
Abstract
Background:
Thyroid follicular cells are characterized by the expression of a specific set of genes necessary for the synthesis and secretion of thyroid hormones, which are in turn regulated by the transcription factors Nkx2-1, Pax8, and Foxe1. Thyroid differentiation is finely tuned by the balance between positive regulatory signals, including thyrotropin (TSH), and by negative regulatory signals, such as transforming growth factor beta (TGF-β), which counteracts the action of TSH. A role for Foxe1 as a mediator of hormonal and growth-factor control of thyroid differentiation has been previously suggested. Therefore, the aim of this work was to study the mechanisms governing Foxe1 expression to define the ligands and signals that regulate one of the important factors in thyroid differentiation.
Methods:
Expression of Foxe1 was evaluated in rat PCCl3 thyroid follicular cells under different treatments. The mouse Foxe1 promoter was cloned, and site-directed mutagenesis was undertaken to study its transcriptional regulation and to identify response elements. Protein/DNA binding assays were performed to evaluate the binding of different transcription factors, and gene-silencing approaches were used to elucidate their functional roles.
Results:
In silico analysis of the Foxe1 promoter identified binding sites for Nkx2-1, Pax8, Foxe1, and Smad proteins, as well as cAMP-response element (CRE) sites. It was found that both CRE-binding protein and CRE modulator were necessary for the TSH-mediated induction of Foxe1 expression via the cAMP/PKA signaling pathway. Moreover, transcription of Foxe1 was regulated by Nkx2-1 and Pax8 and by itself, suggesting an autoregulatory mechanism of activation and an important role for thyroid transcription factors. Finally, TGF-β, through Smad proteins, inhibited the TSH-induced Foxe1 expression.
Conclusions:
This study shows that Foxe1 is the final target of TSH/cAMP and TGF-β regulation that mediates expression of thyroid differentiation genes, and provides evidence of an interplay between CRE-binding proteins, thyroid transcription factors, and Smad proteins in its regulation. Thus, Foxe1 plays an important role in the complex transcriptional network that regulates thyroid follicular cell differentiation.
Introduction
Thyroid epithelial cell differentiation is driven by the coordinated action of Nkx2-1, Foxe1, Pax8, and Hhex transcription factors, which together regulate the expression of thyroid-specific proteins responsible for thyroid hormone (TH) biosynthesis (1). While the activities of these transcription factors during thyroid differentiation and development have been studied extensively, few investigations have addressed their role in the hormonal control of thyroid function.
It is well established that TH synthesis takes place in the thyroid gland by iodination and coupling of tyrosine residues within thyroglobulin (Tg) by thyroperoxidase (TPO) (2). This complex mechanism is positively regulated by thyrotropin (TSH), which controls the transcription of Tg and Tpo genes in differentiated rat cells through the cAMP pathway (3 –5). Accordingly, Tg and Tpo should contain cAMP-response elements (CRE) in their upstream regulatory regions. Strikingly, however, no consensus CRE motif has so far been identified. A CRE-like (6) and a CRE-enhancer (7) have been described in the bovine and human Tg promoter, respectively, although these elements share only partial homology with the canonical CRE. These observations suggest that Tg and Tpo transcription by cAMP is mediated by a distinct mechanism involving other binding sites. Foxe1 binding sites were previously identified as the main TSH/cAMP cis regulatory element (8 –10), which would be consistent with the positive regulation of Nkx2-1 and Pax8 by TSH/cAMP (3,11 –14), and the negative regulation of Hhex (15). Nevertheless, none of the latter three transcription factors show the tight regulatory control by TSH/cAMP as described for Foxe1, providing further evidence for a critical role of this transcription factor in mediating hormonal regulation of TH synthesis (16).
Foxe1, formerly called thyroid transcription factor 2, is a member of the forkhead-box (Fox) family of transcription factors characterized by a well conserved 100 amino acid DNA-binding domain, known as the forkhead domain (17), which functions to open compacted chromatin structures, facilitating the binding of the transcription factor to the promoters of target genes (18). Thus, Foxe1 has been suggested to function as pioneer factor (19) in thyroid development and differentiation (1,18). Foxe1 is considered essential for thyroid gland formation (20) and migration (21) of precursor thyroid cells from the foregut endoderm, and is therefore an intrinsic component of a regulatory network of transcription factors and cofactors that initiate thyroid specification during development (22). Underscoring its importance, mutations in FOXE1 cause syndromic human phenotypes that are associated with thyroid agenesis, among other manifestations (1,2,23,24), and it has also been identified as a susceptibility gene in thyroid cancer (25 –27). Because it is essential for the hormonal control of thyroid-specific gene transcription, Foxe1 is also necessary for the maintenance of the thyroid differentiated state (8,9).
