Experiments of China Medical University. All surgery was performed under anesthesia, and all efforts were made to minimize suffering.
In vivo experimental design
To induce autoimmune thyroiditis, mice were given 0.05% (500 mg/L) sodium iodide (NaI) in their drinking water during the study period. Normal control mice received regular drinking water throughout the experiment. The S1PR functional antagonist FTY720 was obtained from Sigma. FTY720 was dissolved in sterile phosphate-buffered saline (PBS) at a concentration of 0.1 mg/mL and was administered by daily oral gavage at a dose of 0.25 mL (1 mg/kg) once a day during the study period.
Experimental groups
Experiment one
To explore the role of SphKs/S1P/S1PRs signaling in the pathogenesis of autoimmune thyroiditis in the mouse model, we randomly selected 24 female NOD.H-2h4 mice and divided them into two groups: 12 mice with regular drinking water (CON group), and 12 mice with 0.05% NaI drinking water (SAT group).
Experiment two
To explore the preventive and therapeutic effect of the S1PR1 functional antagonist in SAT mice, we randomly selected 102 female NOD.H-2h4 mice and divided them into the following five groups: (i) 21 mice receiving regular drinking water (CON); (ii) 20 mice receiving 0.05% NaI drinking water (SAT); (iii) 20 NOD.H-2h4 for the prevention study (PRE), receiving FTY720 in PBS once a day from the beginning until the 16th week; (iv) 21 NOD.H-2h4 mice for the treatment study (TRE)—after 8 weeks of iodine supplementation, these were given FTY720 daily from the ninth week for another 8 weeks; (v) 20 mice to explore the effect of FTY720 (FTY), treated daily with FTY720 for 16 weeks. Blood (approximately 50 μL) was drawn from all groups every other week to monitor the titer of thyroglobulin antibodies (TgAb). All of the animals were anesthetized and sacrificed at the 16th week of the experiment. Thyroid tissues were removed and then washed with cold normal saline. Then, they were dried on a pad of filter paper and weighed on an electronic balance (BS210S, Sartorius)
Immunohistochemistry analysis of thyroid tissue
Immunohistochemistry was performed as previously described (14 –16). Thyroids were snap frozen in liquid nitrogen, cut into 5-μm sections, air dried, and fixed with cold paraformaldehyde for 10 min. The sections were immunostained using a rabbit polyclonal antibody against SphK1 and SphK2 (1:100, ab71700/Reacts with: Mouse, Rat, Human; 1:100 ab37977/Reacts with: Mouse and Human; Abcam). In brief, after rinsing three times using PBS-Tween, the slides were blocked in 5% normal goat serum for 30 min. Samples were incubated with primary antibodies at 4°C overnight and then incubated with the secondary antibody for 10 min. After washing out the unbound secondary antibody, the sections were subjected to a streptavidin peroxidase reaction using an SP Kit (MAIXIN-BIO). The expression of proteins was determined as previously described (17), and image information object definition (IOD) values were analyzed by Image-Pro Plus 5.0 (Media Cybernetics).
Measurement of S1P by liquid chromatography–tandem mass spectrometry
S1P concentration was measured by a method described previously (18). Tissues were homogenized in 500 μL of 1 M NaCl to keep the ratio of tissue weight to distilled water volume at 10% (w/v). A total of 50 μL of tissue homogenate was used for lipid extraction. Next, 50 μL of plasma were adjusted to a sample volume of 0.5 mL with 1 M NaCl and 0.5 mL MeOH, which then was used for lipid extraction. The stock solutions of S1P were serially diluted with methanol to obtain working solutions. All of the solutions were stored at −20°C and were brought to room temperature before use. Standard samples for S1P were from 0.15–0.8 μg/mL. Volumes of 50 μL homogenate, testing samples or standard samples, 100 μL 6 M HCl, and 0.75 mL CHCl3 were added into a 1.5-mL Eppendorf tube. The mixture was vortexed for 2 min followed by centrifugation at 11,000 g for 3 min, and then the lower organic phase was transferred with glass Pasteur pipettes to a clean glass centrifuge tube. The extraction steps were repeated, and the two CHCl3 phases were combined, evaporated at 50°C for 45 min in a vacuum concentrator, reconstituted in 100 μL MeOH/CHCl3 (4:1, v/v), centrifuged at 11,000 g for 3 min, and then 10 μL of the supernatant was injected into the liquid chromatography–tandem mass spectrometry system for analysis.
Isolation of murine CD4+ T cells and cell treatment
Single-cell suspensions in PBS were prepared from the spleen, filtered through a sterile mesh (BD Falcon), and washed. After erythrocytes were lysed in hemolysis buffer (144 mM NH4Cl and 17 mM Tris-HCl [pH 7.2]), the remaining cells were resuspended in magnetic-activated cell sorting (MACS) buffer (PBS containing EDTA and 3% calf serum) and incubated with anti-CD4 MicroBeads for 20 min on ice. Mice CD4+ T cells were isolated by using MACS according to the manufacturer's instruction (Miltenyi Biotec). The purities of mice CD4+ cells routinely reached >95%.
For experiments shown in Figures 3 –5 and Supplementary Figure S1B–C, CD4+T cells from 6- to 8-week-old normal mice were rested in 10% fetal bovine serum–enriched RPMI 1640 medium and were prestimulated by incubation for 24 h in 6-well plates in 2 μg/mL each of adherent anti-CD3 and anti-CD28 mAbs (eBioscience) before adding experimental reagents. S1P (Sigma) was dissolved in 100% methanol, aliquoted, and stored at −20°C until use. S1P administration for 1 h at a final concentration of 0–1 μM, which had already been established as a suitable dose for S1P-induced signaling (19), was used in all experiments. Pre-treatment with FTY720 (Sigma) for 24 h at 10–100 nM was performed in all of the experiments. W146 and SE2871 were obtained from Santa Cruz Biotechnology and were used at 1–5 mM over 1 h. These concentrations were similar as in previous studies and reflect appropriate concentrations for studying signaling (19,20). The Janus Kinase 2 (JAK) inhibitor AZD1480 was from Santa Cruz Biotechnology and used at the indicated concentrations over 1 h (Fig. 4 and Supplementary Fig. S1).
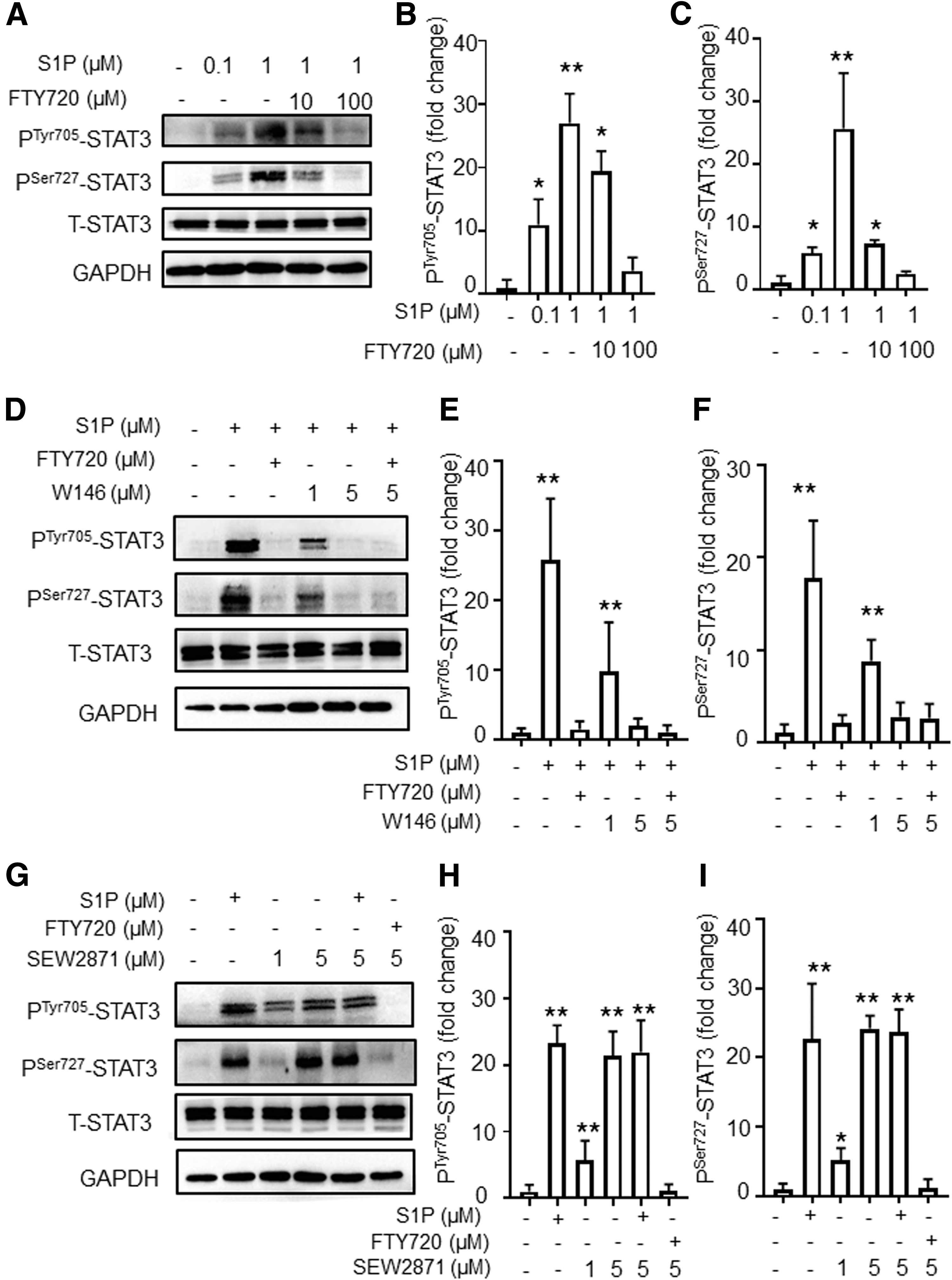
S1P activates STAT3 via S1PR1 in CD4+T cells.
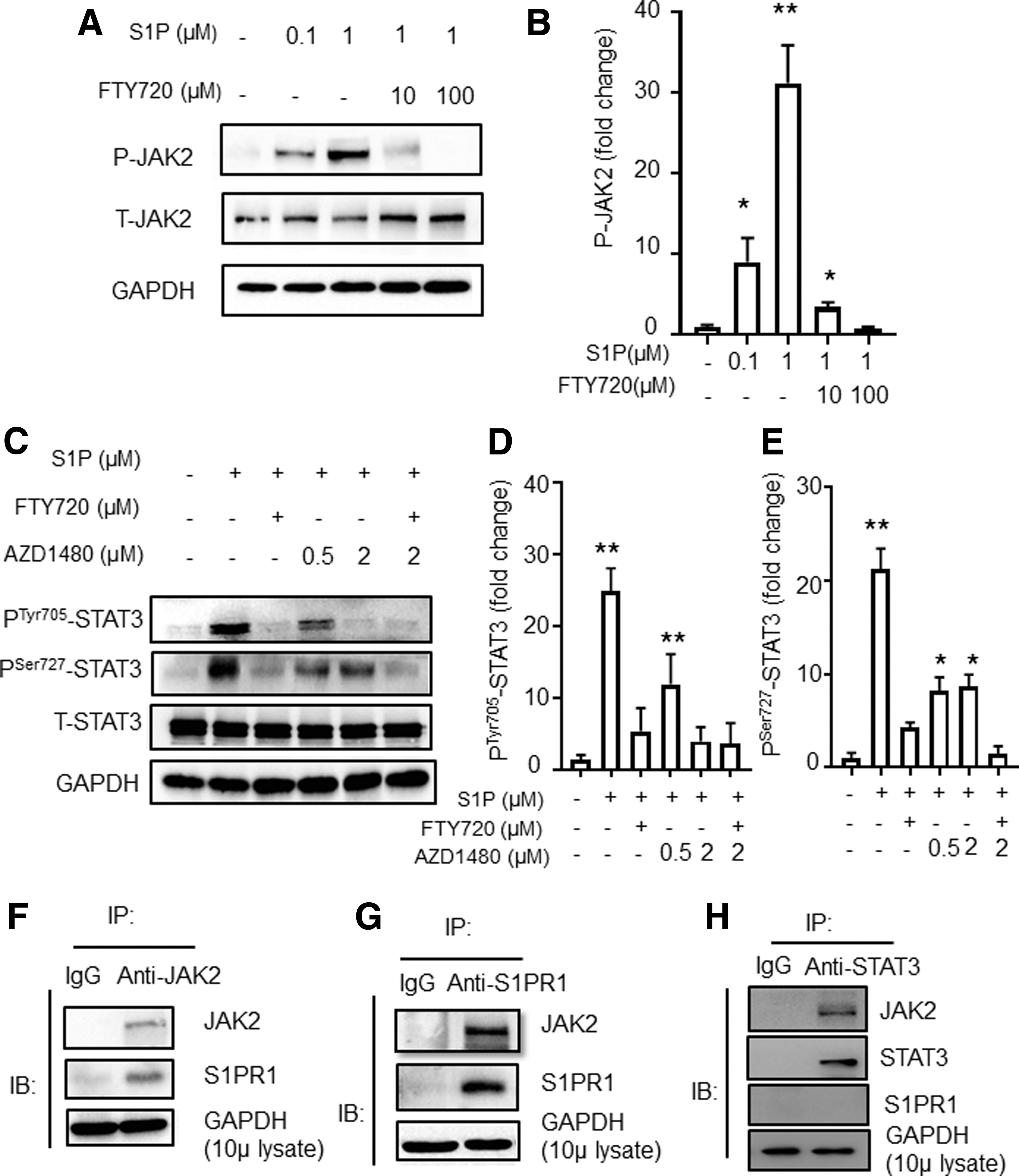
S1P increases PSer705 STAT3 via the S1PR1-JAK2 pathway.