Given the crucial role of Foxe1 in thyroid cell differentiation, this study sought to understand the mechanisms that control its expression, including hormonal and growth factor regulation. It also sought to explore whether Foxe1 is the final target of TSH/cAMP actions that mediate Tg and Tpo gene expression. To this end, in silico analyses were used to screen the mouse and rat Foxe1 promoters for potential binding sites of transcriptional regulators. A variety of in vitro functional tests were then performed to validate the in silico predictions that led to the conclusion that Foxe1 is a crucial factor in the control of thyroid gene differentiation.
Methods
Bioinformatic analysis
To search for CREs and consensus binding sites for Pax8, Foxe1, Nkx2-1, and Smad proteins in the Foxe1 promoter, in silico analyses were performed using two bioinformatic tools: TRANSFAC (
Cloning and site-directed mutagenesis of the mouse Foxe1 promoter
To construct the pFoxe1-3.1Luc and pFoxe1-0.5Luc plasmids, DNA fragments from the mouse Foxe1 promoter corresponding to regions −3035/+193 and −1350/+243 were amplified from mouse genomic DNA by polymerase chain reaction (PCR) using forward primers containing MluI and NheI sites (5′-GGC GAC GCG TGC TAG CTG AAC TCC CAC CTC TGC TGG AT-3′ and 5′-GGC GAC GCG TGC TAG CAC ACA CAC GCA TAC ACC AGT A-3′), respectively, and reverse primers containing XhoI and SmaI sites (3′-GGC CGC TCG AGC CCG GGC GAT GAG CGC GAT GTA GCT GTA-3′ and 5′-GGC CGC TCG AGC CCG GGC GCT CGG TGA TGA ACT TGT A-3′), respectively. The fragments were digested with MluI and XhoI and subcloned into the pGL3basic-Luc vector (Promega, Madison, WI).
Site-directed mutagenesis was performed to disrupt the CRE site mapped at −1627 and the Smad3 binding site of pFoxe1-3.1Luc. For this purpose, the QuikChange Lightning Site-Directed Mutagenesis Kit (Agilent Technologies, Santa Clara, CA) was used according to the manufacturer's instructions. Complementary mutagenic primers, including the mutated regions 5′-ATA AAA AAA AAT GT-3′ and 5′-CAT AGC GT-3′, for CRE and Smad3, respectively, were designed by the web-based QuikChange Primer Design Program (
Cell culture
PCCl3 thyroid follicular cells are a continuous rat differentiated cell line expressing thyroid-specific genes and the thyroid-specific transcription factors Nkx2-1, Foxe1, and Pax8, and are an extensively used model system. Cells were cultured in Coon's modified Ham's F-12 medium supplemented with 5% donor calf serum, glutamine, antibiotics, and a six-hormone medium (complete medium) mixture: 1 nM bovine TSH, 10 μg/mL insulin, 10 ng/mL somatostatin, 10 μg/mL transferin, 10 nM hydrocortisone, and 10 ng/mL glycyl-L-histidyl-L-lysine acetate. The effects of TSH were studied by starving nearly confluent cells of TSH, insulin, and serum (0.1%) for two days (starvation medium). Ligands were added to the culture medium at the following final concentrations: 1 nM TSH (otherwise explained), 10 μM forskolin, and 10 ng/mL transforming growth factor beta (TGF-β). Ligands were added for 16 or 24 hours, depending on the experiment. The PKA inhibitor H89 (10 μM) was added to the medium one hour before hormone addition. The medium was changed every two days. HeLa cells were cultured in Dulbecco's modified Eagle's medium supplemented with 5% fetal serum, glutamine, antibiotics, and sodium pyruvate. All hormones, forskolin, and H89 were from Sigma–Aldrich (St. Louis, MO), and TGF-β was from PeproTech (Rocky Hill, NJ).
Quantitative reverse transcriptase PCR
Total RNA was extracted in TRIzol reagent (Thermo Fisher Scientific, Waltham, MA), and equal amounts of RNA were added to a reverse transcriptase (RT) reaction mix (M-MLV; Promega). Quantitative RT-PCR was performed on the Mx3000P QPCR platform (Agilent Technologies). Amplification reactions were performed with specified primers (Supplementary Table S1) and templates using the KAPA Sybr Fast qPCR Master Mix (Merck KGaA, Darmstadt, Germany) and run for 40 cycles. The specificity of the reactions was determined by melting curve analysis. Stratagene analysis software was used to remove background fluorescence. The number of cycles needed to reach the threshold level (Ct) for each sample was used to calculate the amount of each product using the 2–ΔΔCt method (28). Relative levels of the PCR products were expressed as a function of β-actin.
Protein extraction and Western blotting
Total or nuclear and cytoplasmic protein extracts from cell cultures were prepared as previously described (29). Twenty micrograms of protein were separated by sodium dodecyl sulfate polyacrylamide gel electrophoresis and transferred to nitrocellulose membranes. Membranes were blocked in PBS-T buffer (PBS +0.1% Tween 20, pH 7.5) containing 5% nonfat milk. The following primary antibodies were used: Foxe1 (#PA0200) and Pax8 (#PA0300; Biopat, Milan, Italy), CREB (sc-186), CREM (sc-377496), α-tubulin (sc-5286) β-actin (sc-1616), and Sp1 (sc-17824; Santa Cruz Biotechnology, Santa Cruz, CA). Antibodies were incubated in PBS-T containing 5% nonfat milk. Membranes were then washed four times with PBS-T buffer and incubated with species-specific secondary antibodies conjugated to horseradish peroxidase. After four washes of 10 minutes with PBS-T buffer, immunoreactive bands were visualized with the Luminol Western blot detection reagent (Thermo Fisher Scientific).
Transfections and luciferase assays
Cells were transiently transfected using calcium chloride (30). Transfections were performed on six-well plates using 200,000 cells/well seeded 24 hours before transfection. Different fragments of the mouse Foxe1 promoter fused to luciferase (pFoxe1-3.1Luc or pFoxe1-0.5Luc) were transfected alone or in combination with the following expression vectors: hCREB (31), hCRE modulator (CREM) (32), hPAX8 (33), rFoxe1 (17), hSMAD3 (34), or rNkx2-1 (35). Transfections comprised 1.5 μg/well of promoter luciferase construct and 1.5 μg/well of expression vector. Transfection efficiency was checked using 0.1 μg of the CMV-Renilla vector. In co-transfection experiments, the amount of DNA was normalized using the corresponding empty expression vectors. Cells were harvested after 48 hours, lysed, and analyzed for luciferase and Renilla activities using the dual-luciferase reporter assay system (Promega). The promoter activity was determined as the ratio between luciferase and Renilla activity, and was represented as relative luciferase activity.
Gene silencing
Creb, Crem, and Pax8 genes were silenced using ON-TARGET plus SMARTpool small interfering RNA (siRNA) molecules (Dharmacon, GE Healthcare, Lafayette, CO). At various times after transfection (16, 24, and 48 hours), mRNA and protein were extracted from cells for analysis. Results were normalized against a non-targeting siRNA (scrambled siRNA control).
Electrophoretic mobility shift assay
Band shift probes corresponding to oligonucleotides of CRE, Smad, Pax8, Nkx2-1, and Foxe1 (Supplementary Table S2) binding sites in the rat Foxe1 promoter (Sigma–Aldrich) were labeled with [γ32P]-ATP using T4 polynucleotide kinase (Promega), and purified using Quick Spin G-25 Sephadex columns (Roche Life Sciences, Mannheim, Germany). Recombinant proteins were produced by the in vitro transcription translation (TNT) Kit (Promega). Seven micrograms of each recombinant protein (CREB, CREM, SMAD3, Pax8, Foxe1, or Nkx2.1) was incubated with the labeled probe. Binding reactions were performed in a buffer containing 40 mM Hepes, pH 7.9, 75 mM KCl, 0.2 mM EDTA, 0.5 mM dithiothreitol, 150 ng/μL poly(dI-dC), and 5% Ficoll at room temperature for 30 minutes. Samples were electrophoresed on a 5% polyacrylamide gel in 0.5 × Tris borate-EDTA. For competition assays, a 100-fold excess of the same (“related”) or different (“nonrelated”) unlabeled oligonucleotide was used as indicated in each experiment.
Statistical analysis
Results are represented as fold induction ± standard error of the mean from at least three independent experiments. Student's two-tailed t-test was used to assess differences between measurements. Differences were considered statistically significant at p < 0.05.
Results
The mouse Foxe1 promoter contains consensus binding sites for Nkx2-1, Pax8, Foxe1, and Smad proteins as well as CRE elements
In silico analysis for transcription factor consensus binding sites was performed in the rat Foxe1 promoter, corresponding to 3100 base pairs upstream of the transcription start site. Two canonical CRE motifs were identified at positions −1627 and −672, one binding site for Nkx2-1 at −3049, two binding sites for Foxe1 at −261 and −1051, and two Pax8 binding sites at −279 and −845. In addition, a Smad protein binding site was identified at −672 (Fig. 1 and Supplementary File S1). The homology between the Foxe1 promoter from rat (Rattus novergicus) and mouse (Mus musculus) was >90%. The existence of binding sites for the different transcription factors prompted the transcriptional regulation of Foxe1 gene expression to be investigated, as it plays a key role in the hormonal regulation of thyroid cell differentiation (10,16).