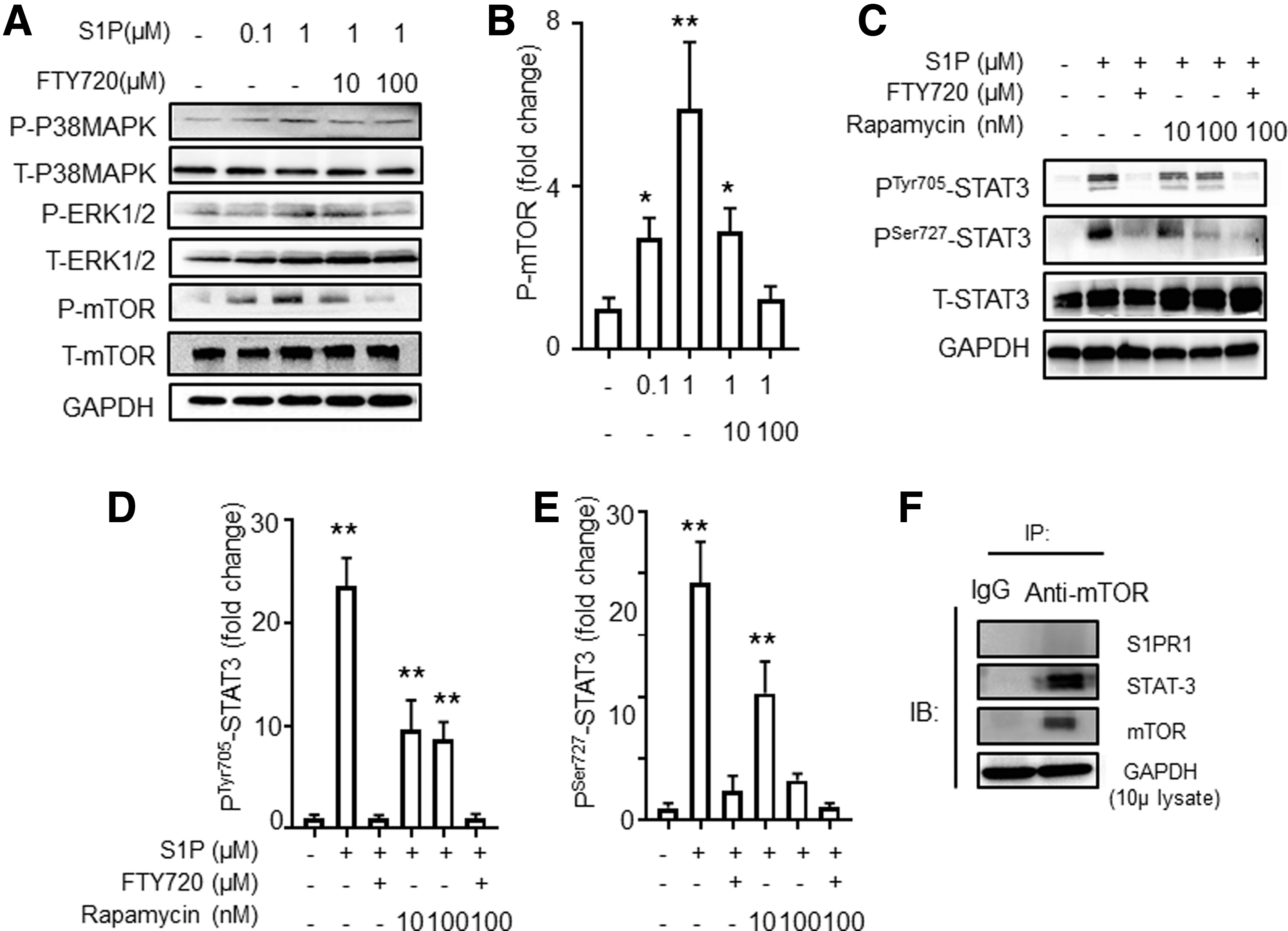
S1P increases PTyr727STAT3 via the S1PR1-mTOR pathway.
Evaluation of thyroiditis and measurement of serum TgAb
Thyroid tissues were fixed in 10% formalin and embedded in paraffin. Tissue sections (5 μm) were prepared and stained with hematoxylin and eosin (H&E). Briefly, H&E-stained thyroid sections were graded on the following scale according to the area of lymphocytic infiltration (21): 0, normal; 1+, at least 125 lymphocyte infiltrates in one or several foci; 2+, up to 25% of the gland; 3+, 25–50% of the gland; and 4+, more than 50% of the gland. The final thyroiditis scores were expressed as means of at least three noncontiguous sections from each thyroid gland.