Schematic representation of the promoter region of the rat Foxe1 gene. The geometric drawing shows the binding sites, identified in silico, of the different transcription factors studied. The putative consensus sequences identified is marked below each transcription factor binding site, together with its relative position from the transcriptional start site (+1). Color images are available online.
TSH induces Foxe1 expression through the cAMP/CREB signaling pathway
The identification of CRE consensus sequences within the Foxe1 promoter led to the hypothesis that TSH via the TSH receptor (TSH-R)/cAMP signaling pathway would activate this promoter and consequently increase Foxe1 proteins levels, which would ultimately lead to the activation of gene promoters containing Foxe1 binding sites, such as Tg and Tpo (8,9). Thus, first, the regulation of Foxe1 gene expression by TSH/cAMP was studied. To do this, PCCl3 cells were starved for 48 hours and then stimulated with TSH for 16 or 24 hours in order to extract RNA or protein, respectively. To determine the involvement of the cAMP/PKA pathway, PCCl3 cells were treated with forskolin, an adenylate cyclase activator, or with H89, a specific inhibitor of PKA. Results showed that whereas both TSH and forskolin significantly increased Foxe1 mRNA levels, these effects were inhibited by H89 (Fig. 2A). These data confirm the previous reported regulation of Foxe1 by TSH via the cAMP/PKA pathway (16). This analysis was extended by demonstrating that the increase in mRNA was mimicked at the protein level (Fig. 2B). Experiments using nuclear and cytoplasmic extracts confirmed that TSH/cAMP/PKA regulation of Foxe1 takes place in the nucleus (Fig. 2C). These data indicate the role of nuclear Foxe1 regulating the transcription of its target genes.
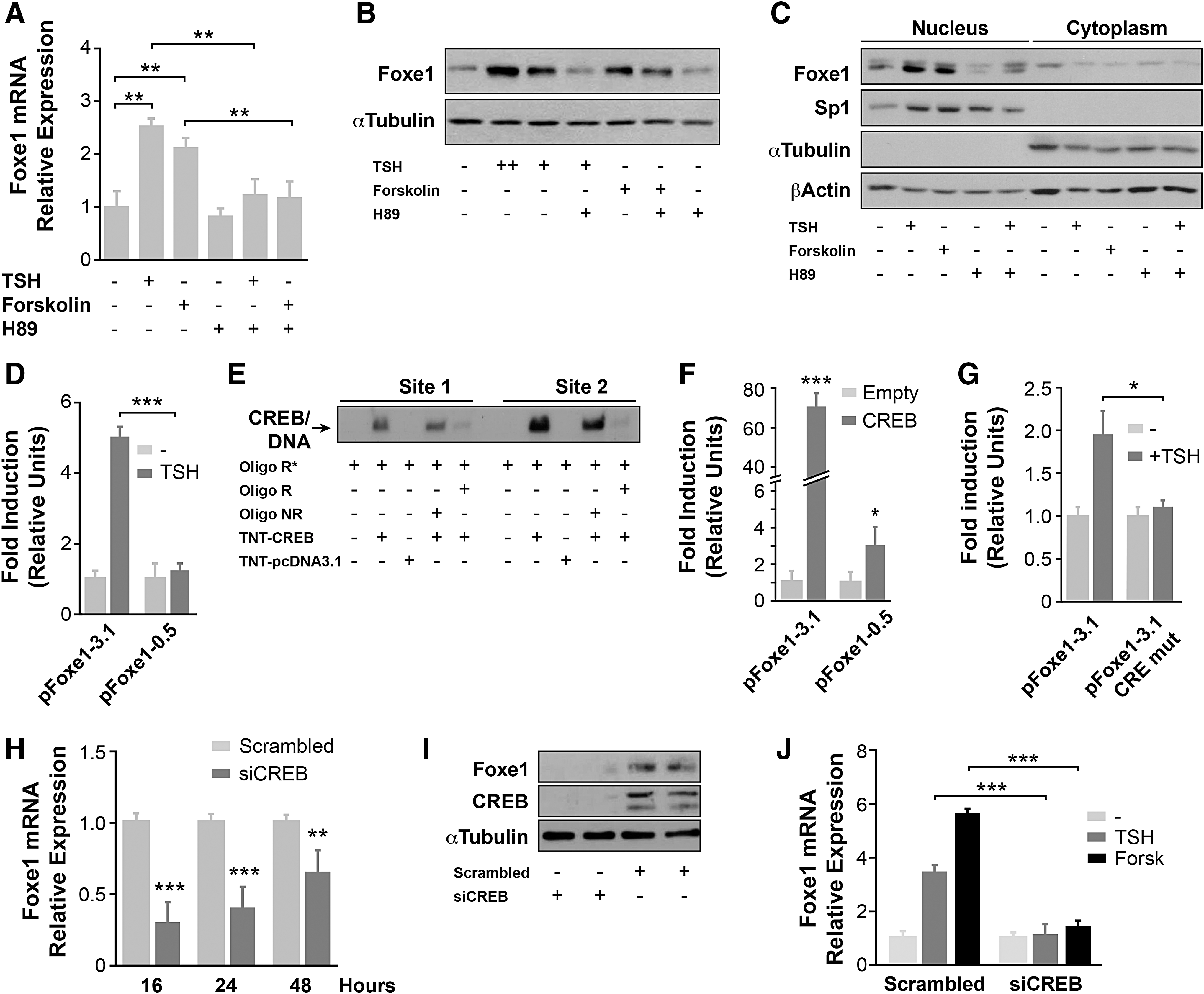
Role of the cAMP/CREB signaling pathway in the thyrotropin (TSH)-mediated regulation of Foxe1 expression. PCCl3 cells were maintained 48 hours in starvation medium (–) and then treated with different stimuli for 16 hours (
To assess whether the TSH-stimulated increase in Foxe1 gene expression operates at the transcriptional level, transient transfection experiments were performed using two Foxe1 promoter constructs: pFoxe1-3.1, which contains the two CRE sites identified in the Foxe1 promoter (−1627 and −672), and pFoxe1-0.5, which contains only the CRE site closest to the transcription initiation site (−672; see Fig. 1). Transfected PCCl3 cells were treated with TSH in starvation medium. It was found that TSH increased the promoter activity of pFoxe1-3.1-transfected cells up to four- or fivefold, whereas activation was significantly lower in cells transfected with pFoxe1-0.5 (Fig. 2D). These results demonstrate that TSH induces Foxe1 gene expression through the activation of its promoter and that the CRE site mapped at −1627 might play a fundamental role.