The preparation for mouse Tg was the same as described previously (21). Murine sera were analyzed in 96-well plates, which were coated overnight with 1 mg/mL of purified mTg in carbonate-bicarbonate buffer (pH 9.6). Plates were washed and blocked with 1% bovine serum albumin for 10 min. Peroxidase-labeled goat anti-mouse immunoglobulin G (0.1 mL, Sigma) was diluted 1:250 with PBS, added to each well, and incubated at 37°C for 30 min. After washing, 0.1 mL tetramethylbenzidine was added to each well and plates were incubated at room temperature for 10 min. The positive control well turned blue and 0.1 mL 2 N hydrochloric acid was added to stop the reaction. Absorbance at 450 nm (TECAN, F200PRO) was measured after termination.
Flow cytometry
Spleens were harvested from freshly sacrificed mice at the indicated time points and pressed through a 200-gauge stainless steel mesh. Splenic mononuclear cells were isolated by Ficoll-isopaque density gradient centrifugation (Gibco BRL, Life Technologies Ltd.) as described previously (22). Cells were washed and immunostained with fluorochrome-conjugated monoclonal antibodies against CXCR5-APC, ICOS-PE-CyTM (eBioscience), CD4-FITC, IL-17A-Alexa Fluor®647, and IFN-γ-APC (BD Pharmingen). All of the extracellular and cell surface staining was performed according to manufacturers' protocols and methods we used previously (23 –25). The stained cells were then analyzed using a multicolor detection FACS Caliber flow cytometer (BD Biosciences) and FlowJo 7.6.1 software (Tree Star Inc.).
Western blot and CO-immunoprecipitation analysis
Whole protein extraction from cells and tissues was performed using a Total Protein Extraction Kit (KeyGen Biotech KGP 210). The supernatant was collected and the protein concentration measured using a BCA Protein Assay Reagent Kit. Tissues and cell lysates were diluted to contain the same concentration of proteins and then boiled for 10 min. Next, Western blotting and analysis were done as previously described (26,27). Proteins (50 μg) in 10-μL aliquots from the respective experimental groups were loaded onto 10% SDS-acrylamide gels then separated by application of a constant voltage. After blocking the nonspecific sites with PBS containing 0.05% Tween-20 and bovine serum albumin, the membranes were incubated overnight at 4°C with the indicated primary antibodies, including STAT3, P-STAT3, mammalian target of rapamycin (mTOR), P-mTOR, extracellular-signal-regulated kinase (ERK), P-ERK, P38 mitogen-activated protein kinase (P38MAPK), P-P38MAPK, (Cell Signaling Technology, Inc.). The blots were then incubated for 2 h with a horseradish peroxidase–conjugated secondary antibody.
For the interaction study of S1PR1 and protein kinases, 300 μg of protein lysates were precleared with protein A/G agarose (Santa Cruz) for 30 min at 4°C. After brief centrifugation, the supernatant was incubated with specific antibodies overnight. Protein agarose was added for an additional 1 h to capture immune complexes. Immunoprecipitates were collected by centrifugation, washed three times with modified RIPA buffer, and subjected to SDS-PAGE and immunoblotting analysis.
Reverse transcription–quantitative polymerase chain reaction analysis
Total RNA of CD4+T cells was extracted with TRIzol (Life Tech) according to a standard procedure, and a Nano Drop 2000C (Thermo Scientific) was used to determine RNA concentration and purity. Total RNA (1 μg) was reverse transcribed with random primers using a Reverse Transcription Reagent Kit (Takara Biotechnology Co., Ltd.) following the manufacturer's protocol. All of the amplification reactions were performed in triplicate on an ABI 9700 PCR meter (Applied Biosystems; Thermo Fisher Scientific, Inc). For quantitative real-time PCR, 25 ng of cDNA was used, and single transcript levels of genes were detected using a QuantiTect SYBR green master mix (TaKaRa). Experiments were performed in triplicate and data were quantified and analyzed using LightCycler 480 (Roche Diagnostics) analysis software. Primers used to detect synaptic transcripts are shown in Supplementary Table S3.
Statistical analysis
Data are expressed as means ± standard error or means ± standard deviation for normally distributed variables, medians with interquartile range for non-normally distributed variables, and as frequencies for categorical variables. For comparisons between study groups, normally distributed variables were assessed using one-way ANOVA followed by Bonferroni correction in paired comparisons. Variables that failed the normality test were assessed using Kruskal–Wallis one-way ANOVA for ranks in groups, and pairwise comparisons were performed using the Mann-Whitney rank sum test. Pearson's chi-squared test was used to compare the sex ratio. All of the calculated p values were two-sided, and p values <0.05 were considered to be statistically significant. All of the statistical analyses were performed with SPSS software (version 20.0).
Results
Expression of SphK1/S1P/S1PR1 signaling in HT patients and the SAT mouse model
Immunohistochemistry indicated that both SphK1 and SphK2 exhibit a similar cellular distribution, as both were mainly expressed within the cytoplasm of thyroid epithelial cells (Fig. 1A, B). The mean IOD of SphK1 was significantly higher in HT than in NOR thyroid samples (p < 0.01) (Fig. 1C); however, the expression of SphK2 showed no difference between the HT and NOR groups (Fig. 1C). Thyroid tissues of 16 HT patients and 16 normal controls were also assessed for SphK1 and SphK2 expression by Western blot (Fig. 1D). SphK1 had a higher expression in the HT group as compared to the NOR group (p < 0.001) (Fig. 1D, E). SphK2 showed no significant difference between the two groups (Fig. 1D, F). Consistent with the human thyroid tissues, Western blotting results of mice thyroid tissues indicated that SphK1 was highly expressed in the SAT group compared to the CON group (Fig. 1G, H). The expression of SphK2 showed no significant difference between the two groups (Fig. 1G, H).