Since the CREB transcription factor is one of the final effectors of the cAMP/PKA signaling pathway, next its functional binding to the two CRE sites was investigated using electrophoretic mobility shift assay (EMSA) analysis. The incubation of oligonucleotides corresponding to CRE site 1 (−672) or 2 (−1627) with recombinant CREB resulted in an electrophoretic mobility shift of the protein in both cases (Fig. 2E), showing that CREB binds to both CRE sites. However, this binding was stronger at site 2 (Fig. 2E). The functionality of CREB binding to the Foxe1 promoter was next assayed in co-transfection experiments in HeLa cells after individual transfection of the two Foxe1 promoter constructs with a CREB expression vector. As shown in Figure 2F, whereas CREB significantly activated both promoter constructs, activation of the pFoxe1-3.1 promoter was ∼30 times greater. Overall, these data confirm the functionality of the two CRE sites in the Foxe1 promoter, suggesting a relevant role for the CRE site at position −1627. To validate the role of this CRE site in the response to TSH, site-directed mutagenesis was used to disrupt the binding site. The pFoxe CRE-1627-mutated promoter (pFoxe1-3.1 mutCRE) and the wild-type promoter pFoxe1-3.1 were transiently transfected into PCCl3 cells, and the promoter activities were analyzed in response to TSH, as described above. The results show that the TSH induction of Foxe1-3.1 promoter previously observed (Fig. 2D) decreased moderately in the CRE-1627-mutated promoter (Fig. 2G), suggesting that the CRE site mapped at −672 also plays a role in Foxe1 promoter activation in response to TSH.
Turning this question on its head, next CREB expression in PCCl3 cells was silenced by siRNA, and Foxe1 mRNA and protein levels were analyzed. Foxe1 mRNA levels were significantly lower in CREB-silenced PCCl3 cells than in cells transfected with the siScrambled vector (Fig. 2H), and the same was found for Foxe1 protein levels measured 48 hours after transfection (duplicate samples are shown in Fig. 2I). Interestingly, in CREB-silenced cells, the levels of Foxe1 mRNA did not increase in response to TSH or forskolin, as was found in control cells (Fig. 2J). These results demonstrate that the CREB transcription factor is necessary for the expression of Foxe1 in follicular thyroid cells and that it is essential for the regulation of Foxe1 expression by TSH via the cAMP/PKA signaling pathway.