Since SphK1 is upstream of S1P synthesis, we were interested in whether S1P would show a similar trend in AIT. Therefore, we measured S1P concentrations in thyroid tissues and serum of patients and mice by liquid chromatography–tandem mass spectrometry. S1P concentrations were higher in the thyroid tissues of HT patients compared to normal thyroid tissues (675.43 ± 209.1 ng/mg vs. 509.6 ± 184.3 ng/mg, p < 0.05) (Fig. 2A). S1P concentrations were also higher in the thyroid tissues of SAT mice compared with the CON group (251.7 ± 31.0 ng/mg. 143.7 ± 12.6 ng/mg, p < 0.05) (Fig. 2B). After demonstrating that S1P concentrations were higher in the thyroids of both patients and the SAT mouse model, we wondered whether circulating S1P changes exist in AIT. In general, HT progresses through three clinical stages that are defined by thyroid function: early stage (euthyroid), second stage (subclinical hypothyroidism), and final stage (overt hypothyroidism). It has previously been found that these clinically defined disease stages are closely correlated to the extent of autoimmune activity. Hence, we determined the S1P concentration in serum of different stages of HT: isolated Ab group, SCH group, and OH group (Supplementary Table S2). We found that S1P concentrations were higher in the OH, SCH, and isolated Ab groups than in the NOR group (Fig. 2C). In the mouse model, S1P levels were also higher in the serum of the SAT group than in the CON group (p < 0.05) (Fig. 2D). These results indicate that thyroid autoimmunity plays an important role in regulating circulating S1P concentrations.
Our previous study demonstrated that CD4+T cell play an important role in the pathogenesis of AIT (28). Since S1P is higher in AIT, we studied whether it affects CD4+T cells. Hence, we measured the expression of S1P receptors (S1PR1–S1PR5) in CD4+T cells from SAT mice and CON mice. Notably, the expression of S1PR1 was higher in SAT mice than in CON mice (p < 0.01). The expressions of S1PR2-5 showed no difference between the groups (Fig. 2E).
Modulation of signal transducer and activator of transcription 3 activity by S1P
Signal transducer and activator of transcription 3 (STAT3), an important multifunctional transcriptional factor involved in CD4+T cell differentiation, is known to be activated in AIT (29). STAT3 transcriptional activity is regulated by phosphorylation on Tyr-705 (PTyr705STAT3) and Ser-727 (PSer727STAT3). Based on our findings that S1P is increased in AIT, we hypothesized that STAT3 may be increased by S1P. To examine whether elevated S1P could induce STAT3 activity, a series of assays was conducted. As shown in Figure 3A–C, the activation of PSer727STAT3 and PTyr705STAT3 was induced by S1P and could be inhibited by FTY720 in murine CD4+T cells. Since multiple-protein coupled receptors (S1PR1-5) can bind to sphingosine-1-phosphate or FTY720, and S1PR1 expression was elevated in the SAT model as indicated in Figure 2E, this prompted us to investigate the role of S1PR1 in the activation of STAT3. We therefore employed the S1PR1-specific antagonist W146. As shown in Figure 3D–F, W146 inhibited the activation of STAT3 for both PTyr705STAT3 and PSer727STAT3. Adding FTY720 to W146 treatment showed no additive inhibitory effect of STAT3 in CD4+T cells (Fig. 3D–F). Next, we performed a stimulation test wherein the S1PR1-specific agonist SEW2871 was added, and elevated levels of PTyr705STAT3 and PSer727STAT3 were observed. Administration of S1P at 1 μM to SEW2871 treatment had no additive activating effect on STAT3 (Fig. 3G–I). Both inhibition and activation experiments demonstrated that S1PR1 may play an important role in S1P-induced STAT3 activation among S1PRs.
Next, we examined the responsiveness of CD4+T cells in physiological and pathophysiological conditions. As indicated in Supplementary Figure S1A, we found that the activation of STAT3 was more robust in CD4+T cells from the SAT group than cells from the control group. This is consistent with our previous findings shown in Figures 2E and 3, that S1PR1 plays a major role in S1P-induced STAT3 activation and that there is a higher expression of S1PR1 in SAT mice.
S1P-induced PTyr705STAT3 activation through JAK2
As JAK2 is known to be upstream of STAT3 in the IL-6-JAK2-STAT3 signaling pathway (30), we wondered whether JAK2 is also involved in S1P-S1PR1-induced STAT3 activation. We found that the level of P-JAK2 was elevated after S1P stimulation and that it was downregulated when combined with FTY720 treatment of CD4+T cells (Fig. 4A, B). Furthermore, upon addition of a JAK inhibitor, AZD1480, to S1P-treated murine CD4+T cells, both PTyr705STAT3 and PSer727STAT3 were downregulated (Supplementary Fig. S1B). However, it was surprising that the level of PTyr727STAT3 only showed a moderate reduction even after increasing the dose of AZD1480 (Fig. 4C–E and Supplementary Fig. S1B). Next, we performed immunoprecipitation in CD4+T cells with an anti-JAK2 specific antibody, followed by Western blot analysis. We detected a S1PR1-JAK2 interaction, which was confirmed by the converse immunoprecipitation with an anti-S1PR1 specific antibody (Fig. 4F, G). We also performed immunoprecipitation with an anti-STAT3 specific antibody, followed by Western blotting (Fig. 4H). We observed a STAT3-JAK2 interaction, which is consistent with a previous study (19).
S1P-induced PSer72 STAT3 activation through mTOR
Since the level of PSer727STAT3 was only partially reduced after the inhibition of JAK2, it is likely that other inflammatory-related pathways are involved in this process. Therefore, we performed Western blot analyses of phosphorylated proteins to assess the activation of S1PR1-downstream inflammatory pathways, including mTOR, ERK, and p38MAPK. As shown in Figure 5A and B, we found that the phosphorylation of mTOR, but not ERK or p38MAPK, was significantly increased after S1P stimulation and was decreased significantly after blocking with FTY720.
Next, we investigated whether mTOR was upstream of STAT3 in the S1P-S1PR1-STAT3 pathway. We analyzed P-STAT3 in murine CD4+T cells by using S1P to stimulate murine CD4+T cells and using rapamycin to inhibit mTOR. We confirmed that the phosphorylation of STAT3 was decreased at both Ser-727 and Tyr-705 after mTOR inhibition (Fig. 5C–E). However, the inhibitory effect of rapamycin on PTyr705STAT3 seemed milder than its effect on PSer727STAT3 even after increasing the dose (Supplementary Fig. S1C). The combination of FTY720 and rapamycin had no additive effect on the suppression of PSer727STAT3. Furthermore, we performed co-immunoprecipitation and demonstrated that mTOR interacted with STAT3 but not S1PR1 (Fig. 5F). These data suggest that S1P-induced PSer727STAT3 activation involves the mTOR pathway.