In addition to CREB, other transcription factors such as CREM have the ability to bind CRE sites present in the promoters of cAMP-regulated genes (36). In thyroid follicular cells, CREM binding has been described in the upstream enhancer of the sodium–iodide symporter gene NIS, termed NUE. This binding was shown to increase NIS promoter activity and consequently its expression (37). Likewise, TSH induces the expression of CREM in thyroid follicular cells, which acts as a transcriptional repressor of TSH-R expression (38). Therefore, next, the study investigated whether CREM is also involved in the expression of Foxe1 by using EMSA to monitor recombinant CREM binding in the presence of an oligonucleotide containing the CRE sequence at site 2 of the Foxe1 promoter. The results showed an electrophoretic mobility shift, which confirms that CREM also binds to the Foxe1 promoter at the CRE site at position −1627 (Fig. 3A). As demonstrated above, this CRE site plays a more relevant role in the regulation of Foxe1 by TSH/cAMP. It was found that the binding of CREM to the Foxe1 promoter was also functional, as demonstrated in HeLa cells co-transfected with the pFoxe1-3.1 and pFoxe1-0.5 promoter constructs together with a CREM expression vector (Fig. 3B). Similar to the findings for CREB, the pFoxe1-3.1 promoter was more responsive to CREM than pFoxe1-0.5 was. In order to demonstrate the role of CREM on Foxe1 expression definitively, CREM in PCCl3 cells was silenced, finding that both the mRNA and protein levels of Foxe1 were reduced in siCREM-silenced cells (Fig. 3C and D, respectively). These results confirm the role of CREM in the regulation of Foxe1 expression in thyroid follicular cells.

Role of CREM in the TSH-mediated regulation of Foxe1 expression. (
Cross-talk between thyroid transcription factors in Foxe1 regulation
The transcription factors Nkx2-1, Pax8, and Foxe1 are co-expressed only in the thyroid gland, where they jointly regulate the expression of differentiating factors in thyroid follicular cells such as Tg, Tpo, or NIS. In addition, these three transcription factors form a core transcriptional network in which the expression of each of them is controlled by the other factor to mediate the expression of their target genes both during embryonic development and in differentiated follicular cells (4). Based on the results of the in silico analysis of the rat Foxe1 promoter, next the study investigated the role of Nkx2-1, Pax8, and Foxe1 in the regulation of Foxe1 expression during follicular thyroid cell differentiation.
First, EMSA was used to monitor the binding of recombinant Nkx2-1, Pax8, and Foxe1 proteins to Foxe1 promoter-derived oligonucleotides containing the in silico identified binding sites for Pax8 (site 1: −279; site 2: −845), Nkx2-1 (−3049), and Foxe1 (−261; see Fig. 1). When each of these oligonucleotides was incubated with their respective recombinant proteins, a mobility shift of the oligonucleotides was observed in all cases (Fig. 4A–D). These results confirm that the thyroid transcription factors can bind to their consensus sequences within the Foxe1 promoter. Moreover, this binding was functional, as assessed by transactivation experiments in pFoxe1-3.1-co-transfected HeLa cells, also revealing that Pax8 was the most active factor (Fig. 4E). Importantly, the combination of two transcription factors resulted in an additive increase in the transcriptional activity of the Foxe1 promoter, and the combination of all three factors induced the maximum activation (Fig. 4E).

Role of transcription factors Nkx2-1, Pax8, and Foxe1 in the regulation of Foxe1 expression. EMSAs were performed with labeled oligonucleotides derived from the Foxe1 promoter. The probes were incubated without protein or with 7 μg (
Given that Pax8 exerted the greatest increase in the activity of the Foxe1 promoter, its functional role in thyroid cells was studied by silencing its expression. The levels of Foxe1 were almost extinguished in Pax8-silenced PCCl3 compared to control-silenced cells (Fig. 4F), pointing to a pivotal role for Pax8 in Foxe1 expression.
TGF-β inhibits the inductive effect of TSH on Foxe1 expression
TGF-β, its receptors, and the Smad signaling proteins are all expressed in follicular thyroid cells, and TGF-β has been described as a growth factor involved in the differentiation, growth, and physiology of thyroid follicular cells (39,40), as well as an inhibitor of TSH and cAMP function (41,42).
Previous work showed that TGF-β inhibits TSH-induced NIS expression via a mechanism based on the reduction of Pax8 expression and the inhibition of its binding to its consensus sequence in the NIS promoter (43). Based on this evidence and the existence of Smad binding sites in the Foxe1 promoter (see Fig. 1), the potential role of TGF-β in the regulation of Foxe1 expression was studied.
Starved PCCl3 cells were treated with TSH and increasing concentrations of TGF-β, finding that TGF-β inhibited the TSH-induced Foxe1 mRNA expression in a dose-dependent manner, reaching 70% inhibition at 10 ng/mL TGF-β (Fig. 5A). Similar results were obtained when Foxe1 protein levels were analyzed (Fig. 5B).