A novel target for the prevention and treatment of autoimmune thyroiditis: S1PR1
Since STAT3 plays an important role in the differentiation of CD4+T cells, and given that it can be inhibited by the S1P-S1PR1 pathway, we explored whether S1PR1 could be a good therapeutic target for AIT. As shown in Figure 6A, mice were divided into five groups. The results indicate that TgAb was higher in the SAT group when compared with the CON group (Fig. 6B). In addition, TgAb was still higher in the TRE group when compared with the CON group even after FTY720 treatment from week 8 to 12. However, there was no increase in the 14th week, and TgAb became lower in the TRE group when compared with the SAT group at week 16. The concentration of serum TgAb was lower in the PRE group when compared to the SAT group (Fig. 6B). Figure 6C indicates the thyroiditis scoring criteria with a scoring system from zero to four. As shown in Fig. 6D, the incidence of AIT was significantly higher in the SAT group compared with the CON group. Furthermore, the incidence of thyroiditis was lower in the PRE group when compared to the SAT group. However, there were no significant differences between the TRE group and the SAT group. As for the severity of the disease, the average thyroiditis score in the PRE group was significantly lower than in the SAT group (0.50 ± 0.13 vs. 3.40 ± 1.33, p < 0.001) and lower than in the TRE group (0.50 ± 0.13 vs. 1.81 ± 0.25, p < 0.001). Nevertheless, the average score in the TRE group was significantly lower than in the SAT group (1.81 ± 0.25 vs. 3.40 ± 1.33, p < 0.001). Three out of 21 mice developed autoimmune thyroiditis spontaneously after 16 weeks on a normal water supply in the CON group. In contrast, the incidence of goiter was higher in the SAT group when compared with the CON group. The incidence of goiter was lower in the PRE group when compared with the SAT group (Fig. 6E). These data suggest that blocking S1PR1 could ameliorate the severity of thyroiditis, but the therapeutic effect was also time dependent.
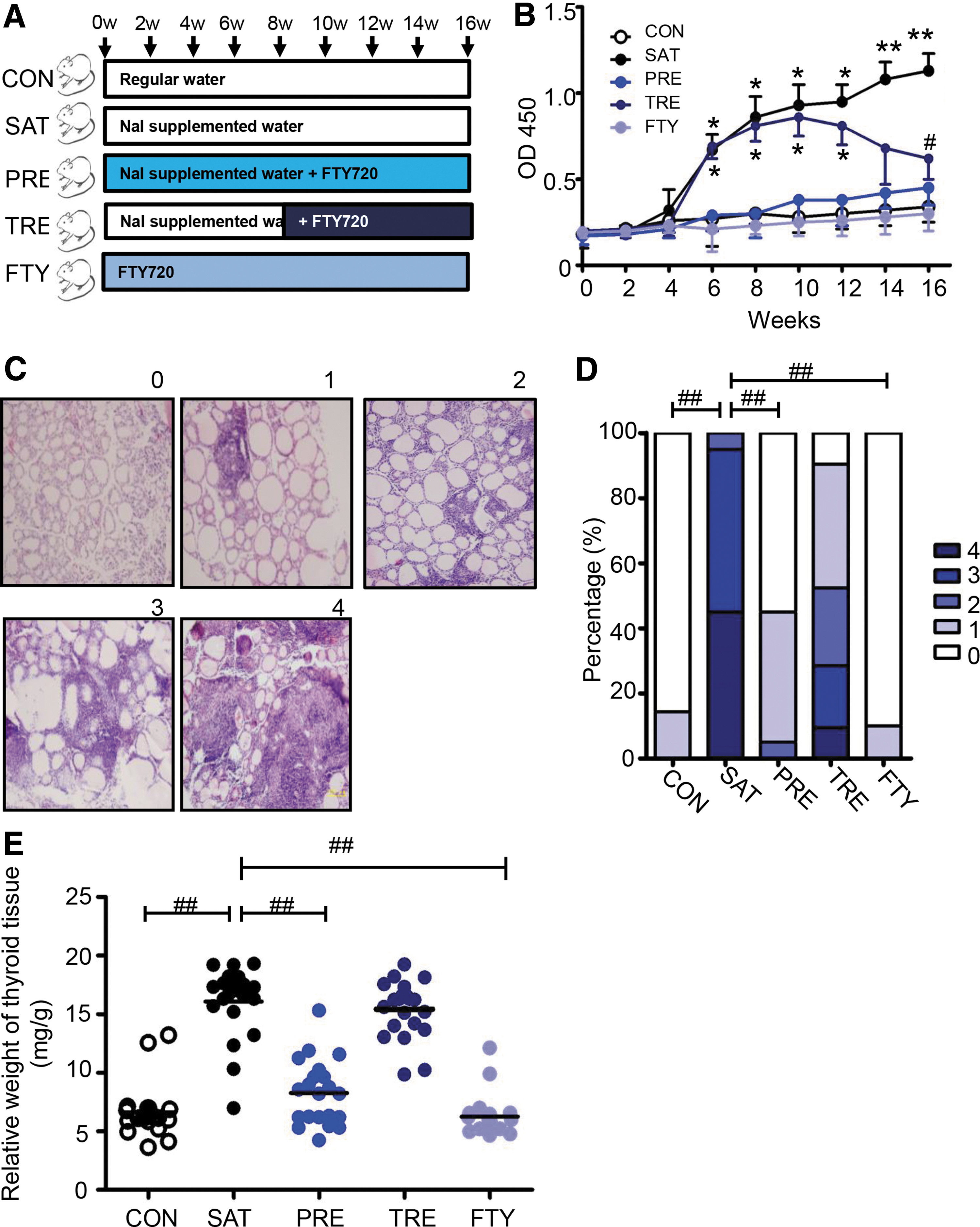
Therapeutic effect of FTY720 in vivo.
STAT3-related T cell subtype changes after FTY720 administration
To further investigate the effect of FTY720 administration, we detected not only STAT3-related cells in the CD4+T cell subtypes, such as T-helper 17 (Th17) cells, but also other inflammatory-related cell subtypes, such as Th1 and follicular helper T (Tfh) cells (Fig. 7). As indicated in Figure 7A and B, the proportion of Th1 cells was higher in the SAT group when compared with the CON group (p < 0.001) and the FTY group (p < 0.001), and the proportion of Th1 cells was significantly reduced in the PRE group but not in the TRE group in splenic CD4+T cells. Consistent with a previous report (28), Th17 cells were elevated in the SAT group when compared with the CON group and the FTY720 group (p < 0.01) (Fig. 7C–D). However, we found that the proportion of Th17 cells was significantly lower after FTY720 administration in both the PRE group and the TRE group when compared with the SAT group (p < 0.05). The proportion of Tfh cells was also higher in the SAT group. The proportion of Tfh cells was significantly reduced after FTY720 treatment in both the PRE group and the TRE group (p < 0.05) (Fig. 7 E, F). These results show that FTY720 can impede the development of AIT by changing the proportion of inflammatory cell subtypes.

Proportion of different T cell subtypes after FTY720 administration.
FTY720 significantly reduces the expression of STAT3-related inflammatory cytokines in CD4+T cells
Next, we performed quantitative PCR analysis to evaluate the expression of STAT3-related cytokines and other inflammatory-related genes in murine CD4+ T cells in different groups (Fig. 8A). We found that IL-17A, IL-21, MMP2, RORγt, IFN-γ, and IL-6 mRNAs were significantly higher in the SAT group than in the CON group, and after FTY720 treatment, the expressions of the cytokines were significantly reduced in both the TRE group and the PRE group. The expression of T-bet and IL-1β was also elevated in the SAT group. However, after FTY720 treatment, the expression of these genes decreased in the PRE group but not in the TRE group. This result is consistent with the FACS results shown in Figure 7, which indicates that the therapeutic effect of FTY720 occurs in a time-dependent manner. Notably, the S1PR1 levels were higher in the SAT group than in the CON group, and FTY720 administration decreased the expression of S1PR1 in CD4+T cells. This is also consistent with the results shown in Figure 2E. These results indicate that the blockade of S1PR1 ameliorates the severity of thyroiditis by inhibiting STAT3 and its related downstream cytokines.
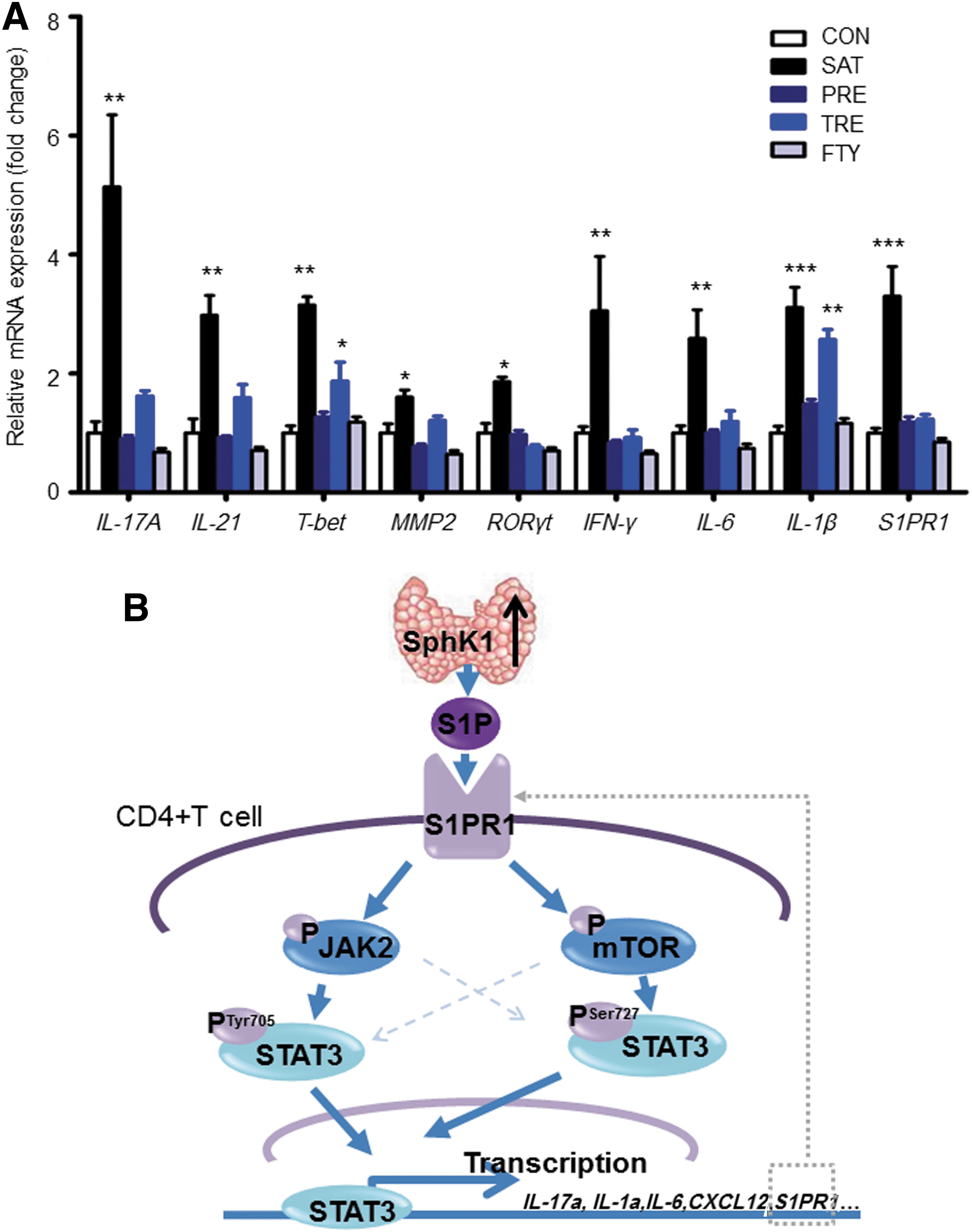
Changes of STAT3-related cytokines and inflammatory-related cytokines after FTY720 treatment.