Role of the Smad proteins in the inhibitory effect of TGF-β on TSH-induced Foxe1 expression. PCCl3 cells were maintained for 48 hours in starvation medium (–) and then treated with or without TSH + TGF-β (1–10 ng/mL) for (
The above TGF-β-mediated effect operated at the transcriptional level, as TGF-β also inhibited, in a time-dependent manner, TSH-induced pFoxe1-3.1 promoter activity in transient transfections of PCCl3 cells (Fig. 5C). As the downstream effectors of TGF-β signaling are the Smad proteins, their participation in Foxe1 transcriptional regulation was studied. EMSA was used to monitor recombinant Smad3 binding to an oligonucleotide containing the Smad binding site sequence mapped to position −746 of the Foxe1 promoter. The results showed that Smad3 bound to the oligonucleotide (Fig. 5D), suggesting that Smad3 specifically binds to its consensus sequence within the Foxe1 promoter. The Smad binding site was disrupted by mutagenesis in the context of the Foxe1 promoter construct, generating the construct pFoxe1-3.1mutSmad. The Foxe1 Smad-mutated promoter responded to TSH but failed to be inhibited by TGF-β (Fig. 5E). Altogether, these data provide evidence that the Smad binding site plays a functional role in the Foxe1 promoter.
Finally, the mechanism by which TGF-β represses TSH induction of Foxe1 transcription was investigated. Co-transfection assays were performed in HeLa cells using the pFoxe1-3.1 together with Pax8, CREB, and Smad3 expression vectors. The results revealed that Smad3 did not appreciably modify the activity of the promoter, but inhibited the inducing effects of CREB and Pax8 individually or together (Fig. 5F). Taken together, these data allow it to be concluded that TGF-β repression of TSH-induced Foxe1 promoter activity occurs through the Smad3 repression of CREB and Pax8 activation of Foxe1.
Discussion
TSH-mediated signaling is critical for thyroid cell proliferation and differentiation. In this regard, it is well recognized that the expression of several thyroid differentiation genes, including Tg, TPO, and NIS, is strongly regulated by TSH via cAMP (3). However, with the exception of a CRE site in an enhancer of the human Tg gene (7) and CRE-like sites in the bovine Tg (6) and in the rat NIS enhancer (44), CRE consensus sites within the promoters of these genes have not been found. Previously, Foxe1 consensus binding sites were identified in both the Tg and Tpo promoters, suggesting that these sites assume the role of a hormone response element (8,9). More recently, it was shown that Foxe1 functionally binds to the NUE in NIS to activate its transcription (45). Accordingly, the role of this transcription factor in the regulation of thyroid differentiation genes is well accepted.
Previously, it was described that Foxe1 mRNA levels increase in response to TSH and insulin-like growth factor 1, the two main regulators of thyroid cell differentiation (16). However, the mechanism by which TSH increases the expression of this transcription factor and whether it results in an increase in Foxe1 protein was unknown. The present results clearly demonstrate that TSH/cAMP regulates the transcriptional expression of Foxe1, which in turn is one of the main mediators of the hormonal regulation of thyroid specific genes. The regulation takes place at the nuclear level, reinforcing the idea that the action of Foxe1 at the transcriptional control is involved in thyroid cell differentiation. This observation is interesting because a gradual decrease in FOXE1 nuclear expression and accumulation in the cytoplasm has been described in thyroid tumors, which are less differentiated that PCCl3 cells (46).
Furthermore, this study provides new insights into the regulation of Foxe1 expression by this signaling pathway through the identification of two CRE sites in the Foxe1 promoter, with the more active of the two located at position −1627 where CREB and CREM transcription factors bind. Nevertheless, since the mutation of this site did not completely abolish the response to TSH, it is believed that the site at −672 also plays a role in the TSH/cAMP regulation of the Foxe1 promoter.
CREB/CREM have been shown to be tightly regulated by TSH in dog thyroid cells, providing evidence that their activity is required for the cAMP-dependent proliferation of dog epithelial cells in primary culture (47). In the case of Foxe1 regulation, both CREB and CREM seem to also play a functional role. However, CREB is thought to be the more critical of the two in the differentiation process. This reasoning is based on previous research describing the importance of CREB in the normal maintenance of the differentiated follicular cell phenotype (48,49), and also on the fact that CREM could have a dual role as an activator of NIS in differentiated cells (37) and as a repressor of the same gene in tumoral cells (50). Nevertheless, further in vivo studies are necessary to confirm the dominance of CREB over CREM on the Foxe1 promoter. Irrespective of which transcription factor elicits the major response to TSH, this study unequivocally identifies CRE sites in the Foxe1 promoter and provides evidence that Foxe1 is the main mediator of cAMP regulation in thyroid differentiation.
Interestingly, Foxe1 not only mediates the TSH/cAMP-dependent positive regulation of responsive genes, it is also involved in the negative regulation triggered by inhibitors of the TSH/cAMP-stimulated actions such as TGF-β. It is well accepted that TGF-β inhibits TSH-induced thyroid differentiation by inhibiting the expression of several differentiation genes such as Tg, TSH-R, or NIS (40,43,51). In this sense, TGF-β also inhibits the TSH-induced expression of Pax8 and its binding to the Tg and NIS promoters (43,52). This study shows that, TGF-β represses Foxe1 expression at the transcriptional level through Smad proteins, similar to what occurs with Pax8. Indeed, it was observed that TGF-β repression of TSH action is mediated by Smad protein inhibition of CREB function.