Discussion
S1P is a bioactive sphingolipid metabolite that regulates diverse cellular processes and is important for inflammation and immune responses (8). Interest in S1P as a signaling molecule has increased dramatically over the last two decades. Studies have demonstrated that S1P can act as a “find-me” signal that may attract lymphocytes. Therefore, the potential role of S1P is of great mechanistic interest in the pathogenesis of autoimmune diseases (31). The present study is the first to demonstrate that SphK1/S1P/S1PR1 signaling is abnormal in HT patients and a SAT mouse model. We found that S1P could activate STAT3 through the S1PR1/mTOR/PSer727STAT3 and the S1PR1/JAK2/PTyr705STAT3 signaling pathways (Fig. 8B). Blocking S1PR1 by FTY720 in the mouse model attenuated the severity and development of thyroiditis by reducing the inflammatory-related T cell subtypes and inhibiting the expression of inflammatory cytokines.
The present study demonstrates that the expression of SphK1 is higher in the thyroid tissues of patients with thyroiditis and in the SAT mouse model (Fig. 8B). This may be due to many of the cytokines in the inflammatory microenvironment, such as tumor necrosis factor (TNF)-α and interleukin (IL)-1β, which can stimulate SphK1 and upregulate its expression (8,32). Several studies have demonstrated that TNF-α can activate SphK1 through ERK1/2 and promote its intracellular functions (8,33). Another study also demonstrated that SphK1 plays a key role in TNF-α-induced arthritis (32). Importantly, it has been demonstrated that proinflammatory cytokines, such as TNF-α and IL-1β, are elevated in thyroid autoimmunity (34). TNF-α, along with IL-1β, increases the apoptosis of thyrocytes which further promotes the development of autoimmune thyroiditis (35 –38). Studies have also demonstrated that sustained expression of TNF-α is important for the maintenance of inflammation and for promoting fibrosis in AIT. Similarly, it has been reported that the expression of SphK1 is elevated in biopsy specimens of patients with primary Sjögren's syndrome (39) and in the synovium of patients with rheumatoid arthritis (10); S1P has also been found to be elevated in the cerebrospinal fluid of multiple sclerosis patients (40). S1PR1 has been found to be elevated in labial salivary glands of primary Sjögren's syndrome patients (39) and in the splenocytes of mice with autoimmune hepatic injury (20). In addition, the expression of S1PR1 is associated with the severity of experimental cerebrospinal encephalomyelitis (11). All of these observations suggest that SphK1/S1P/S1PR1 signaling may be important in the pathogenesis of autoimmune diseases.
Although various autoimmune diseases have been shown to aberrantly elevate S1P, the underlying molecular mechanism of elevated S1P on immune cells remains unclear. In particular, the effect of S1P on CD4+T cells, which is of great importance in the development of HT, has not been well investigated. Therefore, it is critical to unveil the factors that determine STAT3 activity. The present study shows that S1P increases PTyr705STAT3, which is mediated by the S1PR1-JAK2 pathway. This is consistent with previous studies in the field of tumor immunology. Our data also demonstrate that S1P regulates PSer727STAT3 via the S1PR1-mTOR pathway. This is supported by previous reports in which sequence analysis of STAT3 revealed the existence of a putative mTOR signaling motif, indicating the potential mTOR-mediated phosphorylation of STAT3 (41,42).
Our data indicate that the incidence of autoimmune thyroiditis was lower after FTY720 administration in the PRE group. Even though the average score of thyroiditis in the TRE group was lower than it was in the SAT group, the incidence of thyroiditis still showed no statistical difference when compared to the SAT group. This indicates that long-term treatment was more effective than short-term treatment. This may because long-term treatment with FTY720 can regulate the unbalance of CD4+T polarization. Clinically, the role of S1PRs in autoimmune diseases has received considerable attention since the recent clinical approval of the drug fingolimod (also known as FTY720). FTY720 acts as a functional antagonist of S1PR1, has shown to be effective in human multiple sclerosis phase 3 clinical trials, and received Food and Drug Administration approval as an oral therapy for relapsing forms of multiple sclerosis in September 2010 (43,44). Oral FTY720 arrests immune trafficking and effectively decreases the relapse rate up to 50% in patients with multiple sclerosis by blocking S1PR1-STAT3 and reducing the proportion of Th17 cells in the blood (44 –46). Furthermore, FTY720 has also been found to be effective against type 1 diabetes mellitus and rheumatoid arthritis (47,48).
Interestingly, we found the expression of S1PR1 was associated with the activity of STAT3. As reported previously by Lee et al. and several separate groups via chromatin immunoprecipitation (ChIP)-PCR and reporter luciferase assay, STAT3 is the transcriptional factor of S1PR1 and regulates its expression (19). Thereafter, Zhang and colleagues demonstrated by CHIP-Seq assay that STAT3 binds to the promoter of S1PR1 (49). These observations also explain why the S1PR1 was elevated in the SAT model and the mutual regulated relationship between the expression of S1PR1 and the activity of STAT3. Thus, all of these results indicate that there is a S1PR1-mTOR-PSer727STAT3 and a S1PR1-JAK2-PTyr705STAT3 positive forward feedback loop in AIT (Fig. 8B).
In summary, although STAT3 is a promising target for autoimmunity therapy, effectively inhibiting the activity of a transcription factor remains a challenge because of their intracellular localization and lack of enzymatic activity. The present study demonstrates that S1PR1 is important for the activation of STAT3 in the SAT mouse model. Blocking S1PR1 by FTY720 administration could inhibit the forward feedback loop and decrease the phosphorylation of STAT3, which in turn would decrease the expression of S1PR1 (Fig. 8B). However, the importance of S1PR1 in autoimmunity is just beginning to be appreciated. Further studies focusing on male patients and other SAT mouse models are still needed. In addition, it has been reported that FTY720 causes several adverse effects, such as bradyarrhythmias, macular edema, and infection. More specific and optimized drugs and the potential importance as a target for autoimmune thyroiditis therapy remain to be further assessed.
Footnotes
Acknowledgements
We acknowledge that all of the listed authors participated sufficiently in the work and take public responsibility for its content. This work was supported by the Chinese National Natural Science Foundation (Grant 81370893) and the Important Platform of Science and Technology for University of Liaoning Province (16010).
Author Disclosure Statement
The authors hereby confirm that neither the full manuscript nor any part of it has been published or is being considered for publication elsewhere. No competing financial interests exists.
Supplementary Material
Supplementary Figure S1
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.