In addition, the fact that TGF-β inhibits TSH-induced thyroid differentiation supports the existence of a mechanism controlling differentiation based on the establishment of a homeostatic balance between TSH-induced signaling and other signals such as TGF-β, which act in the opposite direction. Furthermore, a role for Foxe1 as a mediator of repression of TSH-induced genes has also been demonstrated in the case of nitric oxide repression of TPO, which occurs through the inhibition of Foxe1 expression (53).
The thyroid follicular cell is the only known cell to express the transcription factors Nkx2-1, Pax8, and Foxe1 jointly. Of note, it was found that Foxe1 transcription is coordinately regulated by the main thyroid transcription factors involved in differentiation and also regulates its own transcription. This autoregulatory mechanism has been also described for murine Pax8 (54), suggesting that the transcriptional autoregulation of these factors is a hallmark of tissue-specific transcriptional control that the cell exploits to maintain a differentiated phenotype. The present results are in agreement with previous research describing an integrated transcriptional network in which the expression of each factor is interdependent with the others (22) and, in turn, participates in the co-ordinate regulation of thyroid differentiation genes (4). This process, which occurs during embryonic development, is maintained in adult thyroid cells and is likely important for the hormonal control of thyroid function. A question thus emerges as to which of these transcription factors is at the center of this network. It has been speculated that Foxe1 has an important role in light of its function as a pioneer factor, reprogramming the chromatin landscape (18). However, these in vitro studies have not yet been confirmed in vivo in thyroid development.
The case for Foxe1 as an important factor for ligand-mediated control is strengthened by the fact that it is a direct target gene of the Gli2 transcription factor, a mediator of the Sonic hedgehog (Shh) signaling pathway (55). As Shh plays an important role in thyroid development (56), we can speculate that Foxe1 is a mediator of this signaling pathway, as it is a mediator of hormonal control of thyroid differentiation in adult cells.
Knowledge of the regulatory mechanisms of Foxe1 has relevance not only in thyroid physiology but also in pathological situations. Loss of function mutations of FOXE1 result in congenital hypothyroidism as one of the hallmarks of Bamforth–Lazarus syndrome (1,23,57). Interestingly, a gain-of-function FOXE1 mutation, associated with high expression of Tg and TPO genes, has been identified in a patient with this syndrome as well (58). Thus, the data obtained herein contribute to a better understanding of the functional role of mutations found in congenital hypothyroidism.
The description of SNPs in FOXE1 conferring thyroid cancer susceptibility (25 –27) remains a matter of study because its role in tumorigenesis is not well understood. A previous study using functional approaches to examine thyroid cancer susceptibility allowed a causal variant within the human FOXE1 promoter to be identified, and a mechanism to explain the role of this gene as a risk factor in thyroid cancer to be proposed (26). These data showed that the risk allele within the human FOXE1 promoter was located close to a CRE-like sequence and recruited a complex of transcription factors, among them CREM and CREB. The present data confirm the potentially important role of these transcription factors in FOXE1 gene expression and open new horizons for the study of FOXE1 in thyroid cancer.
In conclusion, the present results underscore the central role of Foxe1 as a key mediator of TSH/cAMP/CRE induction and TGF-β/Smad repression of genes involved in thyroid differentiation through the establishment of a transcriptional regulatory circuit between Pax8, Nkx2-1, and Foxe1 itself, as delineated in Figure 6. These mechanisms are operative in normal thyroid development and in adults, and can be dysregulated in thyroid pathologies.

Schematic representation of the molecular signals and the transcription factors involved in the regulation of Foxe1 expression, as well as its transcriptional targets in follicular thyroid cells. The arrows indicate activation, and the truncated lines indicate inhibition. Color images are available online.
Footnotes
Acknowledgments
We are grateful to Dr. Kenneth McCreath for helpful comments on the manuscript and to Javier Pérez for the artwork. This work was supported by grants SAF2016-75531-R from the Ministerio de Economía, Industria y Competitividad (Spain), Fondo Europeo de Desarrollo Regional (FEDER), B2017/BMD-3724 TIRONET2-CM from the Comunidad de Madrid, and GCB14142311CRES from Fundación Española contra el Cáncer (AECC) to P.S. A.L.-M. and C.F.-M. hold an FPI fellowship from Universidad Autónoma de Madrid.
Author Disclosure Statement
All authors declare that no competing financial interests exist.
Supplementary Material
Supplementary Table S1
Supplementary Table S2
Supplementary File S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.